富營養水體中2種水生植物的根際微生物群落特征
2010-09-09 03:38:04陸開宏胡智勇梁晶晶朱津永寧波大學應用海洋生物技術教育部重點實驗室浙江寧波315211
中國環境科學 2010年11期
關鍵詞:植物
陸開宏,胡智勇,梁晶晶,朱津永 (寧波大學應用海洋生物技術教育部重點實驗室,浙江 寧波 315211)
富營養水體中2種水生植物的根際微生物群落特征
陸開宏*,胡智勇,梁晶晶,朱津永 (寧波大學應用海洋生物技術教育部重點實驗室,浙江 寧波 315211)
選取寧波市城區內河2個營養類型水體日湖(中-富營養類型)和臥彩江(重富營養類型),于2008年8~11月,利用MIDI Sherlock MIS系統和其他傳統方法,跟蹤監測了2個水體內2種常見水生凈化植物粉綠狐尾藻(Myriophyllum aquaticum)、喜旱蓮子草(Alternanthera philoxeroides)根際微生物的種類、數量及其季節變化,分析了水體主要水質參數、水體微生物與植物根際微生物之間的相關性.結果表明,水生植物根際效應顯著,不同月份植物根際微生物數量均要顯著高于水體環境;植物種類對植物根際微生物影響顯著,在日湖水體,狐尾藻根際微生物數量要顯著高于喜旱蓮子草;在臥彩江,2種植物根際細菌數量無顯著差異,放線菌、真菌數量差異顯著,植物根際微生物數量與水體環境因子之間具有一定的相關性,調查期間,日湖水溫、TN、TP、Chla與植物根際微生物數量間均無顯著相關,僅COD與喜旱蓮子草根際細菌數量顯著負相關;在臥彩江,TP與粉綠狐尾藻、喜旱蓮子草根際放線菌數量顯著負相關,其余水質參數與根際微生物數量均無顯著相關.植物根際優勢菌種以假單胞菌屬(Pseudomonas)、微球菌屬(Micrococcus)、黃桿菌屬(Chryseobacterium)、不動桿菌屬(Acinetobacter)等有機物降解微生物為主,環境因素的變化和植物種類的不同也導致了葡萄球菌屬(Staphylococcus),食酸菌屬(Acidovorax)、節桿菌屬(Arthrobacter)等特有菌種的出現.對植物根際微生物群落結構進行聚類分析表明,除日湖喜旱蓮子草根際微生物群落結構在夏、秋兩個季節相似性較高外,其余幾個處理的根際微生物群落結構相似性在21.60%~51.57%.日湖植物根際微生物多樣性夏季高于秋季,而臥彩江中夏季低于秋季;日湖中2種植物之間無論夏秋均無顯著差異,而在臥彩江,夏季喜旱蓮子草要顯著高于粉綠狐尾藻,秋季則完全相反,據此說明植物根際微生物多樣性受水體富營養化程度和植物種類等多重因素的影響.
富營養水體;粉綠狐尾藻;喜旱蓮子草;根際微生物;群落結構
Abstract:Rhizosphere microorganisms in Myriophyllum aquaticum and Alternanthera philoxeroides were identified and enumerated using Sherlock Microbial Identification System (Sherlock MIS), followed by correlation analyses with environmental factors in Lake Rihu (medium-eutrophic) and River Wocaijiang (high-eutrophic) in Ningbo from August to November 2008. Effects of plant rhizosphere on microbial community composition were significant. The abundance of microbes in the rhizosphere of macrophytes was significantly higher than that in the water column in each month. Community composition of rhizosphere microorganism differed significantly between plant species. In Lake Rihu, the number of microbes in the rhizosphere of M. aquaticum was significantly higher than that of A. philoxeroides. There was no significant difference between the abundances of bacteria in the rhizosphere of two plants, while the abundances of rhizosphere actinomyces and fungi differed significantly between two plant species in River Wocaijiang. Correlations between population size of rhizosphere microbe and environmental factors varied greatly among plant species or sampling sites. Abundance of microbes in the rhizosphere of A. philoxeroides was negatively correlated to chemical oxygen demand (COD), while there was no significant correlation with water temperature, total nitrogen (TN), total phosphate (TP) and Chl.a in Lake Rihu. The number of rhizosphere microbes showed a negative correlation with TP and no significant correlation with other environmental factors in River Wocaijiang. The dominant microbial genera in the rhizosphere of the two macrophytes were Pseudomonas, Micrococcus, Chryseobacterium, Acinetobacter etc. Some other specific functional
microbes, e.g. Staphylococcus, Acidovorax, Arthrobacter, were also found in the rhizospheres due to the heterogeneity of environmental conditions and plants. Cluster analyses using Primer 5.0 showed that the similarity of the community structure between summer and autumn was significantly higher than those between the rest seasons where similarities were more dynamic with the value of 21.60%~51.57% in the rhizosphere A. philoxeroides. Diversity of rhizosphere microorganisms was higher in summer than that in autumn in Lake Rihu. In contrast, rhizosphere microbiota was more diverse in autumn than in summer in River Wocaijiang. There was no significant difference of diversity of rhizosphere microbiota between two plants in Lake Rihu. In River Wocaijiang, the diversity of rhizosphere microorganisms of M. aquaticum was higher than that of A. philoxeroides in summer and with the opposite results in autumn. The results obtained in this study indicated that the rhizosphere microbial community structure was affected significantly by the environmental factors and plant species.
Key words:eutrophic water;Myriophyllum aquaticum;Alternanthera philoxeroides;rhizosphere microorganisms;community structure
水生植物對水體環境的修復除植物本身對氮、磷等污染物的吸收作用外,其根際微生物的作用不容忽視.有機物的降解和氮化合物的脫氮、磷化合物的轉化等主要是由植物根區的微生物活動來完成[1],根際微生物在植物的營養循環、根部環境的維持、有害污染物的降解以及植物的生長代謝方面發揮著重要的作用[2-3].在天然水體中,水生植物根區與非根區的微生物種類和數量存在較大差異,如濕地基質表面和植物根區的功能性微生物種類和數量相對較多,并且能表現出較高的根際效應[4-6].尤其是細菌,由于其在根際環境中對各種根分泌物的利用率和敏感性遠遠超過放線菌、真菌、藻類和原生動物等,它們在根際微生物中最為活躍并占主導地位,能與植物根系聯合生存并定殖和保持在根部[7],表現出強烈的根際效應.因此,研究水生植物處理污水系統中植物根際微生物的群落結構及其變化,對深入探討水生植物的凈化功能及機理具有十分重要的意義.通常植物種類、季節變化等對植物根際微生物具有很大影響
[8-9],這是否導致水生植物凈化效果的明顯差異值得探討.
本實驗通過對寧波市日湖、臥彩江2個不同富營養類型水體內2種常見水生凈化植物-粉綠狐尾藻(Myriophyllum aquaticum)和喜旱蓮子草(Alternanthera philoxeroides)根際微生物的跟蹤調查,并結合主要水化學指標的監測,研究不同營養類型水體植物根際微生物的群落結構變化,以期為進一步分析根際微生物與水生植物在污水凈化系統中的聯合作用機理,強化水生植物的水質凈化效果積累基礎資料.
1 材料與方法
1.1水生植物根樣采集
臥彩江為寧波城區內河,常年接納大量生活污水,夏季(6~9月)常暴發藍藻水華,并有間歇性黑臭現象.日湖為寧波城區公園水體,有機污染較嚴重,但無黑臭現象.2008年8~11月,在2個水體分別選擇2個采樣點,逐月采集水體中的粉綠狐尾藻和喜旱蓮子草新鮮植株根系,以無菌操作方法各采集2~3棵植物根樣,約5g,分別置于無菌10mL離心管中.同時以無菌操作采集2個水體水樣,在0℃條件下與根樣一起立即送回實驗室進行分析.
1.2理化因子的測定與分析
水溫、pH值、溶解氧(DO)等用多功能水質分析儀(Hydrolab Datasonde 4α,美國)現場測定,葉綠素a濃度(Chla)用葉綠素水下熒光分析儀(bbe FluoroProbe,德國)測定.總氮(TN)采用堿性過硫酸鉀消解紫外分光光度法測定,總磷(TP)采用鉬酸銨分光光度法測定,化學耗氧量(COD)采用堿性高錳酸鉀法測定[10].
1.3微生物數量測定
采用漩渦振蕩法分離植物根際微生物.將取自同一水體2個采樣點的同種植物根樣定量后一并放入10mL離心管中,加無菌水至10mL刻度線,手持離心管,使管底與漩渦振蕩器接觸,并與其成90°角,設轉速為1000r/min,持續振蕩15min.
植物根際微生物數量的測定參照文獻[11],采用稀釋平板法,28℃恒溫培養48h后計數,細菌、放線菌和真菌的培養分別采用牛肉膏蛋白胨培養基、高氏一號培養基和馬丁氏培養基.采用MIDI Sherlock微生物鑒定系統分別對8月和11月的微生物進行了種類鑒定[12].
1.4數據處理
利用Shannom-Wiener(H′)多樣性指數評價植物根際環境的微生物多樣性.式中:N為所有單菌落數量;Ni為屬(種)i的單菌落數量; S為所有屬(種)數.

數據處理及方差分析在SPSS13.0統計軟件上進行.其中細菌、放線菌和真菌數量經對數變換后再進行方差分析及多重比較.根際微生物數量與水質參數之間相關性在SPSS軟件中進行分析(采用Pearson系數).利用PRIMER 5.0軟件對根際微生物群落結構進行聚類分析.
2 結果與分析
2.1根際微生物數量及其動態
調查季節內,2種水生植物根際微生物數量均以細菌所占比例最大,其次是放線菌,真菌數量最少(表1、表2).2個水體中水生植物根際效應都非常明顯,各個月份水生植物根際微生物數量均顯著高于水體微生物數量,日湖和臥彩江植物根際微生物量要比水體分別高2~3個和1~2個數量級.

表1 日湖、臥彩江水體不同時期微生物數量(×106CFU/mL)Table 1 Microbe population in the water with various months(×106CFU/mL)

表2 水生植物根際不同時期微生物數量(×106CFU/gDW)Table 2 Microbe population in the rhizosphere of acquatic plants with various months(×106CFU/gDW)

表3 日湖、臥彩江水溫、TN、TP、COD、Chla變化Table 3 Variations of WT, TN, TP, COD and Chla concentrations in Rihu Laker and Wocaijiang River
日湖粉綠狐尾藻根際細菌數量要顯著高于喜旱蓮子草(P<0.05),而放線菌、真菌的數量在2種植物之間無顯著差異(P>0.05);在臥彩江,粉綠狐尾藻與喜旱蓮子草的根際細菌數量無顯著差異(P>0.05),而喜旱蓮子草根際的放線菌數量要顯著高于粉綠狐尾藻(P<0.05),真菌數量在2種植物間亦有顯著差異(P<0.05).
就不同月份而言,日湖植物根際微生物數量在10月份最高,臥彩江則以11月份最盛,2個水體中均9月數量最少.總體而言,秋季(10、11月)植物根際微生物數量要顯著高于夏季(8、9月)(P<0.05).
2.2植物根際微生物數量與主要環境因子的相關性
監測了2個水體8~11月份水溫、TN、TP、COD和Chla等主要理化指標(表3).2個水體間水溫和Chla濃度差異不顯著(P>0.05),TN、TP、COD差異極顯著 (P<0.01),臥彩江富營養程度要顯著高于日湖,其中日湖水體屬中-富營養型,臥彩江則為重富營養類型[13].8~11月,日湖和臥彩江水溫、TN、TP、Chla均呈逐漸下降趨勢,COD含量從8~10月逐漸下降,而在11月份又有所回升,這可能與11月份水體中部分植物的枯萎和腐爛降解有關.
相關性分析表明,植物根際微生物數量雖有明顯的季節差異,但與水溫之間無顯著相關關系.日湖中,TN、TP與植物根際微生物數量間無顯著相關;而臥彩江中,TP與粉綠狐尾藻(r=-0.983, P<0.05)、喜旱蓮子草(r=-0.992,P<0.05)的根際放線菌數量顯著負相關,TN與植物根際微生物數量則無顯著相關.日湖中COD與水體微生物數量(r=-0.994, P<0.05)、喜旱蓮子草根際細菌數量(r=-0.955, P<0.05)同樣呈顯著負相關.2個水體中水體微生物數量和植物根際微生物數量間呈顯著正相關,而水體Chla濃度與植物根際微生物數量無顯著相關性.

表4 日湖水體及水生植物根際微生物群落結構差異Table 4 Differences of microbial community structure between rhizosphere of macrophytes and water column in Lake Rihu
2.3根際微生物群落特征
8月,日湖植物根際優勢微生物主要為微球菌屬(Micrococcus)、假單胞菌屬(Pseudomonas)、葡萄球菌屬(Staphylococcus)、節桿菌屬(Arthrobacter),其中微球菌屬、假單胞菌屬是水體和2種植物的共有種;臥彩江植物根際優勢種也包括微球菌屬、葡萄球菌屬,但也出現了食酸菌屬(Acidovorax)、芽孢桿菌屬(Bacillus)、黃桿菌屬(Sphingomonas),尤其在粉綠狐尾藻根際,優勢微生物種類差異較大,假單胞菌屬、微球菌屬、食酸菌屬是水體和2種植物的共有種.至秋末11月,假單胞菌屬依然是2種植物根際的主要優勢種類之一,且是水體和2種植物的共有種,與夏季不同,根際優勢種中出現不動桿菌屬(Acinetobacter)、黃桿菌屬.由于不同植物根系環境的特殊性,2種植物在不同季節均出現一些特有種(表4,表5).

表5 臥彩江水體及水生植物根際微生物群落結構差異Table 5 Differences of microbial community structure between rhizosphere of macrophytes and water column in River Wocaijiang
應用水生植物在兩個季節的根際微生物種類作統計變量進行聚類分析可以清晰地看出不同情形之間的親疏程度(圖1).8月、11月日湖喜旱蓮子草根際微生物群落結構相似性為75.94%,受季節變化影響最小,而其他樣品的根際微生物群落結構在8月、11月的相似性為31.55%~33.86%,受到季節變化的影響顯著.同一水體內由于植物種類差異導致根際微生物群落結構的差異,8月份日湖和臥彩江內狐尾藻與喜旱蓮子草根際微生物群落結構相似性分別為41.28%和51.57%,11月份則僅分別為38.57%和21.60%;同樣,同種植物由于水體環境差異也導致根際微生物群落結構的不同,8月和11月日湖和臥彩江內狐尾藻根際微生物群落結構的相似性分別為47.28%和21.60%,喜旱蓮子草根際微生物群落結構的相似性則更小,分別為27.53%和24.83%.
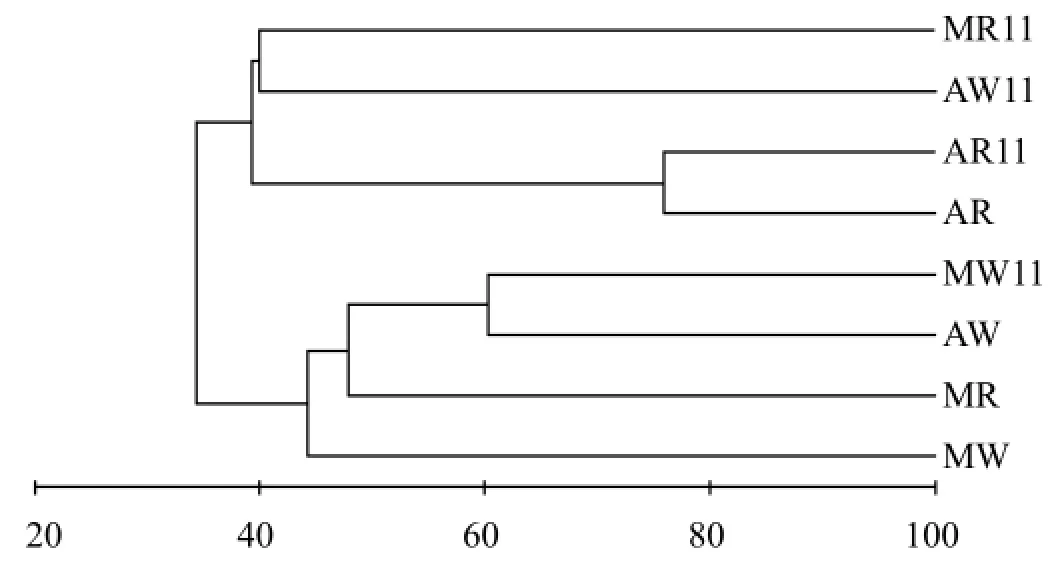
圖1 水體及水生植物根際微生物聚類分析Fig.1 Cluster analysis of population of microbes in rhizosphere of aquatic plants and water
由表6可見,8月份日湖水體和水生植物根際微生物多樣性處于同一水平,臥彩江則以喜旱蓮子草根際微生物多樣性最高, 11月份日湖水體微生物和植物根際微生物的多樣性較8月份都有所降低,而臥彩江則比8月份有所提高,且植物根際微生物多樣性顯著高于水體微生物.總體而言,夏季水質相對較好的日湖植物根際微生物多樣性要高于時有惡臭的臥彩江,而在秋末,隨著臥彩江水質狀況的改善,其微生物多樣性要高于營養水平較低的日湖水體.

表6 水體及植物根際微生物Shannon-Wiener多樣性指數(H′)變化Table 6 The Shannon-Wiener diversity index of microbes in rhizosphere of aquatic plants and water
3 討論
3.1水生植物根際微生物與環境的相互關系
水生植物根際微生物數量的分布受到水溫、植物生長狀況和代謝活動、水體營養鹽濃度等各種因子的影響,導致不同植物間根際微生物的數量有所差異.由于微生物的生長繁殖及活性受到溫度變化的顯著影響,同時植物在不同季節的生長狀況和代謝活動不同,對有機質的吸收利用存在差異,導致植物根際微生物群落的變化,并影響植物對污水中污染物的去除效果[14].研究結果表明,10、11月份植物根際細菌、放線菌和真菌數量都要高于8、9月份數量,這與梁威等[15]對人工濕地植物根區微生物季節變化的研究結果相同.9月底,臥彩江內的主要植物類群粉綠狐尾藻開始枯萎,11月份多數狐尾藻趨于死亡,釋放出的有機物質對水體造成二次污染,對微生物的數量變化產生了較大程度的影響.而真菌能生長在死亡或腐爛的植物上,引起纖維素、木質素果膠等的分解[8],因此真菌數量在秋季有所增加.
有研究表明[14],微生物活動是構建濕地凈化污水系統中TN、COD的主要去除途徑,植物根際微生物的數量分布和水體環境因子變化之間具有相關性,而TP、TSS等與微生物數量間沒有顯著的相關性.在本試驗中,日湖內TN與植物根際微生物呈正相關,說明TN的去除和植物根際微生物有一定的聯系,同時這與水體富營養化程度有關,在臥彩江未出現類似情況,可能是由于TN濃度過高,已超出微生物對其的正常響應.TP、COD與植物根際微生物呈負相關,這可能是因為9月底水生植物優勢種粉綠狐尾藻已開始枯萎,11月份部分狐尾藻已經死亡進而釋放出氮、磷及有機物質,對水體造成二次污染,導致11月份TP、COD濃度都要高于10月份,并因此呈現根際微生物數量與TP、COD的負相關關系,這種情況在重度富營養化水體臥彩江中表現尤為明顯.利用水生植物凈化污染水體的過程中,微生物是污水凈化的主要參與者,對整個凈化生態系統具有重要的影響,但微生物所能發揮的作用受到水體環境條件的綜合影響,在富營養水體中,水溫、TN等對微生物數量的影響較為顯著,而在重度富營養型水體中,水體中溶解氣體等環境因子可能成為更加重要的制約因素.
3.2水生植物根際微生物群落特征
水生植物根際微生物多樣性受植物類型、植物長勢、水體富營養程度等多種因素的影響,同時由于根系的分泌物和脫落物對微生物具有選擇和富集的作用,必然導致植物根際微生物群落多樣性的變化[16].對植物根際微生物群落結構的聚類分析結果顯示,植物根際微生物群落結構受到水體環境條件、植物種類影響顯著.馮勝等[17]對太湖的研究表明,水體中細菌的群落結構隨著水體富營養化程度的改變產生明顯變化,細菌數量呈現隨水體營養水平增加而上升的趨勢,細菌群落多樣性隨水體營養水平增加呈現顯著的下降趨勢,超富營養和接近超富營養湖區的水體中細菌群落多樣性最低.Katleen等[18]對4個不同營養水平淺水湖泊中微生物種群多樣性變化研究結果表明,隨著湖泊營養水平的改變,水體中細菌的群落結構發生了明顯變化.王曉丹等[19]對表流和潛流濕地細菌多樣性進行研究后同樣發現,隨著水體有機負荷的增加,微生物的生長受到抑制,生長速度減慢,多樣性減少.8月份,臥彩江水體微生物多樣性明顯低于日湖,原因可能是臥彩江有機污染嚴重,并經常發生惡臭,限制了中污和寡污帶微生物的生長繁殖,同時,臥彩江頻繁的藻類水華暴發和衰敗,可能對根際微生物群落也產生了一定的影響[20].11月份,雖然水溫的回落不利于微生物的生長,但2個水體富營養化程度較8月份都有所降低,N、P等無機營養鹽因子成為微生物生長的主要限制因子,同時大量藻類、水生植物分泌物及殘體等容易被微生物吸收的溶解性有機物,可能為細菌的生長提供了豐富的營養[21],因而促進了多種微生物的增長.
根際微生物群落特征受到植物根系形態結構與生長特點不同的影響.喜旱蓮子草屬挺水植物,喜固著基質生長,須根短較密;粉綠狐尾藻屬浮葉植物,莖根長而發達;這些均能導致根際微生物種類與數量的不同.本研究分析結果顯示,植物種類不同對其根際微生物群落結構的影響要超過其他因素的影響.各個時期植物根際優勢種也存在顯著差異,有研究認為[22],假單胞菌和芽孢桿菌為土壤典型的發酵類型微生物,它們對植物根系的分泌物和脫落物反應迅速,生長繁殖很快,從植物根際中可以廣泛的分離出來,因而通常具有很高的分離率.沈耀良等[23]研究表明,不同類型濕地基質具有不同的優勢菌屬:人工蘆葦濕地中的優勢菌屬主要有3大類,分別是假單胞桿菌屬、產堿桿菌屬及黃桿菌屬,與天然的蘆葦濕地相比,人工濕地中的優勢微生物種群組成與其基本相同,這幾種優勢菌屬的微生物生長快速,加之其體內大多含有降解質粒,所以它們是廢水中有物污染物降解的主要微生物種群.本研究中,8月份和11月份日湖、臥彩江植物根際都分離到假單胞菌的結果與此相符.對污水凈化濕地模擬系統的研究表明[24],濕地微生物優勢種主要為耐污能力較強的埃希氏菌屬和以碳氫化合物為主要營養來源的假單胞菌屬,尤其是假單胞菌在去除碳源污染物方面起著重要的作用.李輝等[25]對人工濕地中氨化細菌去除有機氮的效果研究后認為,人工濕地去除有機氮主要由于氨化細菌的作用,而芽孢桿菌屬、假單胞菌屬為人工濕地中氨化細菌的優勢菌屬.結構是功能的反映,在污水處理系統中,首先是芽孢桿菌類對大分子結構的有機物進行破壞,繼而由假單胞菌類、大腸菌類等利用這些被破壞的物質,并作進一步地降解.詹發萃等[26]對鳳眼蓮根區微生物異養細菌群落特征研究后發現,其優勢種類主要為氣單胞菌屬、微球菌屬、假單胞菌屬、土壤桿菌屬和芽飽桿菌屬.李科德等[27]也認為假單胞菌屬、產堿桿菌屬和黃桿菌屬等快生型細菌在有機污染物的分解代謝中起著十分重要的作用.本研究結果與該結論在一定程度上吻合,臥彩江和日湖2種水生植物的根際微生物主要優勢種為假單胞菌、黃桿菌和微球菌等,說明它們的存在可能促進水體有機物的降解.8 月份,植物根系優勢菌種包括食酸菌屬、黃桿菌屬、節桿菌屬等,均屬常見具有脫硝作用菌種,適宜的環境條件刺激了這幾類菌群的生長.綜上所述,植物根際微生物與植物種類、根系類型和水體條件等均有密切關系.
值得一提的是,本研究在采用MIDI Sherlock微生物鑒定系統對微生物進行種類鑒定的過程中,仍需依賴于傳統的微生物培養法.由于這種方法人為限定了一些培養條件,無法全面反映微生物生長的自然條件,常常造成某些微生物的富集生長,而另一些微生物缺失,可能導致部分微生物資源遺漏.
4 結論
4.1粉綠狐尾藻和喜旱蓮子草根際效應明顯,不同月份植物根際微生物數量均要顯著高于水體環境,根際微生物數量均以細菌所占比例最大,其次為放線菌,真菌數量最少;根際微生物數量受植物種類、植物生長狀態、水體營養鹽濃度和其他多種環境因子的綜合影響.
4.22種水生植物根際優勢菌種均以假單胞菌屬、微球菌屬、黃桿菌屬、不動桿菌屬等有機物降解微生物為主,但不同植物和水體環境的植物根際微生物群落結構有較大差異.植物根際微生物多樣性受環境條件和植物種類等多重因素的影響,水體高度富營養化和惡臭、藍藻水華等極端環境會導致植物根際微生物多樣性下降,但適度的水體富營養水平有利于更多微生物種類的分布與生長.
[1] Stottmeister U, Wie?ner A, Kuschk P, et al. Effects of plants and microorganisms in constructed wetlands for wastewater treatment [J]. Biotechnology Advances, 2003,22:93–117.
[2] Elsgaard L, Petersen S O, Debosz K. Effects and risk assessment of linear alkylbenzene sulfonates in agricultural soil.1. Short-term effects on soil microbiology [J]. Environmental Toxicology and Chemistry, 2001,20(8):1656-1663.
[3] Filip Z. International approach to assessing soil quality by ecologically-related biological parameters [J]. Agriculture Ecosystems and Environment, 2002,88(2):689-712.
[4] Ottová V, Balcarová J, Vymazal J. Microbial characteristics of constructed wetlands [J]. Water Science and Technology, 1997, 35(5):117-123.
[5] Machate T, Noll H, Behrens H, et al. Degradation of phenanthrene and hydraulic characteristics in a constructed wetland [J]. Water Research, 1997,31(3):554-560.
[6] 吳 偉,胡庚東,金蘭仙,等.浮床植物系統對池塘水體微生物的動態影響 [J]. 中國環境科學, 2008,28(9):791-795.
[7] 李潞濱,劉 敏,楊淑貞,等.毛竹根際可培養微生物種群多樣性分析 [J]. 微生物學報, 2008,48(6):772-779.
[8] 項學敏,宋春霞,李彥生,等.濕地植物蘆葦和香蒲根際微生物特性研究 [J]. 環境保護科學, 2004,24:35-38.
[9] Liang W, Wu Z B, Cheng S P, et al. Roles of substrate microorganisms and urease activities in wastewater purification in a constructed wetland system [J]. Ecological Engineering, 2003. 21:191-195.
[10] 魏復盛.水和廢水監測分析方法 [M]. 北京:中國環境科學出版社, 2002.
[11] 范秀榮,李廣武,沈 萍.微生物學實驗 [M]. 北京:高等教育出版社, 1980.
[12] Sherlock Microbial Identification System, Version 415, MIS Operating Manual [M]. Newark, DE: MIDI, Inc, 2002.
[13] 金相燦.中國湖泊環境 [M]. 北京:海洋出版社, 1995.
[14] 梁 威,吳振斌,詹發萃,等.人工濕地植物根區微生物與凈化效果的季節變化 [J]. 湖泊科學, 2004,16(4):312-317.
[15] 梁 威,吳振斌,周巧紅,等.構建濕地基質微生物與凈化效果及相關分析 [J]. 中國環境科學, 2002,22(3):282-285.
[16] Wasaki J, Rothe A, Kania A. Root exudation, Phosphorus acquisition and microbial diversity in the rhizosphere of white lup ine as affected by phosphorus supply and atmospheric carbon dioxide concentration [J]. Journal of Environmental Quality, 2005, 34(6):2157-2167.
[17] 馮 勝,秦伯強,高 光.細菌群落結構對水體富營養化的響應[J]. 環境科學學報, 2007,27(11):1823-1829.
[18] Katleen V G, Tom V, Nele V. Characterization of bacterial communities in four freshwater lakes differing in nutrient load and food web structure [J]. FEMS Microbiology Ecology, 2005, 53:205-220.
[19] 王曉丹,翟振華,趙 爽,等.北京翠湖表流和潛流濕地對細菌多樣性的影響 [J]. 環境科學, 2009,30(1):280-288.
[20] Yannarell A C, KentA D, Lauster G L, et al. Temporal patterns in bacterial communities in three temperate lakes of different trophic status [J]. Microbial Ecology, 2004,46:391-405.
[21] Baines S B, Pace M L. The production of dissolved organic matter by phytoplankton and its importance to bacteria patterns across marine and freshwater systems [J]. LimnolOceanogr, 1991, 36:1078-1090.
[22] 湛方棟,陸引罡,關國經,等.烤煙根際微生物群落結構及其動態變化的研究 [J]. 土壤學報, 2005,42(3):488-494.
[23] 沈耀良,王寶貞.人工濕地系統的除污機理 [J]. 江蘇環境科技, 1997,3:1-6.
[24] 陳博謙,尹澄清.污水凈化濕地模擬系統中細菌和藻類的生態分布研究 [J]. 生態學報, 1998,18(6):634-639.
[25] 李 輝,徐新陽,李培軍,等.人工濕地中氨化細菌去除有機氮的效果 [J]. 環境工程學報, 2008,2(8):1044-1047.
[26] 詹發萃,鄧家齊,夏宜琤,等.鳳眼蓮根區異養細菌的群落特征與異養活性的研究 [J]. 水生生物學報, 1993,17(2):150-156.
[27] 李科德,胡正嘉.蘆葦床系統凈化污水的機理 [J]. 中國環境科學, 1995,15(2):140-144.
Characteristics of rhizosphere microbial community structure of two aquatic plants in eutrophic waters.
LUKai-hong*, HU Zhi-yong, LIANG Jing-jing,ZHU Jin-yong (Key Laboratory of Applied Marine Biotechnology, Ministry of Education, Ningbo University, Ningbo 315211, China). China Environmental Science, 2010,30(11):1508~1515
X172
A
1000-6923(2010)11-1508-08
陸開宏(1964-),男,浙江余姚人,教授,博士,主要從事水污染生態學研究.發表論文60余篇.
2010-02-21
國家“863”項目(2007AA10Z409);國家自然科學基金資助項目(30771658);浙江省自然科學基金資助項目(Z505319)
* 責任作者, 教授, lukaihong@nbu.edu.cn
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
