番茄葉霉病菌產毒條件研究
2010-09-28 01:34:10李小波康立功李景富
植物保護 2010年1期
李小波, 康立功, 路 盼, 張 賀, 李景富
(東北農業大學園藝學院番茄研究所,哈爾濱 150030)
番茄葉霉病是番茄的主要病害之一,在歐洲、美洲、亞洲、大洋洲、非洲都有該病發生的報道,在我國以華北和東北地區受害最為嚴重[1]。番茄葉霉病是由半知菌亞門黃枝孢菌引起的真菌性病害,具有間歇暴發的特點。近年來,番茄普遍采取保護地種植方式,而保護地環境條件客觀上促進了葉霉病的大規模發生和流行,因此該病害在保護地有逐年加重的趨勢[2],目前,已成為保護地番茄高產、穩產的重要限制因素。
番茄葉霉病菌分泌的毒素是導致葉霉病發生及給番茄造成傷害的關鍵生化因子[3],因此探明番茄葉霉病菌分泌毒素的條件,有利于番茄抗/感葉霉病菌生化機理的研究和篩選、鑒定番茄抗葉霉病種質資源。前人研究結果表明:番茄葉霉病菌產生毒素為多組分糖蛋白[4],該毒素對番茄組織細胞具有明顯的損傷作用,主要包括:誘導葉組織壞死,加速胼胝質沉積和電解質滲漏,提高脂加氧酶活性和脂質過氧化作用等[5-6]。
番茄葉霉病菌具有明顯的生理分化現象,至少有13個生理小種已被報道[7]。目前,國內外對番茄葉霉病菌的研究,僅限于番茄葉霉病菌的分離、純化和生理小種鑒定等;而有關番茄葉霉病菌毒素的產生條件國內外均鮮見報道。本試驗以東北農業大學番茄研究所保存的番茄葉霉病菌生理小種1.2.3.4及番茄感病品種東農704為研究材料,系統詳盡地對番茄葉霉病菌產毒條件、提取方法、濃度測定及生物學活性分析方法進行了研究,以期為番茄抗葉霉病育種奠定基礎和提供現實條件。
1 材料與方法
1.1 供試菌株
番茄葉霉病菌生理小種1.2.3.4及番茄感病品種東農704,均由東北農業大學番茄研究所提供。
1.2 培養條件
1.2.1 培養液種類
在無菌條件下,分別采用 Czapek、改進的Czapek、Fries、Richard 、PS 、PD 6 種培養液,培養液配方見文獻[8],放入直徑為8mm的菌碟5片,pH 7,25℃振蕩培養20 d,3次重復。
1.2.2 pH
在無菌條件下,分別在 pH 3、5、6、7、9的 PS 培養液中,放入直徑為8mm的菌碟5片,25℃恒溫振蕩培養20 d,3次重復。
1.2.3 培養時間
在無菌條件下,分別在pH7的PS培養液中放入直徑為8mm的菌碟5片,25℃恒溫振蕩培養7、14、21 d和28 d,3次重復。
1.2.4 溫度
在無菌條件下,分別在pH7的PS培養液中放入直徑為8mm 的菌碟5 片,分別在 22、24、26、28、30℃及34℃下振蕩培養20 d,3次重復。
1.2.5 培養方式
在無菌條件下,在pH 7的PS培養液中放入直徑為8 mm的菌碟5片,分別在25℃下進行靜止培養、每天振蕩12 h培養和全天振蕩培養20 d,3次重復,定時測定毒素濃度。
1.2.6 光照
在無菌條件下,在pH 7的PS培養液中放入直徑為8 mm的菌碟5片,分別在25℃下振蕩培養20 d,設黑暗、每天光照12 h和24 h共3個處理,3次重復,定時測定毒素濃度。
1.2.7 菌碟數
在無菌條件下,分別在pH7的PS培養液中放入3、5、7碟直徑為8 mm的菌碟,在25℃下振蕩培養20 d,3次重復,定時測定毒素濃度。
1.3 葉霉病菌粗毒素的提取與濃度的測定
1.3.1 毒素蛋白的提取
參照陳旭升[9]介紹的方法,略作改進。具體為:將上述培養的菌液在無菌條件下,每樣品每次吸取5m L,5 000 r/min離心15m in,取上清液,再經濾膜(d=0.45μm)過濾,粗毒素濾液做顯微觀察,確定無菌后用 90%飽和度硫酸銨沉淀過夜。再經15 000 r/min離心20 min,棄上清液得蛋白質粗提物。用0.05 mol/L磷酸緩沖液含有0.15 mol/L NaCl(pH=7.0)溶解蛋白質,15 000 r/min離心5 min,得上清蛋白質溶液。
1.3.2 毒素蛋白的濃度測定
按考馬斯亮藍G-250法[10],以牛血清白蛋白作標準曲線(圖1),在722分光光度計上595 nm處測定濾液中的毒素蛋白濃度,以磷酸緩沖液作對照。
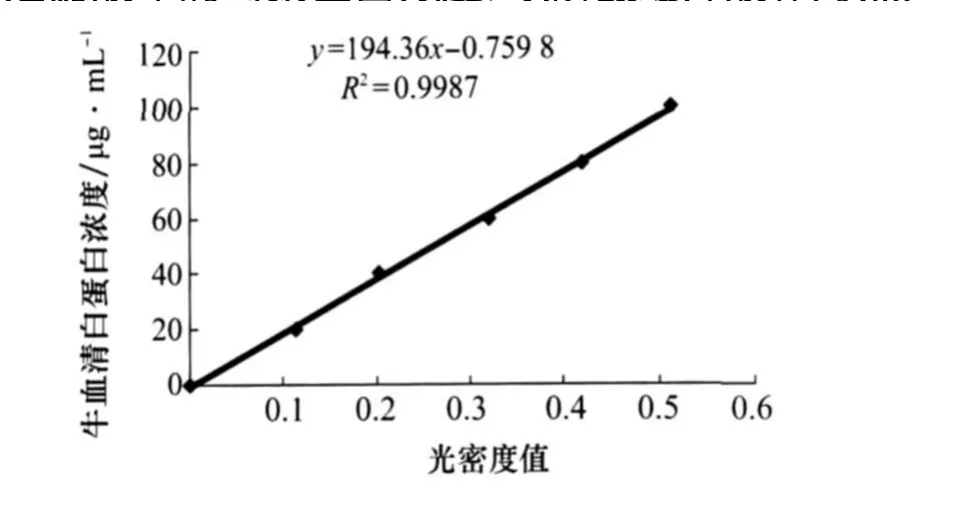
圖1 牛血清白蛋白的標準曲線
1.4 葉霉病菌毒素生物測定
種子萌發法[3]:種子在40℃水中浸泡4 h,用0.1%高錳酸鉀浸泡15 min。將種子放在鋪有濾紙的培養皿中,每皿放50粒,分別各自加入上述7個濃度的毒素液2m L,然后每日加1m L,并透氣1 h,置于28℃培養箱中黑暗培養,以未接菌的培養液作對照。5 d后測其萌發率,計算萌發抑制率。

2 結果與分析
2.1 不同培養液對產毒的影響
由圖2可以看出,培養液對病菌產毒有很大的影響,其中以在PS液體培養液中產毒量最高,并且種子萌發抑制率最高達88%;其次是PD液體培養液;在Czapek、Richard這2種培養液中幾乎不產毒素,說明它們是最不適合產毒的培養液。

圖2 培養液的種類對番茄葉霉病菌產毒的影響
2.2 pH對產毒的影響
由圖3可以看出,培養液pH對病菌產毒有一定的影響,偏酸性的環境有利于產毒,pH為6時,毒性最強,種子萌發抑制率也達到最大為82%。

圖3 pH對番茄葉霉病菌產毒的影響
2.3 培養時間對產毒的影響
由圖4可以看出,從第7天到第21天產毒增長量都很明顯,第28天趨于平緩。雖然第21天的粗毒素蛋白含量比第28天稍低一點,但是它的種子萌發抑制率達到最大為94%,所以21天為最適培養時間。
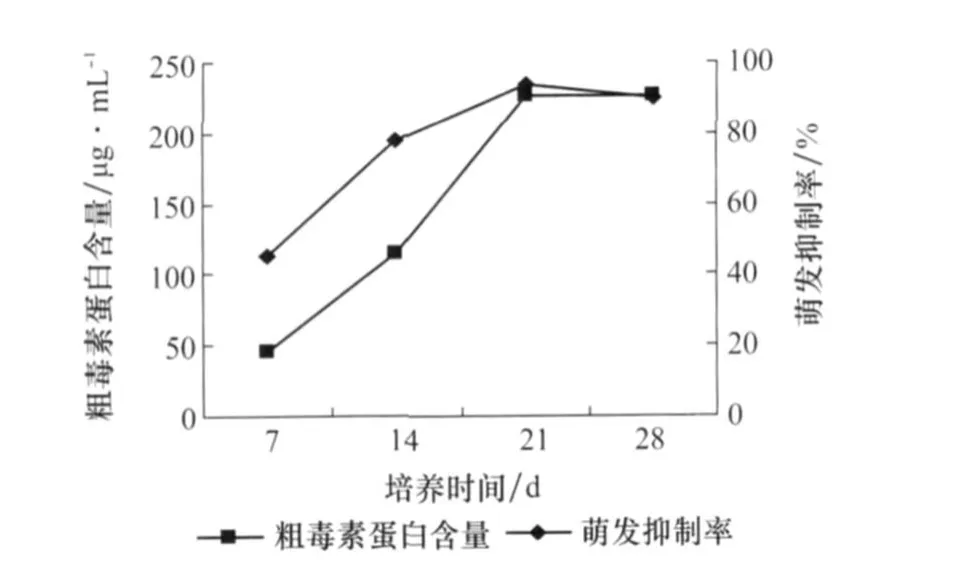
圖4 培養時間對番茄葉霉病菌產毒的影響
2.4 溫度對產毒的影響
由圖5可以看出,溫度對產毒量有一定的影響,溫度偏高或偏低都不利于產毒,以26℃產毒量最多,種子萌發抑制率也最高,這可能跟菌絲產毒時所需酶的活性有關,28、30℃產毒量與種子萌發抑制率僅次于26℃,所以產毒的最佳溫度范圍為:26~30℃。
2.5 培養方式對產毒的影響
由圖6可以看出,全天振蕩培養產毒量與種子萌發抑制率均達到最高。可能是振蕩培養加速了氣體的交換,提供了充足的氧氣,也加速了培養液中營養的利用,從而增強了菌絲的產毒能力;而靜止培養菌落僅僅漂浮于培養液表面生長,無法獲得充足的氧氣與營養,致使產毒的量較低。

2.6 光照對產毒的影響
由圖7可以看出,總的來說光照對產毒的影響不大。光照不但沒有使毒素增加,反而有減少的趨勢,可能光照對產毒有抑制作用,所以本試驗采用黑暗培養。
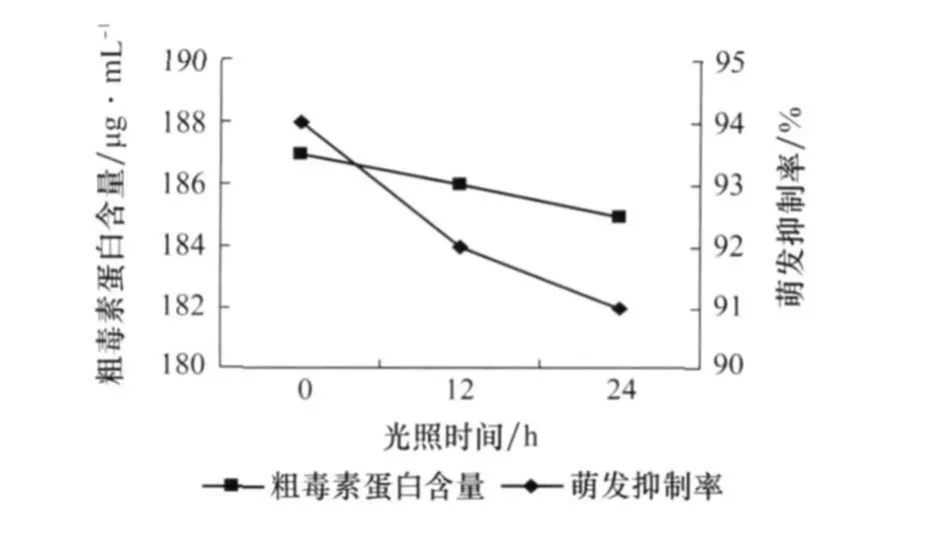
圖7 光照對番茄葉霉病菌產毒的影響
2.7 接菌數量對產毒的影響
由圖8可以看出,接菌數量從1碟到5碟產毒量與種子萌發抑制率逐漸上升,接菌數量為5碟時產毒量與種子萌發抑制率達到最高;7碟時有所降低,其可能原因是菌碟數量太多導致培養液營養供應不足。
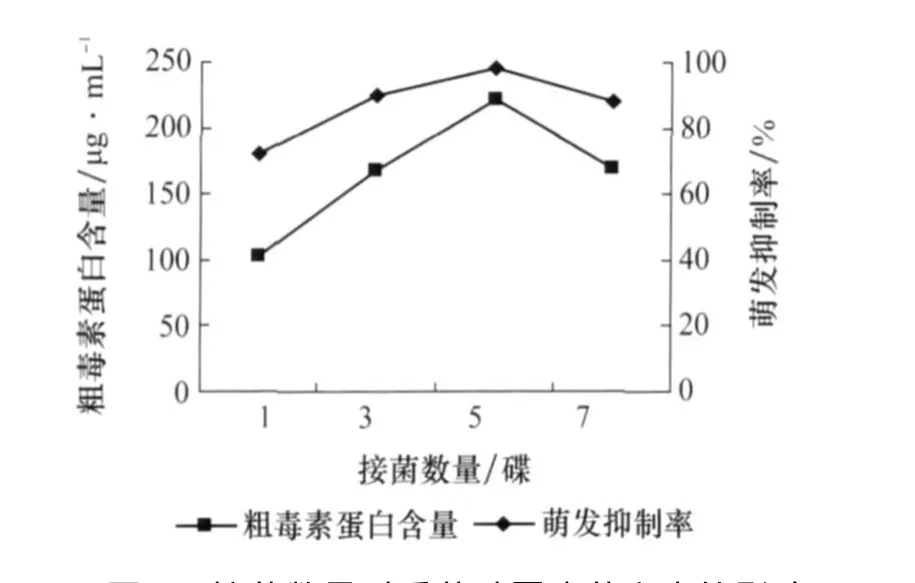
圖8 接菌數量對番茄葉霉病菌產毒的影響
3 討論
培養液對病原菌產毒影響很大,Plich's培養液最適于辣椒疫霉(Phy tophthora capsici)的生長和產毒[11]。高洪敏用4種培養基培養禾谷鐮孢菌,以Richard培養基產生毒素的活性最高[12]。番茄葉霉病菌毒素的產生也同樣受到培養液的影響,試驗結果表明,PS培養液是番茄葉霉病菌的最佳產毒培養液。
在科研和生產中,都需要盡可能地獲得最多的毒素。培養條件如:培養溫度、培養基的pH及培養時間等是影響毒素產量的重要因素。石曉燕等對禾谷鐮刀菌(F.gram inearum)液體培養產毒條件研究表明,禾谷鐮刀菌產毒的最適溫度為20℃,pH為5~6,光照可抑制毒素的產生,一般培養21~28 d達到產毒高峰[13]。劉亞光等[8]得出大豆灰斑病菌(Cerospora sojina)的最適產毒條件為:pH6~7,蔗糖濃度為3%,在25~28℃靜止培養25~27 d。本試驗得到的最適培養條件為:pH為6、溫度為26~30℃、培養時間21 d、黑暗條件下振蕩培養、接菌數量5碟。
許多研究[14-17]表明,各種培養條件分別達到最優,組合起來即為最優培養條件。這為本研究尋找到最優產毒條件提供了理論基礎。
趙曉軍等[18]研究表明,同一菌株毒素對胚芽的抑制率大于胚根。因此,在利用毒素測定不同菌株的生物活性時,采用種子萌發抑制率為宜。所以,本試驗沒有對胚根抑制率進行測定。
多數植物病原真菌可以產生毒素,這些毒素在其致病過程中的作用不同。由于有些植物病原菌分泌的毒素在植物上可以引起與活菌相似的癥狀和病理變化,因此可以替代活菌進行品種的抗性鑒定。這種方法快速、簡單、定量性好,使繁重的田間抗病性鑒定工作可在室內完成[19]。劉慧芹在試驗中證實了中雜8號與其他品種相比較,對毒素的敏感性最小,與其田間抗病性強一致[3]。這表明可以利用葉霉病菌毒素對番茄進行抗病性鑒定。
[1] 陳宇飛.我國番茄葉霉病研究進展[J].東北農業大學學報,2000(12):411-414.
[2] 張滿良.農業病理學[M].中國農業出版社,1997.
[3] 劉慧芹.葉霉病菌粗毒素對番茄的生物活性測定[J].天津農學院學報,2004(11):10-13.
[4] LazarovitsG,Higgins V J.Biological activity and specificity of a toxin produced by Cladospor ium fulvum[J].Physiology and Biochem istry,1979,69(10):1056-1061.
[5] Dow B JM,Callow J A.Partial characterization of glycopeptides from cu lture filtrates of Fulvia fulva(Cooke)Ciferri,the tomato leaf mould pathogen[J].Journal of General M icrobiology,1979,113:57-66.
[6] Tobin L P,Higgins V J.Electrolyte leakage,lipoxygenase,and lipid peroxidation induced in tomato leaf tissue by specific and nonspecificelicitors from Cladospor ium fulvum[J].Plan t Physiol,1989,90:867-875.
[7] 盧鋼,楊悅儉,壽森炎,等.番茄分子育種研究進展[J].分子植物育種,2003,1(5/6):805-811.
[8] 劉亞光.大豆灰斑病菌毒素組分、致病性及其誘導抗性的研究[D].哈爾濱:東北農業大學農學院,2002.
[9] 陳旭升,陳永萱,黃駿麒,等.棉花黃萎病菌致萎峰蛋白氨基酸組分及其有關生化特性分析方法[J].江蘇農業學報,2000,16(1):10-14.
[10]夏正俊,顧本康,吳藹民.毒素法苗期快速檢測棉花品種抗黃萎病性研究[M]∥董金皋,李樹正.植物病原菌毒素研究進展.北京:中國科學技術出版社,1997:200-204.
[11]謝丙炎,牛國仁.辣椒疫霉致病毒素[J].菌物系統,1997,16(4):274-280.
[12]高洪敏,陳捷.玉米莖腐菌毒素的產生條件和化學特征的初步研究[J].沈陽農業大學學報,1999,30(3):223-226.
[13]石曉燕,鄧福友.禾谷鑲刀菌液體培養產毒條件研究初報[J].河北農業大學學報,1992,15(4):34-38.
[14]孫文元,翟玉柱,趙鳳巖,等.草莓灰霉病菌的培養及其毒素的生物測定[J].華北農學報,1999,14(增刊1):112-116.
[15]陸寧海,齊尚紅,吳利民,等.番茄褐斑病菌產毒培養條件及其毒素的致病范圍[J].微生物學雜志,2006,26(4):36-38.
[16]陸寧海,吳利民,田雪亮,等.黃瓜褐斑病菌的產毒培養條件及其毒素的致病范圍[J].遼寧農業科學,2007(3):42-43.
[17]唐樹戈,莊敬華,高增貴,等.玉米彎孢菌毒素培養條件的研究[J].安徽農業科學,2007,35(1):1-3.
[18]趙曉軍,王美琴,劉慧平,等.番茄葉霉病菌不同菌株粗毒素對種子萌發的影響[J].山西農業科學,2005,33(1):50-52.
[19]臺蓮梅.大豆根腐病菌(Fusar ium oxysporum)毒素及其對大豆根部致病作用的研究[D].哈爾濱:東北農業大學農學院,2003.
