長白落葉松幼苗對銨態氮和硝態氮吸收的動力學特征
2010-10-08 05:39:12魏紅旭徐程揚馬履一江俐妮李雪蓮楊卓
植物營養與肥料學報 2010年2期
關鍵詞:生長
魏紅旭,徐程揚*,馬履一,江俐妮,李雪蓮,楊卓
(1北京林業大學林學院,北京100083;2北京林業大學森林培育與保護重點實驗室,北京100083;3吉林市龍潭區江密峰苗圃,吉林132206;4北京林業大學水土保持學院,北京100083)
隨著農業生產集約化的發展,過量施肥特別是氮肥引起的諸多問題是植物營養、土壤、環境等學科關注的熱點問題[1]。據報道,由于不合理施肥使所施氮肥的32%~85%無法被植物吸收利用[2-3]。除了氮素的淋失、揮發等損失外,植物對氮素的吸收、利用、轉化以及根系的吸收動力學特征等因素都會影響氮肥的利用率[4]。
20世紀50年代初,Epstein和Hagen[5]首先將酶促反應動力學方程應用于植物對離子吸收的研究,開創了植物吸收養分動力學研究的先河。70年代以后,不斷有學者對動力學方程進行改進,在這期間確立了經典的以米氏酶促反應動力學方程為基礎的植物根系吸收離子的表達方程[6-7]。近年來國內外在養分吸收動力學研究方面已經開展了大量的工作,但所采用的試驗材料多為水稻、大豆和生菜[8-12]等農作物,關于林業苗木的養分吸收動力學研究的報道相對較少[13-14]。
長白落葉松(Larix olgensis)是中國東北地區速生豐產林的主要樹種,因其材質優良、速生豐產等特點,具有極強的經濟價值和生態效益。生產上常規的長白落葉松養分管理方式十分粗放,不僅造成極大的人力和物力的浪費,而且不利于優質苗木的培育。本研究以當年生春季播種的長白落葉松幼苗為試驗材料,在不同氮源供給的情況下探討苗木對NH+4和NO-3吸收的動力學特征及其動態變化,以期為落葉松的動力學和養分培育方面的研究提供理論依據。
1 材料與方法
1.1 材料準備
試驗材料為東北地區苗木生產普遍采用的當年生長白落葉松苗木,采集于吉林市龍潭區江密峰苗圃(126°45′E,43°45′N),其種源來自吉林省小北湖 。于2008年4月17日進行整地,以生產強度施入基肥后進行播種,播種量為5.5 g/m2,播種方法及出苗期水分管理與生產方式同步。分別于生長初期、速生期和木質化期分3次選取長勢均一的苗木進行收獲,每次收獲量為150株,每次收獲苗木生長情況見表1。將收獲苗木帶回實驗室后洗凈根系表面泥土,用去離子水饑餓處理24 h后用于養分動力學試驗處理。
1.2 試驗設計
不同氮素形態的吸收動力學試驗采用離子耗竭法。設 3個營養液濃度處理:1)NH+4處理[以(NH4)2SO4為氮源];2)NO-3處理[以Ca(NO3)2為氮源];3)NH+4∶NO-3=1∶1 處理(濃度比,以NH4NO3為氮源,每種氮離子濃度與單一氮源時相同)。每處理設置 4 個濃度梯度:0.5、1.0、1.5、2.0 mmol/L,同時設置不供氮處理為對照(CK)。將經饑餓處理的苗木移入已裝有130 mL營養液的瓶中,起始pH為6.5。為減少通氣不均對根系吸收能力的影響,在每瓶中加入0.1 mL 3%的H2O2以提供O2。為防止NH+4硝化,在裝有NH+4的營養液內加入7 μ mol/L的硝化抑制劑(C2H4N4)。每5株苗木為一個處理單位,每處理3次重復,以海綿固定,處理18 h。
1.3 測定方法
每次處理后取出苗木根系,切除地上部分,用吸水紙吸干根系表面水分后立即稱重得到根系鮮重。測定處理后的營養液體積,同時測定其中NH+4、NO-3離子濃度。NO-3離子用直接比色法測定,NH+4離子采用納氏試劑比色法測定[14],測定儀器為Agilent 8543型分光光度計。
1.4 參數計算方法
離子吸收速率[μ mol/(g?h),plant]的計算參考楊洪強等[15]的方法:
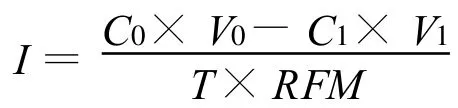
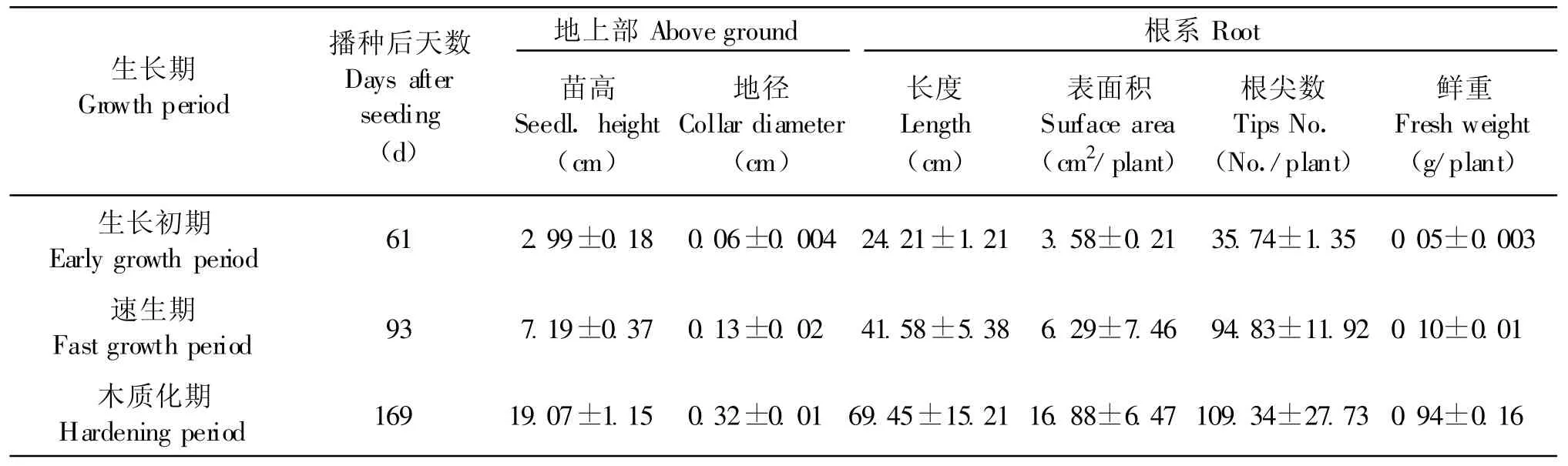
表1 長白落葉松幼苗的生長情況Table 1 Growth parameters of Larix olyensis seedling under different growth periods
式中:C0、C1分別表示處理前后營養液的離子濃度(mmol/L);V0、V1分別表示處理前后營養液體積(L);T為處理時間(h);RFM為根系鮮重(g/plant,FM)。
Michaelis-Menten方程描述的計算采用雙倒數Lineweaver-Burke法[16]:
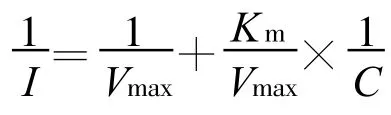
式中:I為離子吸收速率[μ mol/(g?h),plant];C 為處理前離子濃度(mmol/L);Vmax為最大吸收速率[μ mol/(g?h),plant];Km 為米氏常數。 Vmax反映了植物吸收某種離子的最大潛力,其值越大,表明吸收該離子的內在潛力就越大;同時Vmax值還與轉運離子的載體數量和效率成正比[11]。Km為表觀米氏常數,是1/2Vmax時的外液濃度,它表示載體植物根系對離子的親和力,Km值越大,根系與養分離子的親和力越小[13,17]。
輔助分析參數Cmin、α、β的計算分別參考華海霞等[16]和翟明普和蔣三乃[13]所報道的方法,具體如下:
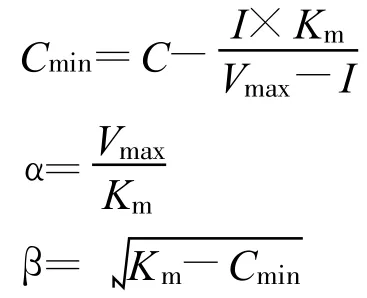
式中:Cmin為離子吸收補償點,表示凈吸收速率為零時的營養液離子濃度,Cmin越小,根系的吸收能力越強[13];α值反映養分流入根系的速率,α值越大,養分離子流入根系的速率越快,反之越慢[13,15];β值是由前人提出的用來研究植物耐瘠薄能力的參數,其值的大小與植物耐瘠薄能力成反比[18]。
為了對比2種形態氮同時存在與單一形態氮存在間的動力學參數差異,引入一新參數Ii,其計算方法為:
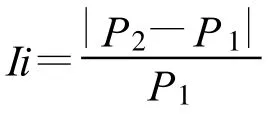
式中:P2為NH+4和NO-3以1∶1濃度比同時存在時某一動力學參數值;P1為NH+4或NO-3單獨存在時某一動力學參數值。兩指標的Ii值差距越大,表明其中一種指標(較大者)對苗木的生理學影響越劇烈。
2 結果分析
2.1 單一氮源時不同生長期苗木對NH+4、NO-3的吸收動力學特征
單一氮源條件下,供試苗木在不同生長階段對NH+4、NO-3的吸收速率隨著NH+4、NO-3濃度的變化規律均符合Michaelis-Menten方程的描述,并且吸收NH+4、NO-3的速率(圖1)和 Vmax、α值(表2)均隨著苗木的生長呈下降趨勢。說明隨著苗木的生長吸收NH+4和NO-3的載體的數量和效率逐漸下降,導致離子流入根系的速率下降。生長初期和速生期,苗木對NH+4吸收的 Vmax、α值(表2)和速率均高于NO-3(圖1),而在木質化期二者的 α值和吸收速率間均無明顯差異。可見,單一氮源下苗木對于NH+4的吸收速率整體上強于NO-3。
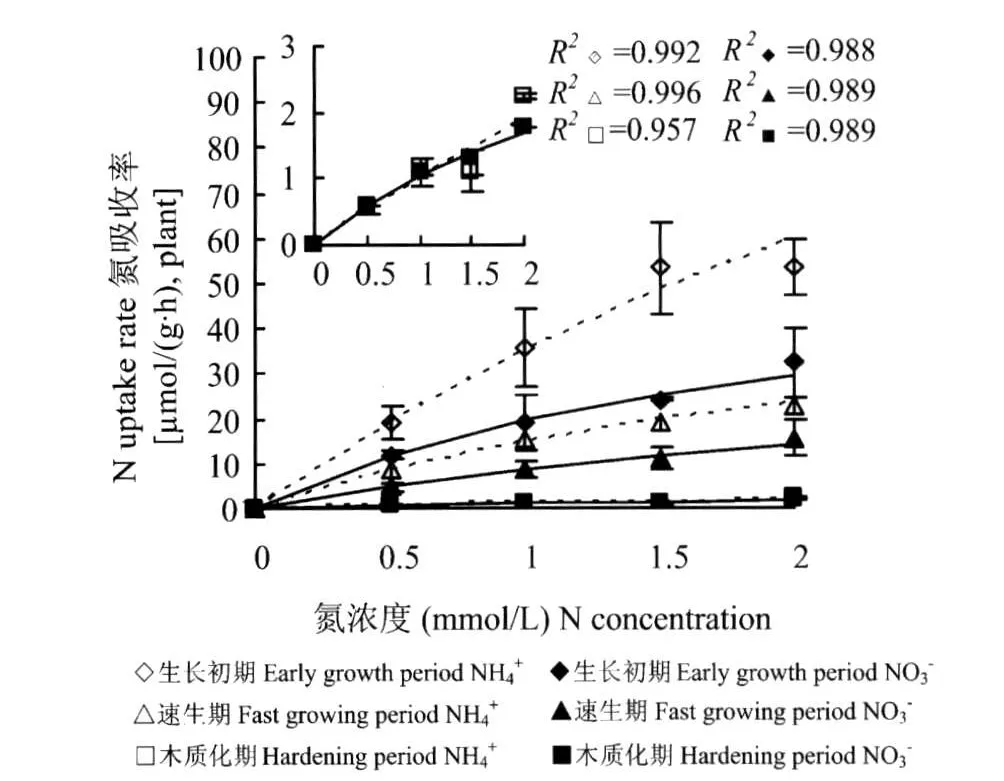
圖1 不同生長期內單一(NH4)2SO4或Ca(NO-3)2供給下苗木對NH+4和NO-3的吸收動力學特征Fig.1 Kinetics of NH+4and NO-3uptakes by seedlings with NH+4or NO-3as N source separately in different growth periods
2.2 NH+4和NO-3共存時不同生長期苗木對NH+4的吸收動力學特征
無論NO-3存在與否,供試苗木在不同生長階段對NH+4的吸收速率隨著NH+4濃度的變化規律均符合Michaelis-Menten方程的描述(圖2),并且NH+4的吸收速率均隨著苗木的生長而下降。與無NO-3時相比,NO-3的加入大幅度降低了苗木在速生期對于NH+4的吸收速率,而并未明顯影響苗木在生長初期和木質化期對于NH+4的吸收速率。
在不同生長階段,苗木吸收NH+4的IiKm均大于IiVmax(表2),說明苗木對NH+4的吸收主要受載體親和力的影響,而并非載體數量和效率。在生長初期和硬化期,IiKm和 IiVmax間差距分別只有4.738和0.001,并且加入NO-3后的Km較單NH+4時分別只提高了76.924和13.463。而速生期的IiKm和IiVmax間差距高達199.464,同時隨著NO-3的加入,Km較單NH+4時提高了682.811。說明在速生期時苗木對于NH+4的吸收受親和力的影響增加,NO-3的加入降低了根系對NH+4的親和力,降低了NH+4流入根系的速率和吸收速率。而在生長初期和木質化期,由于根系對NH+4的親和力較低,NO-3的加入并未對NH+4的吸收產生明顯影響。
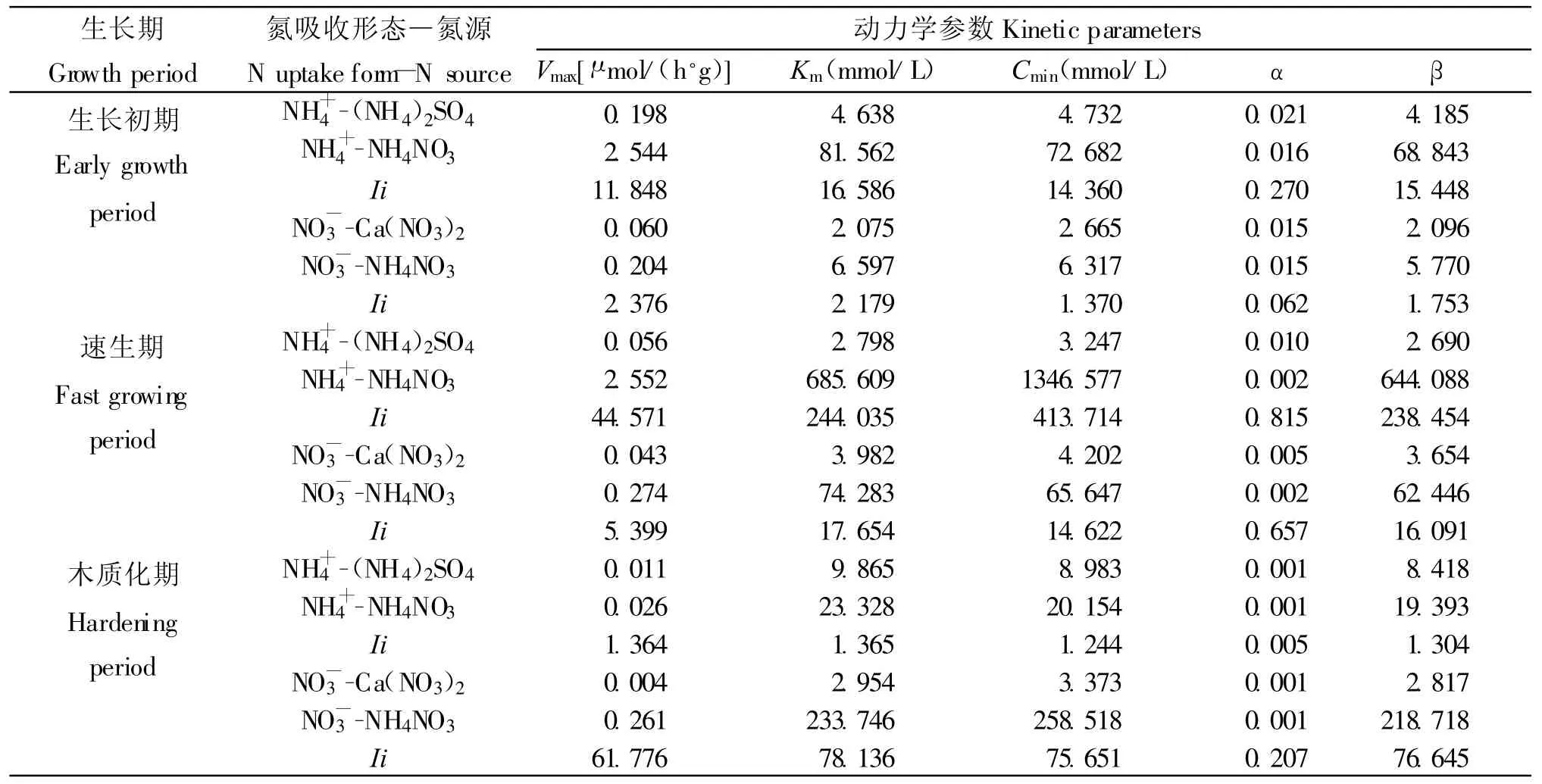
表2 不同生長期內不同氮源的NH+4、NO-3的吸收動力學參數動態Table 2 Dynamics of kinetic parameters of NH+4and NO-3uptakes from different N sources at different growth periods
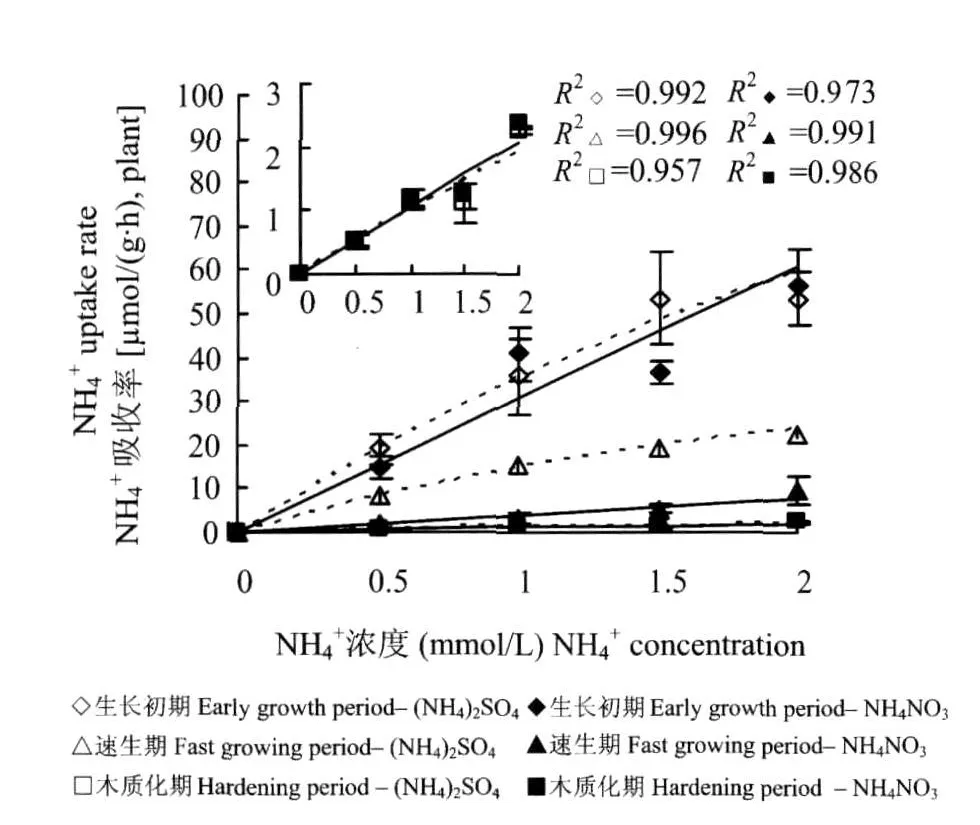
圖2 不同生長期內(NH4)2SO4或NH4NO-3供給下苗木對NH+4的吸收動力學特征Fig.2 Kinetics of NH+4uptake by seedlings with(NH4)2S O4or NH4NO3supplied in different growth periods
2.3 NH+4和NO-3共存時不同生長期苗木對NO-3的吸收動力學特征
無論NH+4存在與否,供試苗木材料在不同生長階段對NO-3的吸收速率隨著NO-3濃度的變化規律也同樣顯著符合Michaelis-Menten方程的描述,并且NO-3的吸收速率隨著苗木的生長而下降(圖3)。與單NO-3為氮源時相比,NH+4的加入(1∶1)降低了苗木在速生期對于NO-3的吸收速率,卻大幅度提高了根系在生長初期對于NO-3的吸收速率,而對于木質化期的吸收促進的影響不明顯。
長白落葉松苗木在生長初期對于NO-3的吸收主要受載體數量和效率影響(IiVmax=2.376;IiKm=2.179),雖然NH+4的加入使得Km提高了4.522(表2),NO-3的吸收速率仍較單NH+4為氮源時有所提高。進入速生期后,隨著NH+4的加入NO-3流入根系的速率的有所下降(α=0.002),導致根系對NO-3的親和力的影響強于載體數量和效率的影響(IiKm-IiVmax=12.255)。因此隨著NH+4的加入,根系對NO-3的親和力的下降導致NO-3的吸收下降。
3 討論與結論
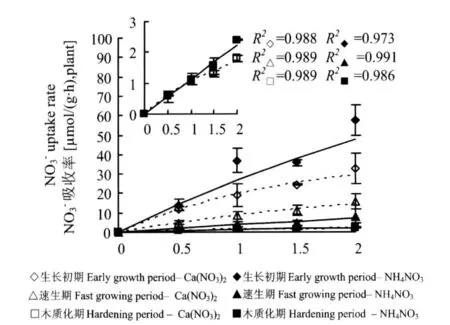
圖3 不同生長期內Ca(NO3)2或 NH4NO3供給下苗木對NO3-的吸收動力學特征Fig.3 Kinetics of NO3-uptake by seedlings with Ca(NO3)2 or NH4NO3supplied in different growth periods
不同的植物種類在離子吸收上往往存在著較大的差異[10]。本次試驗設置的 0~ 2.0 mmol/L的NH+4、NO-3營養液離子濃度在前人關于水稻[19]、生菜[11]和水曲柳[14]等試驗材料的報道中都已被采用過,并證實其與根系吸收離子間的關系符合動力學描述。雖然本次試驗結果亦發現了0~2.0 mmol/L的NH+4、NO-3營養液離子濃度與長白落葉松幼苗吸收速率間關系符合動力學描述(圖1~圖3),但是卻沒有出現前人發現的吸收速率曲線隨著營養液離子濃度的增加而逐漸趨于平緩的規律。這也許是因為長白落葉松苗木對于NH+4、NO-3離子的吸收水平高于前人所采用的植物材料[11,19],也可能是由于前人所采用的材料的準備時間比較短[14],其具體原因有待進一步進行試驗分析。
已有報道指出[19],增NO-3營養液可以促進根系對NH+4的吸收[8-9],但也有認為增加NO-3反而引起了NH+4的吸收速率下降[20]。本研究結果顯示,在速生期,NO-3的加入抑制了載體與NH+4的親和力,明顯降低了NH+4流入根系的速度,從而導致速生期NH+4的吸收速率下降(圖2)。但段英華等[19]則認為,NO-3對NH+4吸收的影響主要在于NH+4載體的運轉速率而非吸收位點與NH+4之間的親和性。前人的研究大多采用水稻等農作物為試驗材料[8-9,19-20],得到的普遍結論是,NO-3的吸收會增加根系細胞內負電荷數量,有利于陽離子的積累[12]。但是,不可忽略的是,植物在吸收還原NO-3的時候需要消耗大量能量。苗木在速生期時由于葉片生長對于NH+4的需求較強[21],和NO-3的吸收還原造成的能量消耗,很有可能通過影響根系中谷氨酰胺合成酶(GS)的合成,進而影響NH+4的同化而降低親和力,影響根系對其的吸收。而段英華等[19]試驗發現,NO-3對于GS的活性具有促進作用,這可能與其測定GS的部位是葉片有關。由于缺少相關試驗結果的支持,以上結論的合理性還需要通過以苗木為試驗材料進一步進行試驗驗證。
有關NH+4影響NO-3的吸收的問題,目前很多研究結果比較統一,即NH+4的存在會明顯抑制根系對NO-3的吸收[8-9,14,17]。王波等[11]將NH+4對NO-3吸收的影響途徑歸納為2種:1)影響控制NO-3載體蛋白合成的基因表達,進而減少細胞膜上載體蛋白的總量;2)NH+4的存在可能影響載體所處細胞膜上的周邊環境,例如膜極化程度的改變[22]、膜結構的改變[23]等。本研究結果顯示,在速生期,NH+4的加入會抑制苗木根系與NO-3的親和力(表2),進而影響NO-3的吸收速率(圖3);但是在生長初期NH+4的加入會提高吸收NO-3的載體數量和效率,因此提高了根系對其的吸收速率。苗木在生長初期相對以根系生長為主,NH+4的加入有利于蛋白的合成,從而提高了吸收NO-3的載體數量,促進其吸收。此結果與王波等[11]報道的結果相反,可能與試驗材料的種類和取樣時期有關。在速生期時,苗木相對以地上部分生長為主,此時根系需要吸收大量的NO-3以滿足地上部分生長的需要。有試驗結果表明,根系在吸收NO-3的過程中要分泌大量的有機酸,降低根際周圍溶液的pH[12]。任軍等[14]認為在水曲柳的培養液中加入NH+4后影響NO-3吸收的原因是由于NH+4和H+的交換降低了根系周圍溶液的pH值,使得根細胞液內OH-濃度升高,不利于NO-3的吸收。本次試驗結果很可能是由于NH+4的加入降低了根際周圍溶液的pH值,影響了有機酸的分泌,進而影響NO-3的吸收。但是,由于缺乏試驗結果的支持,以上推斷還有待進一步論證。
綜上所述,長白落葉松苗木對于NH+4和NO-3的吸收速率隨著生長而下降。當兩種離子同時存在時,長白落葉松苗木在生長初期NH+4會提高根系吸收NO-3的載體數量和效率,從而提高其吸收速率;在速生期,兩種離子會降低彼此與相應載體的親和力,導致彼此的吸收速率都有所下降;在木質化期,由于苗木的生長基本已經停止,根系對于兩種離子的吸收間沒有明顯差異。因此,在長白落葉松苗木的施肥措施中,應以銨態氮肥為主,如磷酸二銨、尿素和硫酸銨等,但是在生長初期可以適當增施硝態氮肥,如硝銨等,以提高苗木對氮素的利用效率。
[1] 王春陽,周建斌,鄭險峰,等.不同栽培模式對小麥-玉米輪作體系土壤硝態氮殘留的影響[J].植物營養與肥料學報,2007,13(6):991-997.Wang C Y,Zhou J B,Zheng X F et al.Effect of different cultivation methods on soil residual nitrate underwinterwheat-summermaize cropping systemp[J].Plant Nutr.Fert.Sci.,2007,13(6):991-997.
[2] Juntunen M L,Hammar T,Rikala R.Leaching of nitrogen and phosphorus during production of forest seedlings in containers[J].Environ.Qual,2002,31:1868-1874.
[3] Juntunen M L,HammarT,Rikala R.Nitrogen and phosphorus leaching and uptake by container birch seedlings(Betula pendula Roth)growth in three different fertilizations[J].New.For,2003,25:133-147.
[4] 孫敏,郭文善,朱新開,等.不同氮效率小麥品種苗期根系的NO3-、NH4+吸收動力學特征[J].麥類作物學報,2006,26(5):84-87.Sun M,Guo W S,Zhu X K et al.Kinetics of nitrate and ammonium uptake by different wheat genotypes at seedling stage[J].J.Trit.Crops,2006,26(5):84-87.
[5] Epstein E,Hagen C E.A kinetic study of the absorption of alkali cations by barley roots[J].Plant Physiol.,1952,27:457-474.
[6] Claassen N,Barber S A.A method for characterizing the relation betweennutrient concentration and flux into roots of intact plants[J].Plant Physiol.,1974,54:564-568.
[7] Baligar V C,Barber S A.Genotypic differences of corn for ion uptake[J].Agron.J.,1979,71:870-873.
[8] Kirk G J D.Plant-mediated processes to acquire nutrients:Nitrogen uptake by rice plants[J].Plant Soil,2001,232:129-134.
[9] Kronzucker H J,Siddiqi M Y,Class A D M et al.Nutrient ammonium synergism in rice:A subcellular flux analysis[J].Plant Plysiol.,1999,119:1041-1045.
[10] 孫小茗,封克,汪曉麗.K+高親和轉運系統吸收動力學特征及其受NH4+影響的研究[J].植物營養與肥料學報,2007,13(2):208-212.Sun X M,Feng K,Wang X L.Kinetics of high affinity system for K+and effects of NH4+[J].Plant Nutr.Fert.Sci.,2007,13(2):208-212.
[11] 王波,賴濤,沈其榮.不同銨硝配比營養液對典型生菜硝酸鹽吸收動力學的影響[J].植物營養與肥料學報,2007,13(6):1098-1104.Wang B,Lai T,Shen Q R.Effects of NH4+-N/NO3--N ratios on kinetics of nitrate uptake by two typical lettuce genotypes in hydropoics[J].Plant Nutr.Fert.Sci.,2007,13(6):1098-1104.
[12] 高祖明,張春蘭,倪金應,等.黃瓜等九種蔬菜與NO3--N親和力的研究[J].南京農業大學學報,1990,13(1):75-79.Gao Z M,Zhang C L,Ni J Y et al.Study on compatibility of several vegerables with NO3--N[J].J.Nanjing Agric.Univ.,1990,13(1):75-79.
[13] 翟明普,蔣三乃.小鉆楊和刺槐根系養分吸收的動力學研究[J].北京林業大學學報,2006,28(2):29-33.Zhai M P,Jiang S N.Dynamics of nutrient absorption in root systems of Populus×xiao zhuanica and Robinia pseudoacacia[J].J.Beijing For.Univ.,2006,28(2):29-33.
[14] 任軍,徐程揚,林玉梅,等.水曲柳幼苗根系吸收不同形態氮的動力學特征[J].植物生理學通訊,2008,44(5):919-922.Ren J,Xu C Y,Lin Y M et al.Kinetic characteristics of different forms of absorbing nitrogen in root system of Fraxinus mandshurica Rupr.seedling[J].Plant Physiol.Commun.,2008,44(5):919-922.
[15] 楊洪強,張連忠,戚金亮,等.蘋果磚木根系鈣素吸收動力學研究[J].園藝學報,2003,30(3):253-257.Yang H Q,Zhang L Z,Qi J L et al.The kinetics of Calcium uptake in apple rootstock roots[J].Acta Hor.Sin.,2003,30(3):253-257.
[16] 華海霞,梁永超,婁運生,等.水稻硅吸收動力學參數固定方法的研究[J].植物營養與肥料學報,2006,12(3):358-362.Hua H X,Liang Y C,Lou Y S et al.Comparison of research methods for silicon uptake kinetics of rice[J].Plant Nutr.Fert.Sci.,2006,12(3):358-362.
[17] 趙越,馬鳳鳴,張多英.甜菜對不同氮素吸收動力學的研究[J].東北農業大學學報,2006,37(3):294-298.Zhao Y,Ma F M,Zhang D Y.Study on the absorption kinetics of different nitrogen in sugar beet[J].J.Northeast Agric.Univ.2006,37(3):294-298.
[18] 蔣廷惠,鄭紹建,石錦琴,等.植物吸收養分動力學研究的幾個問題[J].植物營養與肥料學報,1995,1(2):11-l7.Jiang T H,Zheng S J,Shi J Q et al.Several considerations in kinetics research on nutrients uptake by plants[J].Plant Nutr.Fert.Sci.,1995,1(2):11-17.
[19] 段英華,張亞麗,沈其榮.增硝營養對不同基因型水稻苗期吸銨和生長的影響[J].土壤學報,2005,42(2):260-266.Duan Y H,Zhang Y L,Shen Q R.Effect of nitrate on the ammonium uptake and growth of different genotypes of rice(Oryza sativa)at the seedling stage[J].Acta.Ped.Sin.,2005,42(2):260-266.
[20] 張亞麗,董園園,沈其榮,等.不同水稻品種對銨態氮和硝態氮吸收特性的研究[J].土壤學報,2004,41(6):918-924.Zhang Y L,Dong Y Y,Shen Q R.Characteristics of NH4+and NO3-uptake by rices of different genotypes[J].Acta.Pedol.Sin.,2004,41(6):918-924.
[21] 吳楚,王政權,范志強.氮素形態處理下水曲柳幼苗養分吸收利用與生長及養分分配與生物量分配的關系[J].生態學報,2005,25(6):1282-1291.Wu C,Wang Z Q,FanZ Q.Relationships between nutrient uptake,use efficiency and growth,and between nutrient partitioning patterns and biomass partitioning patterns in Fraxinus mandshurica seedlings supplied with different ratios of nitrogen forms[J].Chin.J.Appl.Ecol.,2005,25(6):1282-1291.
[22] Crawford N M,Glass A D M.Molecular and physiological aspects of nitrate uptake in plants(Reviews)[J].Trends Plants Sci.,1998,3(10):389-395.
[23] Colmer T D,Bloom A J.A comparision of NH4+and NO3-net fluxes along roots of rice and maize[J].Plant Cell Environ,1998,21:240-246.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
