外源NO對銅脅迫下番茄光合、生物發(fā)光特性及礦質(zhì)元素吸收的影響
2010-11-21 07:13:44張義凱韓小嬌金洪陳秀靈崔秀敏吳小賓徐艷如
植物營養(yǎng)與肥料學(xué)報 2010年1期
關(guān)鍵詞:植物
張義凱,韓小嬌,金洪,陳秀靈,崔秀敏*,吳小賓,徐艷如
(1山東農(nóng)業(yè)大學(xué)資源與環(huán)境學(xué)院,山東泰安271018;2中國林業(yè)科學(xué)研究院,北京100091;3國際網(wǎng)絡(luò)竹藤中心,北京100102)
銅(Cu)是植物生長發(fā)育的必需微量元素,參與植物的光合作用等許多過程,在植物的新陳代謝過程中起著重要的作用,但植物組織中過量的Cu會對植物產(chǎn)生毒害作用[1]。目前Cu礦的開采和冶煉廠三廢的排放,含Cu農(nóng)業(yè)化學(xué)物質(zhì)(殺真菌劑,殺蟲劑和化肥)和有機肥(高Cu豬糞,雞糞和廄肥)的施用可使農(nóng)田土壤,特別是溫室土壤含Cu量達(dá)到一般農(nóng)田土壤的幾倍乃至幾十倍[2-4];溫室土壤重金屬污染已變得日益嚴(yán)峻[5-6]。因此,研究重金屬毒害的緩解機理已成為農(nóng)業(yè)可持續(xù)發(fā)展中亟待解決的問題。
一氧化氮(nitric oxide,NO)是最近發(fā)現(xiàn)的生物體內(nèi)的生長調(diào)節(jié)物質(zhì),同時也是一種活性極強的兩性分子,具有毒害和保護細(xì)胞的雙重功效;但NO的這種毒害或保護效應(yīng)與細(xì)胞內(nèi)生理環(huán)境和NO的有效生理濃度、作用部位及細(xì)胞內(nèi)生理環(huán)境不同而異[7-8]。NO廣泛存在于植物組織中,參與植物對各種非生物脅迫的調(diào)節(jié)。據(jù)報道,NO能緩解鹽脅迫對黃瓜、水稻、小麥和玉米的傷害作用[9-12];能通過誘導(dǎo)氣孔關(guān)閉[13]和增強根細(xì)胞質(zhì)膜內(nèi)向K+通道的活性[14]等提高小麥的抗旱性,能緩解Cd脅迫對玉米、水稻、向日葵的傷害[15-16];能緩解Fe、Cu脅迫對玉米、人參、水稻的傷害[17-19];此外還能促進(jìn)Cu脅迫下小麥種子的萌發(fā)[20]。然而,目前關(guān)于NO對Cu脅迫下番茄光合作用和礦質(zhì)元素的研究尚未見報道。為此,開展了NO對Cu脅迫下的番茄光合特性,超微弱發(fā)光(ultraweak luminescence,UWL)、熒光、磷光強度以及礦質(zhì)營養(yǎng)元素吸收的影響研究,為緩解重金屬毒害提供依據(jù)。
1 材料與方法
1.1 試驗設(shè)計
試驗在山東農(nóng)業(yè)大學(xué)溫室內(nèi)進(jìn)行。番茄品種為“玫瑰”(Lycopersicon esculentum Mill.cv.Meigui)。種子經(jīng)55℃溫湯浸種消毒15 min,然后在潤濕的吸水紙上26℃催芽。待種子露白后,播于洗凈的蛭石中,萌發(fā)后用1/4 Hoagland[21]營養(yǎng)液澆灌。當(dāng)幼苗具有3~4片真葉時,挑選生長一致的植株洗凈根部蛭石后,移栽于5 L塑料盆中,用厚度為3 cm的泡沫塑料板做成鍥形蓋子,覆蓋在塑料盆頂部。每盆栽5株,用1/2 Hoagland營養(yǎng)液進(jìn)行栽培。1周后換成完全營養(yǎng)液,此后每3 d更換一次營養(yǎng)液。營養(yǎng)液栽培期間用電動氣泵24 h連續(xù)通氣。當(dāng)植株具有5~6片真葉時,對番茄幼苗進(jìn)行處理。
NO供體硝普鈉[Na2Fe(CN)5]NO,SNP,購自Sigma公司,先用蒸餾水配成200 mmol/L的母液,4℃避光保存,用時按所需濃度稀釋。血紅蛋白(Hb,Sigma公司生產(chǎn),上海生工分裝)為NO的清除劑;其他試劑均為國產(chǎn)分析純。由于SNP分解的產(chǎn)物除NO外,還有NO2-/NO2-和Fe(CN)25-等陰離子[13],在試驗中加入一定濃度的NaNO2/NaNO3和Na3Fe(CN)6(后者也是SNP的相似物,但不能產(chǎn)生NO)作為相關(guān)對照,二者在本試驗條件下均不能產(chǎn)生NO。
試驗設(shè)6個處理:1)CK,Hoagland完全營養(yǎng)液為對照 ;2)50 μ mol/L CuCl2;3)50 μ mol/L CuCl2+100 μ mol/L SNP;4)50 μ mol/L CuCl2+100 μ mol/L NaNO2+100 μ mol/L NaNO3(其中 NaNO2+NaNO3以NOx-代替 ,下同);5)50 μ mol/L CuCl2+100 μ mol/L Na3Fe(CN)6;6)50 μ mol/L CuCl2+100 μ mol/L SNP+0.1%Hb,分別用 CK 、Cu 、Cu+S 、Cu+N 、Cu+F 、Cu+S+H表示,3次重復(fù),在溫室內(nèi)隨機排列。處理期間每天更換營養(yǎng)液,用低濃度KOH或HCl調(diào)節(jié)至pH 5±0.2。溫室內(nèi)光照約12 h,白天最高溫度32℃,夜間最低溫度15℃。處理8 d后,取相同節(jié)位的葉片用于分析測定。
1.2 測定項目與方法
葉綠素含量的測定:取植株中上部功能葉片,用直徑為1cm的打孔器采取葉圓片,加80%的丙酮和少許MgCO3,低溫避光條件下勻漿,離心,采用Arnon的方法測定葉綠素含量[22]。
光合參數(shù)的測定:凈光合速率(Pn)、蒸騰速率(Tr)、氣孔導(dǎo)度(Gs)、胞間CO2濃度(Ci)用英國PP-systems公司生產(chǎn)的CIRAS-1便攜式光合系統(tǒng)測定,測定光強為 800 μ mol/(m2·s)。
超微弱發(fā)光強度的測定:參照張新華等[23]的方法。取葉片鮮重 0.5 g左右,白熾燈[光強 10 μ mol/(m2·s)]照射5 min后立即放入測量杯中測量超微弱發(fā)光。超微弱發(fā)光儀(BPCL)由中國科學(xué)院生物物理研究所研制。測量參數(shù)為:調(diào)節(jié)高壓為800 V,標(biāo)準(zhǔn)光源發(fā)光強度7000 counts/s,本底強度5 counts/s,采樣時間200 s,采樣間隔時間1 s。樣品測定在暗室及恒溫(20±1)℃、恒濕(RH 75%±2%)條件下進(jìn)行。
葉片熒光強度的測定:測定儀器為日立F-4500熒光儀,激發(fā)光EX波長440 nm,發(fā)射光EM波長 500~ 700 nm,縫寬 EX/EM 為 10.0 nm/5.0 nm,掃描速度12000 nm/min,掃描間隔EX/EM為10.0 nm/5.0 nm[24]。
葉片磷光強度的測定:儀器為日立F-4500熒光儀,EX波長210 nm,EM 波長400~500 nm,縫寬EX/EM 10.0 nm/5.0 nm,掃描速度240 nm/min,掃描間隔EX/EM 10.0 nm/5.0 nm[24]。
礦質(zhì)元素的測定:樣品在105℃下殺青30 min,并在65~70℃下烘干至恒重,樣品磨碎過篩,HNO3-HClO4法消煮后,采用FP640型火焰分光光度計測定K,AA370MC型原子吸收分光光度法測定Ca、Fe、Cu 、Zn、Mn等礦質(zhì)元素的含量[25]。
SAS統(tǒng)計軟件對平均數(shù)用Duncan’s新復(fù)極差法進(jìn)行多重比較。
2 結(jié)果與分析
2.1 外源NO對銅脅迫下番茄葉片葉綠素含量的影響
表1看出,50 μ mol/L的Cu處理(Cu)顯著降低番茄葉片的葉綠素a、葉綠素b和葉綠素a+b的含量,降低葉綠素a/b比值,而外施100 μ mol/L的SNP處理(Cu+S)能顯著緩解Cu脅迫下對番茄葉片葉綠素a、葉綠素b和葉綠素a+b的抑制,提高葉綠素a/b比值;在添加NO清除劑血紅蛋白處理(Cu+S+H)后,SNP的緩解效果被消除。在Cu處理液中加入 100 μ mol/L NO-x(Cu+N)或 100 μ mol/L 亞鐵氰化鈉(Cu+F),與Cu脅迫處理差異不顯著。
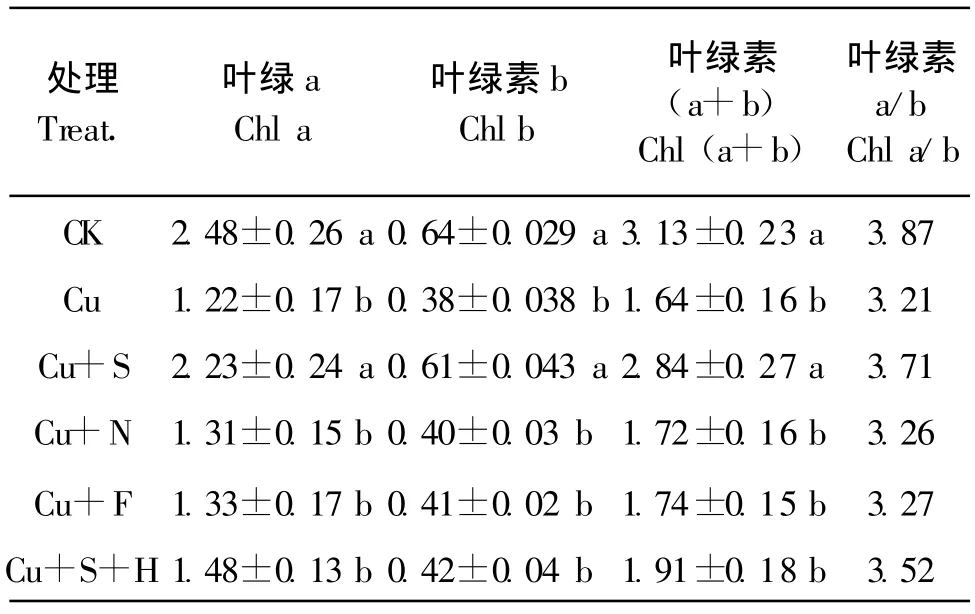
表1 外源NO對銅脅迫下番茄葉片葉綠素含量的影響Table 1 Effects of exogenous nitric oxide on the chlorophyll contents in leaves of tomato under Cu stress(mg/kg,FW)
2.2 外源NO對銅脅迫下番茄葉片光合參數(shù)的影響
由圖1看出,與CK 處理相比,50 μ mol/L的Cu處理8 d番茄葉片的Pn,Tr和Gs顯著降低,番茄葉片Ci顯著上升,而外施100 μ mol/L的SNP處理(Cu+S)能顯著提高Cu脅迫下番茄葉片的Pn、Tr和Gs,降低番茄葉片Ci;用NO清除劑血紅蛋白處理(Cu+S+H)后,顯著消除SNP的緩解效果,而Tr未達(dá)到顯著水平。在50 μ mol/L的Cu處理培養(yǎng)液中加入100μ mol/L NO-x(Cu+N)或 100 μ mol/L 亞鐵氰化鈉(Cu+F),與Cu處理相比,對番茄葉片光合參數(shù)影響差異不顯著。

圖1 外源NO對銅脅迫下番茄葉片凈光合速率Pn、蒸騰速率 Tr、氣孔導(dǎo)度 Gs和胞間 CO2濃度 Ci的影響Fig.1 Effects of exogenous nitric oxide on the net photosynthetic rate(Pn),transpiration rate(Tr),stomatal conductance(Gs)and intercellular CO2concentration(Ci)in leaves of tomato under Cu stress
2.3 外源NO對銅脅迫下番茄葉片生物發(fā)光強度的影響
50 μ mol/L的Cu脅迫處理顯著提高番茄葉片UWL強度、熒光發(fā)光強度和磷光發(fā)光強度,外施100μ mol/L SNP處理(Cu+S)顯著降低UWL、熒光和磷光的發(fā)光強度,然而,添加NO清除劑處理(Cu+S+H)可以顯著消除SNP的效果。在Cu處理液中加入 100 μ mol/L NO-x(Cu+N)或100μ mol/L 亞鐵氰化鈉(Cu+F),與Cu處理相比,對番茄葉片UWL、熒光強度的影響差異不顯著,磷光強度顯著增加(圖2)。
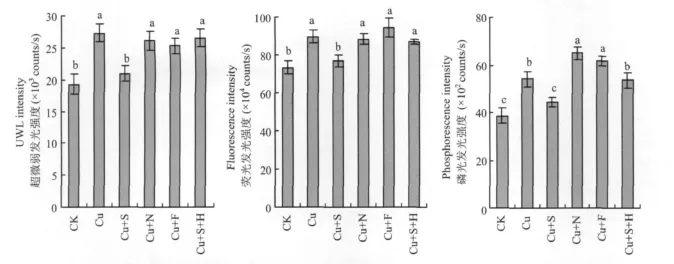
圖2 外源NO對銅脅迫下番茄葉片超微弱發(fā)光強度、熒光強度和磷光強度的影響Fig.2 Effects of exogenous nitric oxide on ultraweak luminescence intensity,fluorescence intensity,phosphorescence intensity in leaves of tomato under copper stress
2.4 外源NO對銅脅迫下番茄植株礦質(zhì)營養(yǎng)元素含量的影響
與CK相比,50 μ mol/L的Cu顯著增加番茄葉片、根系中的Cu含量,顯著降低番茄葉片中Ca、Fe、Zn、Mn和根系中K 、Fe、Zn 的含量 ,然而顯著增加根系中Ca的含量,對葉片中K含量和根系中Mn含量影響不顯著。與Cu脅迫處理相比,施加SNP處理(Cu+S)略微降低葉片、根系中Cu的含量,但未達(dá)到顯著水平;SNP顯著提高葉片中Ca、Fe、Zn、Mn以及根系中K、Fe、Zn的含量,并且顯著降低根系中Ca、Mn的含量;在添加NO清除劑血紅蛋白處理(Cu+S+H)后,SNP的效果被消除。在Cu處理液中加入 100 μ mol/L NO-x(Cu+N)或 100 μ mol/L 亞鐵氰化鈉(Cu+F),與Cu脅迫處理相比,對番茄葉片中K、Ca、Fe、Cu、Zn 、Mn 含量影響不顯著,對番茄根系中K、Ca含量影響差異不顯著;而Cu+N處理顯著降低番茄根系中Fe、Cu、Zn、Mn含量,Cu+F處理顯著增加根系中Fe含量,降低Cu、Mn含量(表2)。
3 討論
葉綠素含量是反映光合強度的重要指標(biāo)。植物受到重金屬脅迫時,各種生理過程都會受到影響,直接或間接地影響到葉綠素的含量。Cu脅迫下,番茄葉片葉綠素含量顯著下降。施加SNP顯著增加番茄葉片葉綠素a、葉綠素 b、葉綠素a+b的含量,提高葉綠素a/b比值(表1);而添加血紅蛋白后,SNP的效果被顯著消除,施用NO-x和Na3Fe(CN)6,均不能減輕Cu脅迫對葉綠素含量的影響。SNP是NO的供體,而NO-x是NO的分解產(chǎn)物,Na3Fe(CN)6是SNP的相似物或分解產(chǎn)物,兩者均不能產(chǎn)生NO,血紅蛋白是NO的清除劑。因此,可以得出SNP釋放出NO,提高番茄葉片葉綠素含量,維持Cu脅迫下番茄較高的光合速率。
光合作用的強弱對于植物生長、產(chǎn)量及其抗逆性都具有十分重要的影響,常作為判斷植物生長和抗逆性大小的指標(biāo)。Cu脅迫下,引起植物葉片光合速率降低的植物自身因素主要有兩類,一是受氣孔導(dǎo)度影響的氣孔限制;二是受葉肉細(xì)胞光合活性的下降影響的非氣孔限制。Farquhar和Sharkey[26]認(rèn)為,Ci值的大小是評判氣孔限制和非氣孔限制的依據(jù)。Pn、Gs和Ci值同時下降時,Pn的下降為氣孔限制;相反,如果葉片Pn的降低伴隨著Ci值的提高,說明光合作用的限制因素是非氣孔限制。本試驗結(jié)果表明,Cu脅迫處理8 d后,番茄葉片的Pn、Gs下降,Ci升高(圖1),說明此時非氣孔限制成為主要因素;而外施NO能顯著減輕Cu脅迫下番茄葉片Pn、Tr和Gs的下降幅度,從而抑制了番茄葉肉細(xì)胞光合活性下降,提高光合速率。

表2 外源NO對Cu脅迫下番茄植株礦質(zhì)營養(yǎng)元素含量的影響(mg/kg,DW)Table 2 Effects of exogenous nitric oxide on mineral contents in tomato under Cu stress
植物UWL與植物體的氧化代謝、信息傳遞、光合作用、細(xì)胞分裂、死亡及生長調(diào)控等基本生命過程存在著內(nèi)在聯(lián)系,也與植物的抗逆性密切相關(guān)[27]。當(dāng)植物受到外界不良環(huán)境刺激時,活性氧在體內(nèi)積累,其中羥自由基與細(xì)胞膜上多聚不飽和脂肪酸通過脂質(zhì)過氧化連鎖反應(yīng),形成單線態(tài)氧,處于激發(fā)態(tài)的單線態(tài)氧退激回到基態(tài)時向外發(fā)射光子,形成UWL[28]。本試驗條件下,Cu脅迫顯著增加番茄葉片UWL強度(圖2),這與活性氧機制[29]相符合。外施NO顯著降低葉片UWL強度,表明NO能增強番茄體內(nèi)活性氧清除機制,保護質(zhì)膜免受重金屬傷害,維持膜結(jié)構(gòu)完整性。Cu脅迫下,番茄的光合作用受到顯著抑制,過多的光能以熒光的形式或熱能耗散,以保護光合作用機構(gòu)免于強光破壞。本研究表明,過量的Cu能顯著提高番茄葉片的熒光強度,表明非化學(xué)能量耗散加強,葉片光能利用率降低。Cu脅迫使光合電子傳遞過程受到抑制[30],光合機構(gòu)受到損害,不利于番茄葉片把所捕獲的光能轉(zhuǎn)變?yōu)榛瘜W(xué)能。而外施NO能顯著降低葉片的熒光強度,緩解Cu毒害對光合機構(gòu)的破壞。磷光是一些處于第一電子激發(fā)三重態(tài)的有機分子發(fā)生輻射躍遷而下降到基態(tài)的各振動能級時產(chǎn)生的。通常情況下,組成蛋白質(zhì)的氨基酸可以產(chǎn)生磷光發(fā)射[31]。在不良環(huán)境下植物葉片內(nèi)的蛋白質(zhì)受到脅迫的“刺激”,體內(nèi)的氨基酸分子會發(fā)生大量的能級躍遷,導(dǎo)致植物葉片磷光強度增加[32]。在Cu脅迫處理8 d后,番茄葉片磷光強度增加,而外施NO能顯著降低葉片磷光強度,表明NO能減輕重金屬Cu對番茄葉片蛋白質(zhì)的毒害。由此可以推斷,NO有可能通過降低重金屬對葉綠體中蛋白質(zhì)的影響,減少葉綠素的分解,保護葉綠體結(jié)構(gòu)和功能,從而提高重金屬脅迫下的光合速率。
重金屬Cu對植物的傷害是多方面的。作為植物必需營養(yǎng)元素,過量的Cu吸收會打破養(yǎng)分平衡,抑制營養(yǎng)元素的吸收和運轉(zhuǎn)。Alva等[33]和黎耿碧等[34]報道,過量的Cu明顯抑制柑橘對N、P、K、Ca、Mg、Fe、Zn、Mn等必需礦質(zhì)元素的吸收;Alaoui-Sossé等[35]認(rèn)為,Cu脅迫顯著減少菠菜葉片中K、Mg、Ca、Fe、Na的含量。本試驗表明,外源NO可顯著提高根系中K、Zn、Fe 的含量以及葉片中Ca、Zn、Fe、Mn 的含量,降低根系中Ca、Mn的含量。然而,NO不能顯著抑制Cu的吸收(表2),表明外源NO可能通過某種機制提高番茄對K、Ca、Fe、Zn、Mn的選擇性吸收,維持胞質(zhì)離子穩(wěn)態(tài),從而緩解Cu的毒害。植物受到Cu毒害時經(jīng)常伴隨失綠癥的出現(xiàn),而失綠癥很可能是由于缺Fe引起。Alva等[33]報道,過量的Cu顯著抑制柑橘對Fe的吸收;Lidon等[36]也報道,過量Cu會抑制水稻對Ca、Fe、Mn等的吸收。Dokiya等[37]在溶液培養(yǎng)中證實,Fe含量增加能夠抑制水稻幼苗對Cu的吸收。Chen等[38]報道,Fe缺乏時能顯著增加鴨跖草中Cu的含量。說明Cu和Fe在一定濃度范圍內(nèi)存在拮抗作用。Graziano等[39]發(fā)現(xiàn),NO對植物葉片中Fe向葉綠體的分配具有重要的作用。本試驗結(jié)果表明,外施NO能顯著增加番茄葉片和根系中Fe、Zn等的含量。由此推斷,外源NO提高葉綠素含量的原因可能是通過提高對Fe、Zn等的吸收和運輸,增加向葉綠體的分配,進(jìn)而提高葉片葉綠素的含量和保護葉綠體結(jié)構(gòu)的完整性,維持番茄較高的光合速率。
[1]Marschner H.Mineral nutrition of higher plants(2nd)[M].London:Academic Press,1995.333-347.
[2]Kaplan M.Accumulation of Cu in soils and leaves of tomato plants in greenhouses in Turkey[J].J.Plant Nutr.,1999,22:237-244.
[3]Brun L A,Maillet J,Hinsinger P et al.Evaluation of copper availability to plants in copper-contaminated vineyard soils[J].Environ.Pollut.,2001,111:293-302.
[4]盧東,宗良綱,肖興基,等.華東典型地區(qū)有機與常規(guī)農(nóng)業(yè)土壤重金屬含量的比較研究[J].農(nóng)業(yè)環(huán)境科學(xué)學(xué)報,2005,24(1):143-147.Lu D,Zong L G,Xiao X J et al.A comparison of heavy metals concentration in soils of organic and conventional farming in typical regions of eastern China[J].J.Agric.Environ.Sci.,2005,24(1):143-147.
[5]黃治平,徐斌,張克強,楊秀春.連續(xù)四年施用規(guī)模化豬場豬糞溫室土壤重金屬積累研究[J].農(nóng)業(yè)工程學(xué)報,2007,23(11):239-243.Huang Z P,Xu B,Zhang K Q,Yang X C.Accumulation of heavy metals in the four years’continual swine manure-applied greenhouse soils[J].Trans.Chin.Soc.Agri.Eng.,2007,23(11):239-243.
[6]曾希柏,李蓮芳,白玲玉,等.山東壽光農(nóng)業(yè)利用方式對土壤砷累積的影響[J].應(yīng)用生態(tài)學(xué)報,2007,18(2):310-316.Zeng X B,Li L F,Bai L Y et al.Arsenic accumulation in different agricultural soils in Shouguang of Shandong province[J].Chin.J.Appl.Ecol.,2007,18(2):310-316.
[7]Beligni M V,Lamattina L.Is nitric oxide toxic or protective?[J]Trends Plant Sci.,1999,4:299-300.
[8]Beligni M V,Lamattina L.Nitric oxide counteracts cytotoxic processes mediated by reactive oxygen species in plant tissues[J].Planta,1999,208:337-344.
[9]Shi Q H,Ding F,Wang X F,Wei M.Exogenous nitric oxide protect cucumber roots against oxidative stress induced by salt stress[J].Plant Physiol.Biochem.,2007,45:542-550.
[10]阮海華,沈文飆,葉茂炳,徐朗萊.一氧化氮對鹽脅迫下小麥葉片氧化損傷的保護效應(yīng)[J].科學(xué)通報,2001,46(23):1993-1997.Ruan H H,Shen W B,Ye M B,Xu L L.Protection of nitric oxide on salt induced membrane oxidation inwheat leaves[J].Bull.Sci.,2001,46(23):1993-1997.
[11]張艷艷,劉俊,劉友良.一氧化氮緩解鹽脅迫對玉米生長的抑制作用[J].植物生理與分子生物學(xué)學(xué)報,2004,30(4):455-459.Zhang Y Y,Liu J,Liu Y L.Nitric oxide alleviates growth inhibition of maize seedlings underNaCl stress[J].J.Plant Physiol.Mol.Biol.,2004,30(4):455-459.
[12]Uchida A,Jagendorf A T,Hibino T et al.Effectsof hydrogen peroxide and nitric oxide on both salt and heat stress tolerance in rice[J].Plant Sci.,2002,163:515-523.
[13]Mata C G,Lamattina L.Nitric oxide induces stomatal closure andenhances the adaptive plant responses against drought stress[J].Plant Physiol.,2001,126(3):1196-1204.
[14]聞玉,趙翔,張驍.水分脅迫下一氧化氮對小麥幼苗根系生長和吸收的影響[J].作物學(xué)報,2008,34(2):344-348.Wen Y,Zhao X,Zhang X.Effects of nitric oxide on root growth and absorption in wheat seedlings in response to waterstress[J].Acta A-gron.Sin.,2008,34(2):344-348.
[15]Hsu Y T,Kao C H.Cadmium toxicity is reduced by nitric oxide in rice leaves[J].Plant Growth Regul.,2004,42:227-238.
[16]Laspina N V,Groppa M D,Tomaro M L,Benavides M P.Nitric oxide protects sunflower leaves against Cd-induced oxidative stress[J].Plant Sci.,2005,169:323-330.
[17]敬巖,孫寶騰,符建榮.一氧化氮改善鐵脅迫玉米光合組織結(jié)構(gòu)及其活性[J].植物營養(yǎng)與肥料學(xué)報,2007,13(5):809-815.Jing Y,Sun B T,Fu J R.Nitric oxide improves photosynthetic structure and activity in iron-deficient maize[J].Plant Nutri.Fert.Sci.,2007,13(5):809-815.
[18]Yu C C,Hung K T,Kao C H.Nitric oxide reduces Cu toxicity and Cu-induced NH4+accumulation in rice leaves[J].J.Plant Physiol.,2005,162:319-330.
[19]Tewari R K,Hahn E J,Paek K Y.Modulation of copper toxicity-induced oxidative damage by nitric oxide supply in the adventitious roots of Panax ginseng[J].Plant Cell Rep.,2008,27:171-181.
[20]Hu K D,Hu L Y,Li Y H et al.Protective roles of nitric oxide on germination and antioxidant metabolism in wheat seeds under copper stress[J].Plant Growth Regul.,2007,53:173-183.
[21]Hoagland D R,Arnon D I.The water-culture method for growing plants without soil[J].Calif.Expt.Sta.Circ.,1950,347:1-39.
[22]Arnon D I.Copper enzymes in isolated chloroplasts,polyphenol oxidase in Brta vulgaris[J].Plant Physiol.,1949,24:1-15.
[23]張新華,楊洪強,李富軍,張偉.杏花開放過程中超弱發(fā)光和ATP及活性氧含量的變化[J].植物生理與分子生物學(xué)報,2004,30(1):41-44.Zhang X H,Yang H Q,Li F J,Zhang W.Changes in ultraweak luminescence,ATP and active oxygen contents during apricot florescence[J].J.Plant Physiol.Mol.Biol.,2004,30(1):41-44.
[24]楊一心,趙天成,白晉濤,等.丙氨酸咪唑釤的三維熒光激發(fā)和發(fā)射光譜[J].光子學(xué)報,2002,31(9):1165-1168.Yang Y X,Zhao T C,Bai J T et al.Three-dimension fluorescence excitation and emission spectrum of alanine imidazole Sm compound[J].Acta Photon.Sin.,2002,31(9):1165-1168.
[25]中國土壤學(xué)會.土壤農(nóng)業(yè)化學(xué)分析方法[M].北京:中國農(nóng)業(yè)科技出版社,2000.315-336.Soil Science Society of China.Analyses method of soil and agricultural chemistry[M].Beijing:China Agricultural Science andTechnology Press,2000.315-336.
[26]Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Ann.Rev.Plant Physiol.,1982,33:317-3451.
[27]王維江,韓俊英.生物超弱發(fā)光機制及其檢測方法研究進(jìn)展[J].廣東工業(yè)大學(xué)學(xué)報,2000,17(1):49-54.Wang W J,Han J Y.Advance in the study of ultra-weak bioluminescent mechanisms and detecting methods[J].J.Guangdong Univ.Tech.,2000,17(1):49-54.
[28]Foyer C H,Lopez-Delgado H,Dat J F,Scott I M.Hydrogen peroxide-and glutathione-associated mechanisms of acclimatory stress tolerance and signalling[J].Physiol.Plant,1997,100:241-254.
[29]Boveris A,Cadenas E,Chance B.Ultraweak chemiluminescence:a sensitive assay for oxidative radical reactions[J].Fed.Proc.,1981,40:125-128.
[30]Maksymiec W,Wójcik M,Krupa Z.Variation in oxidative stress and photochemical activity in Arabidopsis thaliana leaves subjected to cadmium and excess copper in the presence or absence of jasmonate and ascorbate[J].Chemosphere,2007,66:421-427.
[31]魏玉霞,董川.磷光分析法在生命科學(xué)中的應(yīng)用新進(jìn)展[J].生命的化學(xué),2003,23(4):320-322.Wei Y X,Dong C.Applicationprogress of phosphorimetry analysis in life science[J].Chem.Life,2003,23(4):320-322.
[32]國穎,楊洪強.高溫對平邑甜茶幼苗生物發(fā)光與能量代謝的影響[J].園藝學(xué)報,2008,35(1):99-102.Guo Y,Yang H Q.Effects of high temperature on bioluminescence and energy metabolism of Malus hupehensis(Pamp)Rehd.var.pingyiensis Jiang[J].Acta Hort.Sin.,2008,35(1):99-102.
[33]Alva A K,Chen E Q.Effects of external copper concentrations on uptake of trace elements by citrus seedlings[J].Soil Sci.,1995,159:59-64.
[34]黎耿碧,陳二欽,Alva A K.外界銅離子濃度對柑桔小苗常量元素吸收特性的影響[J].廣西農(nóng)業(yè)大學(xué)學(xué)報,1996,15(3):195-201.Li G B,Chen E Q,Alva A K.Effects of external copper concentrations on uptake of routine elements by citrus seedling[J].J.Guangxi Agric.Biol.Sci.,1996,15(3):195-201.
[35]Alaoui-Sossé B,Genet P,Vinit-Dunand F et al.Effect of copperon growth in cucumber plants(Cucumis sativus)and its relationships with carbohydrate accumulation and changes in ion contents[J].Plant Sci.,2004,166:1213-1218.
[36]Lidon F C,Henriques F S.Copper toxicity in rice:diagnostic criteria and effect on tissue Mn and Fe[J].Soil Sci.,1992,54:130-135.
[37]Dokiya Y,Owa N,Mitsui S.Comparative physiological study of iron,manganese and copper adsorption by plantsⅢ.Interaction between Fe,Mn and Cu onthe absorptionof elementsby rice and barley seedlings[J].Soil Sci.Plant Nutr.,1968,14:169-174.
[38]Chen Y,Shi J,Tian G et al.Fe deficiency induces Cu uptake and accumulation in Commelina communis[J].Plant Sci.,2004,166:1371-1377.
[39]Graziano M,Beligni M V,Lamattina L.Nitric oxide improves internal iron availability in plants[J].Plant Physiol.,2002,130:1852-1859.
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
