全公豬肉膻味的調控
2010-11-22 00:51:22柳東陽朱桂姿編譯
豬業科學 2010年12期
柳東陽,朱桂姿 編譯
(1.華中農業大學動物醫學院,湖北 武漢 430070;2.河南省安陽縣職業中專,河南 安陽 455112)
氣味是肉品的重要感官特性,決定了消費者是否購買。豬肉氣味很大程度上受到雄性膻味的影響,目前認為膻味主要由雄烯酮(androstenone)和糞臭素(skatole)構成。傳統上對生產豬肉的小公豬進行閹割,目的在于增加胴體中的脂肪含量、減少攻擊性和性行為、降低產生膻味的危險。現在消費者傾向于購買瘦肉,閹割得以保留原因主要是降低膻味和減少公豬攻擊性。但閹割的優越性也因消費者對豬肉生產中動物福利的關注而受到質疑。此外,因改進了飼料轉化率和提高了瘦肉率,飼養全公豬的效益更高。與閹豬相比,全公豬多聚不飽和脂肪酸和蛋白含量高,在營養上具有優勢。
歐洲食品安全局(EFSA 2004)報告不同研究者對膻味的描述是:令人不愉快的氣味、像尿味、獸味、汗味和糞味。歐洲六國一個研究小組發現:雄烯酮與尿味有關;糞臭素與糞臭味有關,還有少許萘的味道。即使通過大量測試和統計學對參與者的描述偏差做以糾正,膻味的感覺特性仍然非常復雜。原因可能是培訓中缺乏對膻味有一個統一的定性和定量標準。依據個人敏感度,對于膻味的反應差異很大。豬肉膻味取決于膻味化合物合成和清除的速度。這些過程受到一系列相關代謝酶類表達的影響。
1 雄烯酮的合成與代謝
雄烯酮有較強的脂溶性,睪丸中合成后一部分進入血液,再進入脂肪,加熱時產生膻味。一部分進入唾液,作為性外激素發揮作用。最后一部分進入肝臟經過一期和二期降解分泌入膽汁,經糞便排泄。
1.1 雄烯酮的合成
雄烯酮與其他睪丸同化激素一樣產生于公豬睪丸間質細胞,此外腎上腺和卵巢中也可合成,因此母豬和閹豬血液中也可以檢測到雄烯酮。細胞色素P450C17(CYPI7A1)和細胞色素b5(CYPB5)在合成雄烯酮時起著重要作用。雄烯酮來源于17-乙炔睪酮,經andien-β-合成酶合成。雄烯酮和其他睪丸激素的合成受控于下丘腦——垂體——性腺軸,在出生后2~4 周,下丘腦——垂體——性腺軸有一個短暫的活化,增加了循環中雄烯酮的含量。青年公豬雄烯酮合成較少,隨著性成熟而逐漸增加。神經內分泌系統對青春期公豬睪丸間質細胞合成雄烯酮具有很大的決定作用。遺傳背景對合成雄烯酮的強度也有很大影響,Bonneau 等于1987 年鑒定出了血漿中睪酮和黃體生成素(LH)含量正常,而脂肪中雄烯酮含量較低的成年公豬。
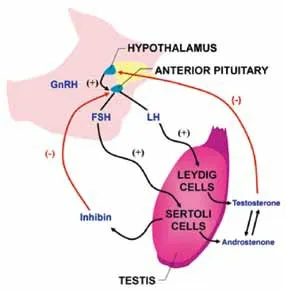
圖1 雄烯酮合成調節示意圖
1.2 雄烯酮的代謝
雄烯酮代謝主要在睪丸和肝臟,由3α-羥化類固醇脫氫酶、3β-羥化類固醇脫氫酶和二期結合酶(羥化類固醇磺基轉移酶:SULT2A1 或 SULT2B1 和UDP-葡萄糖苷轉移酶UGTs)參與。
在睪丸中合成的雄烯酮立即由羥化類固醇磺基轉移酶(SULT2A1/SULT2B1)連上磺基。體內和體外試驗表明,雄烯酮的代謝物為3α-雄烷醇和3β-雄烷醇,最后雄烷醇代謝為極性更強的結合型類固醇。
肝臟與睪丸中雄烯酮代謝不同的是,產生的以上2 種代謝物的百分比不同。雄烷醇經過二期代謝生成葡萄糖苷結合物和磺基結合物。羥化類固醇磺基轉移酶是肝臟中16- 雄烯酮類固醇代謝的關鍵酶。最近發現,SULT2B1 也參與其中。此外,雄烯酮也可以通過SULT2B1 對第3 位的酮基烯醇化而加上一個磺基結合基團。Sinclair and Siquires(2005)等報道一些公豬的血漿中雄烯酮大部分是磺基化的,而Zamratskaia(2007)等報道稱磺基化的雄烯酮要比游離的雄烯酮低很多。同樣, Tuomola(1997)采用47%和100%的甲醇來分離結合和游離的雄烯酮,并未發現在血漿中有任何結合型的雄烯酮。今后應更進一步研究雄烯酮結合磺基之后對其消除和脂肪中積累的影響。
一部分雄烯酮可以經循環進入到頜下 腺。Both(1987)用H3標 記3α-雄烷醇,轉化為雄烯酮,雄烯酮轉化為3β-雄烷醇,雄烷醇保持不變。
2 糞臭素的合成與代謝
糞臭素有脂溶性和水溶性,在大腸合成吸收進入血液,一部分轉入肝臟經一期和二期代謝,經膽汁排泄。另一部分進入脂肪產生膻味。
2.1 糞臭素生物合成
糞臭素來源于L-色氨酸,在豬大腸中合成。色氨酸首先轉變為3-吲哚乙酸,隨后在大腸桿菌和梭菌的某些種的作用下產生吲哚-3-乙酸,之后乳酸桿菌轉變其為糞臭素(3-甲基吲哚)。因此糞臭素最終數量取決于色氨酸的利用度和腸道中菌群組成和活力。色氨酸主要由腸中的黏液細胞轉化產生。一部分糞臭素由糞便排出,一部分經過腸道吸收入血。全公豬脂肪中的糞臭素含量很高,不同性別豬糞中糞臭素含量卻接近一致。
2.2 糞臭素的代謝
肝臟代謝分為兩步。首先加上1 個羥基,也可在二期代謝加上羥基。形成結合物增加了水溶性,促進其經尿液和膽汁排泄,減少脂肪組織的吸收。一期代謝由位于肝細胞內質網上含有血紅蛋白的細胞色素P450 同工酶Ⅱ型(CYP2E1,CYP2A)參與。同樣,CYP2E1,CYP2A 和醛氧化酶也參與了對糞臭素的氧化代謝,CYP2E 為主要酶。
Diaz(1997)采用肝微粒體技術確定了糞臭素代謝的一系列代謝物,定量化為:3-羥基-3-甲基吲哚(45%),其他為:5-羥基-5-甲基吲哚,6-羥基-3 甲基吲哚,3-甲羥基吲哚,吲哚-3- 甲 醇,和2- 氨 基- 苯 乙 酮。這些代謝產物有些可以作為二期代謝底物加上硫酸鹽和葡萄糖醛酸。血漿中高濃度的6-磺基糞臭素可以作為一個指標來判斷公豬具有快速代謝和排泄糞臭素的能力,但仍需要進一步研究。苯磺酸轉移酶具有熱穩定性。對糞臭素代謝中酶類的修飾和表達也許是促進代謝降低膻味的一個有用方法。
糞臭素在肝臟的代謝對于其在脂肪中蓄積有著很大影響。CYP2E1,CYP2A 活力較高的成年公豬脂肪中的糞臭素的含量較低,反之則較高。關于CYP2E1,CYP2A 在糞臭素代謝中作用仍存在爭論,也許與研究中采用的動物的年齡不同有關。其他研究發現,糞臭素在青年公豬中的代謝與成年公豬有所不同。
3 雄烯酮、糞臭素和動物的激素狀態
雄烯酮的合成與其他睪丸激素一樣,受到下丘腦——垂體——性腺軸的控制。因此,在青春期雄烯酮與其他睪丸類固醇類激素數量快速增加。隨著睪丸類固醇增加,糞臭素的量也在增加。可能是睪丸類固醇對肝糞臭素代謝加以調控。雌激素含量與糞臭素的水平也存在正相關聯系。
體外實驗發現,雄烯酮是一個潛在的對CYP2E1 CYP2A 的抑制子。肝臟中CYP2E1 的啟動子受到轉錄因子COUP-TF1 和HNF-1 的活化。Sinclair(2006)發現活化組成型雄甾醇受體(CAR)、孕酮X 受體(PXR)和法尼醇X 受體(FXR)等核受體的配體增加了睪丸間質細胞中SULT2A1的表達。Gillberg(2006)發現原代豬肝臟細胞CAR 受體的活化增加了CYP2A 的活力。因此,似乎糞臭素與雄烯酮的代謝途徑受到這些核受體的共同調節。也許是雄烯酮的一些代謝產物與這些核受體結合而影響到糞臭素代謝相關酶的基因表達。

圖2 閹割
4 對膻味的調控
4.1 免疫閹割對雄烯酮生物合成的修飾
免疫閹割可以消除促性腺激素(GnRH),減少睪丸類固醇的產生,除了可以減少雄烯酮的合成,還可以減少生殖器官的尺寸、精子數目和進攻性。此外,糞臭素和吲哚的含量也可以下降。輝瑞公司的ImprovacTM疫苗可以進行2 次免疫,2 次之間間隔4 周以減少膻味。免疫閹割抑制了睪丸類固醇的產生,使得肝臟中對糞臭素的代謝活力提高,減少脂肪組織中糞臭素蓄積。同時IGF-1的含量下降,也減少了糞臭素的合成。因此通過免疫閹割,既使肉品質提升又可生產全公豬肉。當進行圈養時,宰前的體重過大而不易免疫操作,因此在第2 次免疫注射和屠宰之前有更長的間隔期是最好的。免疫閹割的一個負面影響導致內源性類固醇下降,豬的生長率和瘦肉率下降。從安全的角度考慮,理想的疫苗注射一次最好。
4.2 通過日糧對糞臭素合成和代謝調控
總的來說,脂肪中的糞臭素的量依賴于腸道中產生的數量、腸道停留時間、小腸吸收和肝臟的代謝。前三者通過調節日糧組成可以加以調控,目前對這個日糧組分的模型正處于研究之中。以前研究發現,日糧中一定的碳水化合物影響著腸道中的微生物區系和腸道的功能,反過來又誘導腸道中產生糞臭素的變化。未消化的碳水化合物可以影響增加糞的濕重和干重,減少糞便在腸道停留時間,減少大腸對糞臭素的吸收。腸道吸收糞臭素量依賴于在水中溶解度和腸道停留時間,然而人們對糞臭素從腸道吸收的機制知之甚少。
菊苣多糖和粗制土豆淀粉(RPS)在豬的日糧中可降低組織和糞中的糞臭素的水平。碳水化合物經腸道內微生物區系加以發酵利用,具有對動物保健和調控肉品的潛在功能。先前研究認為,飼喂RPS 減少吲哚是因為抑制了大腸中細胞凋亡,減少了對色氨酸的利用。RPS對同種動物的糞臭素和吲哚產生影響不同,說明了RPS 除了影響凋亡之外,還影響到其他因素。這些因素也許影響了糞臭素的合成,而對于吲哚的影響甚少。
日糧可以影響到肝臟中CYP 的同工酶,后者隨即又影響底物代謝。然而,日糧與糞臭素之間的關系并沒有確定。Zamratskaia(2005,2006) 報 道全公豬飼喂RPS 或者不飼喂,CYP2E1和CYP2A 的活力是相同的。Whiteer(2004)報道含有菾菜根的日糧增加了CYP2E1 的表達水平。
4.3 提前宰殺
全公豬在年輕時及體重較低時宰殺可以減少膻味。因不同品種的青春期不同,即使同一品種的不同個體之間,造成的研究結果也不相同。Zamratskaia(2005)研究發現90 kg 和115 kg 屠宰的豬雄烯酮沒有差別。Chen(2007)發現在體重較大的豬體內脂肪中的雄烯酮較高。Nicoke(2007)發現較重豬脂肪中兩種膻味物質的含量較高。Aldalfaixan(2005)研究杜長大三元組合豬在75 kg 時宰殺,并不能完全得到消除膻味的豬肉。這些研究提示降低屠宰活重并不能完全消除膻味。

圖3 豬群
5 未來的研究方向
5.1 膻味化合物
雄烯酮和糞臭素在組成膻味中特別重要,但膻味的組成不單限于這2 種化合物,其他如:吲哚、雄烷醇、苯甲亞基丙酮和短鏈脂肪酸等尚需要確定。
還要建立對膻味進行有效和一致感覺評估的標準。目前尚缺乏膻味的閾值標準,尤其是雄烯酮。通常使用糞臭素的閾值為0.2~0.25μg/g,雄烯酮為0.5~1.0μg/g。雄烯酮閾值這么大的變化是因為不同消費者對于膻味的感覺能力不同。因此雄烯酮的閾值范圍應該不依賴于人群的差異和檢測方法的不同。為了確定膻味感覺閾值,最根本是要有一個基于標準檢測程序的檢測結果。
5.2 膻味的生物化學
糞臭素在大腸中的合成已經進行了很多研究,但糞臭素產生的微生物機制仍然知之不多。研究產生高或低糞臭素豬的腸道微生物區系,有助于了解色氨酸在大腸內是怎樣降解,以便于我們進一步操控微生物環境以減少糞臭素的產生。CYP2E1 和CYP2A 2 種酶在糞臭素的代謝中起著關鍵作用,要進一步調查各種睪丸類激素對糞臭素在肝臟代謝影響。此外,雄烯酮形成磺基結合物及其對于雄烯酮在脂肪中消除和積累也應進一步研究。
5.3 膻味的遺傳學
要慎重采用在其他動物中評估動物性成熟的辦法來評估公豬膻味的遺傳潛能。可以從現有商用品種中采集大量數據來解釋遺傳標記SNPs(single nucleotide olymorphisms,單核苷酸多態性)與膻味之間的聯系。更重要的是確定膻味數量性狀基因座,確定這些基因座可以用來比對染色體上膻味基因的位置。采用具有相同類固醇產生潛力,而表現型具有高或低膻味的豬來進一步對基因表達做研究,以確定候選基因。最后,也應對膻味物質代謝整合過程研究進行支持,這些研究可以發現更多的候選基因。
5.4 免疫閹割
免疫閹割與全公豬的成本效益分析尚未有人進行對比。進行此實驗要消耗人工、材料和大量的免疫費用。免疫閹割的時間是否可以比現在推薦的時間更早一些?免疫閹割對豬肉品質影響的研究甚少,僅有幾篇文獻報道了保水能力和色澤。
5.5 膻味檢測方法
屠宰場內快速檢測膻味物質面臨著挑戰。現有技術要么處于開發階段,要么需要高素質技術人員來操作。理想的技術應該在較短時間內(幾秒~幾分鐘)就應得到結果。很多情況下,采樣的時間耗去了實驗的大部分時間。另外,現有的檢測方法昂貴,能效比決定著業界是否采用新的檢測技術。
理想技術的檢測結果是:區分可接受樣品和不可接受樣品應該是百分之百的正確。鑒于不同國家的市場情況,低的假陽性可以接受,但假陰性必須為零。指紋印跡檢測技術仍然有太高的假陰性(5%~10%)。目前沒有一種適用于各種屠宰條件的技術。還需要開發相應的工作軟件,要考慮到投入產出比、自動化、高通量,取樣簡單易行。
本文摘編自:
K. Lundstrom,K.R.M atthews,J.E.Haugen.Pig meat quality from entire males.Animal,2009,11(3):1497-1507.
G.Zamaratskaia,, E. J. Squires.Biochemical,nutritional and genetic effects on boar taint in entire male pigs.Animal,2009,11(3):1508-1521.
