太平洋牡蠣雜交三倍體與誘導三倍體的生長比較*
2011-01-10 09:41:26王昭萍于瑞海張躍環李曉瑜李雅琳郭希明
中國海洋大學學報(自然科學版) 2011年7期
關鍵詞:生長
孔 靜,王昭萍**,于瑞海,張躍環,劉 劍,李曉瑜,李雅琳,郭希明
(1.中國海洋大學海水養殖教育部重點實驗室,山東青島266003;2.青島嶗東海珍品良種培育有限公司,山東青島266105;3.Haskin Shellfish Research Laboratory,Institute of Marine and Coastal Sciences,Rutgers University,Port Norris,New Jersey 08349 USA)
貝類多倍體[1]的誘導主要是研究貝類三倍體。三倍體較二倍體具有絕對的生長優勢,個體大且繁殖季節死亡率低。又因其性腺高度不育,在繁殖季節,仍具有豐富的糖原,口感好,很受消費者喜歡,具很高的經濟價值[2]。三倍體貝類比正常貝類個體多一套染色體,不會對自然群體造成物種污染,既提高了貝類產量,增加經濟價值,又為海產貝類育種提供了新的育種概念等,均是三倍體貝類的研究的重要原因。
以往研究從理化誘導和生物方法均獲得過貝類三倍體。就牡蠣多倍體的誘導而言,最先采用的就是化學誘導方法,Stanley等[3]最早采用不同濃度的細胞松弛素B處理美洲牡蠣(Crassostrea virginica)受精卵,得到8個月三倍體子代。之后長牡蠣(C.gigas)[4-7]、食用牡蠣(Ostrea edulis)[8]、悉尼巖牡蠣(Saccostrea commercialis)[9]和僧帽牡蠣(O.cucullata)[10]等牡蠣也成功誘導出三倍體。因理化誘導極易受外界因子如溫度、密度等的影響,三倍體誘導率無法達到100%,但生物雜交方法通過四倍體牡蠣和二倍體牡蠣雜交,可產生全三倍體牡蠣[11],前景較好[12]。
以往研究證明了雜交獲得三倍體的誘導率高,方法簡單,避免理化誘導對受精卵造成的影響,提高了孵化率和變態率[13],也初步證實了三倍體牡蠣的生長優勢[11,24]。但如何利用不同地域的二倍體群體進行大規模生產,進一步證實和提高雜交三倍體性狀,研究不多。本實驗利用中國太平洋牡蠣自交、誘導及中國二倍體太平洋牡蠣與美國四倍體太平洋牡蠣雜交,對誘導獲得的三倍體群體及雜交獲得的三倍體,進行生長發育比較來評估2種獲得三倍體的方法,為三倍體誘導方法的選擇及其今后的大規模生產奠定了基礎。
1 材料與方法
1.1 親貝的來源與促熟
中國太平洋牡蠣取自威海文登海區。美國四倍體太平洋牡蠣(雄)來自美國華盛頓州。親貝在嶗東海珍品培育有限公司育苗池網箱中暫養促熟。
1.2 實驗設計
1.2.1 實驗方案 本試驗采用3個試驗組,分別為2NC、3NC、3N H;其中,2NC二倍體對照組:中國二倍體太平洋牡蠣自交組;3NC誘導三倍體群體組:中國二倍體太平洋牡蠣(2NC)雌♀×中國二倍體太平洋牡蠣(2NC)雄♂,再經150μmol/L的6-二甲氨基嘌呤(6-DMAP)抑制受精卵第二極體的排放;3NH雜交三倍體組:中國二倍體太平洋牡蠣(2NC)雌♀×美國四倍體太平洋牡蠣(用4NA表示)雄♂。
1.2.2 受精 取性腺少量細胞,顯微鏡下鑒別雌雄,并檢查精卵的成熟狀況。雌雄牡蠣嚴格分開,以免精子污染。用解剖刀逐個刮取成熟性腺部,將卵用250目篩絹過濾后至標記好的小桶中熟化1 h,舍棄上層卵液。受精前檢查卵子是否已經受精,將獲得的精、卵按照實驗設計組合受精,迅速混合,攪拌均勻,于孵化桶中孵化。整個操作過程中,各實驗組嚴格隔離,使其不受外來精、卵的影響。
1.3 幼蟲及稚貝的培育
受精卵經24 h發育為D形幼蟲。選優后,幼蟲培育密度設為8~10個/mL,培育水溫21℃,餌料每天投喂3次,前期為金藻,后期為扁藻與小球藻(1∶1)混合投喂。每天全量換水,為防止不同實驗組幼蟲之間混雜,換水網箱單獨使用。定期調整密度,消除養殖密度對幼蟲生長的影響。
1.4 數據測量
(1)分別測量各組卵徑、卵裂率、D幼率和D形幼蟲大小。
(2)生長用殼高、殼長衡量。幼蟲階段每3 d測量1次;稚貝階段在40、70、110和140日齡測量。每次測量設3個重復,每個重復隨機測量30個個體。(3)測定浮游期各試驗組幼蟲的存活率。
(4)測定各試驗組倍性。幼蟲期取D形幼蟲以及在稚貝期取個體鰓組織,收集到1.5 m L離心管中,加入DAPI熒光染液1 m L,振蕩器震蕩0.5 min,用Partec PAⅡ流式細胞儀(FCM)進行倍性檢測。
1.5 數據處理
采用Excel作圖,SPSS 13.0統計軟件進行分析處理,不同試驗組間數據的比較采用單因素方差分析方法,差異顯著性設置為P<0.05。
2 結果
2.1 卵徑、卵裂率、D幼率、D形幼蟲大小及倍性
試驗各組的卵徑、卵裂率、D幼率、D形幼蟲大小及三倍體誘導率見表1。3個試驗組在卵徑和卵裂率方面均無顯著性差異(P>0.05),雜交三倍體的D形幼蟲規格顯著大于誘導群體組和二倍體自交組(P<0.05)。但在D幼率上看,3N H組個體顯著高于3NC群體組(P<0.05),與對照二倍體組無顯著差異(P>0.05)。在三倍體誘導率方面,雜交獲得的三倍體率為100%,明顯高于誘導群體組獲得的三倍體倍化率。
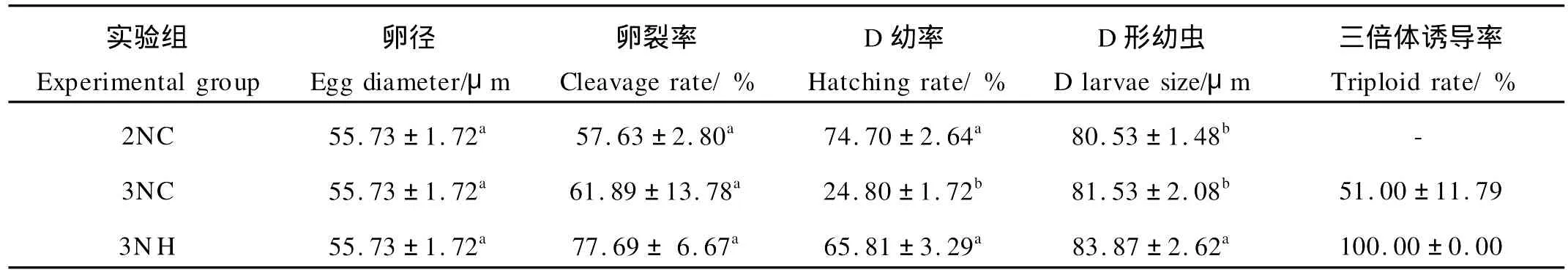
表1 誘導三倍體群體與雜交三倍體卵徑、卵裂率、D幼率、D形幼蟲大小及倍性測定的比較Table 1 Comparison of egg-diameter,cleavage rate,hatching rate,D larval size and ploidy for trip loid Pacific oyster by hybridized and induced
2.2 浮游期幼蟲存活率
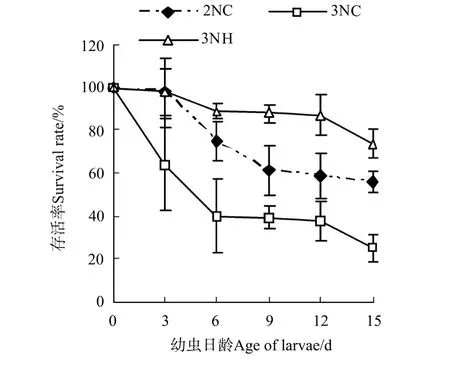
圖1 誘導三倍體群體與雜交三倍體幼蟲存活比較Fig.1 Survival rates of trip loid Pacific oysters by hybridized and induced
將D形幼蟲的存活率定義為100%,由各試驗組存活曲線(見圖1)可以看出,隨著幼蟲日齡的增長,各組幼蟲的存活率下降,且各試驗組的存活率變化不同。3日齡時,2NC和3N H組存活率彼此間無顯著差異(P>0.05),但顯著高于3NC群體(P<0.05);6~15日齡期間,各試驗組的幼蟲存活率順次為3NH>2NC>3NC群體,且各組間差異顯著(P<0.05)。
2.3 雜交三倍體與誘導三倍體群體生長比較
2.3.1 幼蟲的生長 各試驗組幼蟲的生長曲線如圖2。3組的殼高生長速度分別為2NC組(13.73±0.98)μm/d,3NC群體組(15.80±0.83)μm/d和3NH組(16.41±0.61)μm/d。9日齡以前,各試驗組幼蟲的大小彼此間無顯著差異;9~18日齡期間,3NH組殼高生長速度最快,表現出明顯的生長優勢。整個幼蟲浮游期間,從幼蟲的生長速度上看:3NH組>3NC群體組>2NC組,且3組間差異均顯著(P<0.05)。
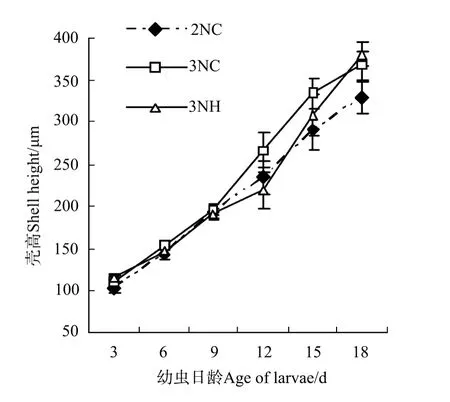
圖2 誘導三倍體群體與雜交三倍體幼蟲生長比較Fig.2 Shell height of trip loid Pacific oysters by hybridized and induced
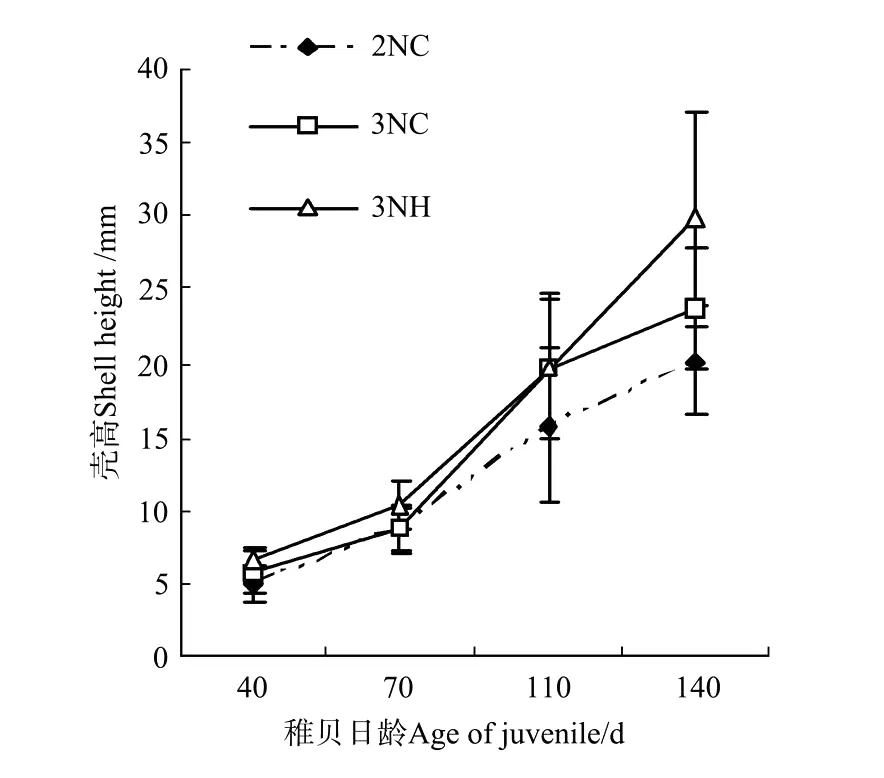
圖3 誘導三倍體群體與雜交三倍體稚貝殼高的生長比較Fig.3 Shell height of juvenile of triploid Pacific oysters by hybridized and induced

圖4 誘導三倍體群體與雜交三倍體稚貝殼長的生長比較Fig.4 Shell length of juvenile of triploid Pacific oysters by hybridized and induced
2.3.2 稚貝的生長 各試驗組稚貝期殼高和殼長的生長曲線如圖3、4。隨著培養時間的延長,3組殼長和殼高均保持相同的增長趨勢。稚貝期3組的殼高生長速度分別是2NC組(0.151±0.025)mm/d,3NC群體組(0.179±0.028)mm/d和3NH組(0.230±0.065)mm/d;殼長的生長速度分別是2NC組(0.099±0.010)mm/d,3NC群體組(0.110±0.016)mm/d和3NH組(0.132±0.031)mm/d。稚貝期各試驗組殼高、殼長的生長速度:3NH>3NC群體>2NC,且彼此間的生長速度差異顯著(P<0.05)。
2.4 稚貝期三倍體群體中二倍體和三倍體與雜交三倍體生長比較
在稚貝期(110日齡)對誘導三倍體群進行了倍性檢測,并對分離出的三倍體和二倍體以及雜交三倍體進行了生長測量,殼高和殼長的生長情況如表2。結果表明,到110日齡的稚貝期,誘導群體中的三倍體在殼高和殼長2方面稍小于雜交三倍體,但無明顯差異(P>0.05),而誘導群體中的二倍體則比二倍體對照組生長快,尤其殼高明顯大于對照組。
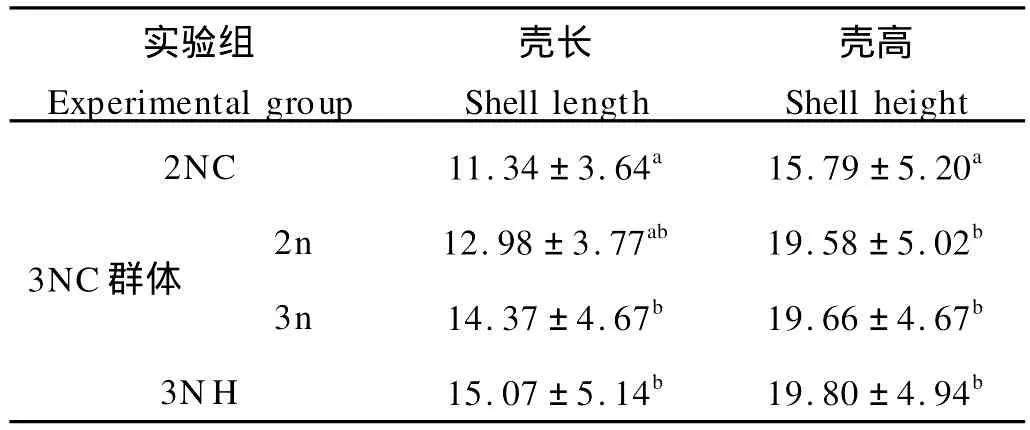
表2 稚貝期(110日齡)三倍體群體中二倍體和三倍體與雜交三倍體生長性狀比較Table 2 Comparison of shell length and shell height of juvenile of dip loid and trip loid Pacific oyster by hybridized and induced(Day 110)/mm
3 討論
本實驗所選6-二甲氨基嘌呤誘導三倍體的方法是因為6-二甲氨基嘌呤作為誘導劑,誘導率高,毒性小,操作方便,價格便宜[14],現已被廣泛采用。
雜交、誘導2種方法獲得的三倍體卵裂率高,生長速度快,D形幼蟲大小均大于二倍體對照組,但D形幼蟲率很低。尤其是誘導獲得的三倍體群體組D形幼蟲率低,由于6-DMAP的影響,使得牡蠣幼蟲出現畸形,該組死亡率在前期迅速下降。但雜交三倍體組存活率很穩定。應與化學誘導劑對受精卵產生的影響有關,這與田傳遠等[15]結果一致。12日齡以后,各組均出現幼蟲大量沉底現象,闕華勇等[13]雜交多倍體實驗中也有類似現象出現。與對照組相比,雜交三倍體和誘導三倍體群體從浮游幼蟲階段就表現出生長優勢,此結果與王昭萍等[16]結論一致。從三倍體率來看,雜交獲得100%的三倍體,較誘導群體組誘導率高且穩定。
通過對誘導三倍體群體和雜交三倍體的對比,從卵裂率、D幼率、存活率、生長發育和倍性等方面均體現了雜交三倍體具有優勢。在稚貝期將誘導三倍體群體中的三倍體與雜交三倍體進行生長比較,雜交三倍體稍大于前者,但是差異不顯著,這與王昭萍等[27]結果相吻合。分析本實驗誘導三倍體群體與雜交三倍體之間的生長差異原因可能有2個:一是在誘導三倍體群體中二倍體含量過高,降低了三倍體的實際生長水平;二是雜交三倍體是中美牡蠣雜交,即為兩種不同地理群體雜交,表現出了雜種優勢。而誘導群體中的二倍體則比二倍體對照組生長快,可能是誘導群體中的二倍體經過了藥物處理所致。
雜交獲得三倍體即為誘導多倍體方法中的生物方法,該方法簡單易操作,三倍體率高達100%,避免了理化處理對胚胎發育和環境的影響。這同陶抵輝、于瑞海等[17]觀點一致。此結果同時也驗證了雜交三倍體在生產應用上的可行性,同先前的于瑞海等[18]研究結果一致。
雜交獲得三倍體的方法具有很多優點,但雜交方法中的四倍體親本的獲得是該方法的關鍵,也是瓶頸。雖有很多種養殖牡蠣[8,19-21]進行四倍體誘導,但目前只有太平洋牡蠣、美洲牡蠣和近江牡蠣建立了穩定的四倍體群系[22-24]。故四倍體的研究仍是多倍體方面需要繼續探索的一個方向。
闕華勇等進行了四倍體太平洋牡蠣(G)和二倍體近江牡蠣(A)的雜交試驗[25]。種間雜交子代GGA組幼蟲個體較GA組大,眼點幼蟲形成也在GA組之前。同時證實了雜交三倍體兼具了兩親本的特性,如適鹽范圍變寬了。而本實驗兩親本太平洋牡蠣分別來自于中國威海和美國華盛頓,為種內雜交三倍體子代。種內雜交,是目前雜種優勢利用的主要途徑[26]。本實驗并未對子代的生理等進行試驗,不能確定是否兼具不同地理群體兩親本的優勢,是本實驗需要繼續研究的一個方向。
牡蠣作為水產經濟貝類之一,本實驗通過對雜交三倍體和誘導三倍體群體的生長狀況具體比較,驗證雜交的方法獲得的全三倍體,提高了牡蠣生產的產量,存活率也很高,適合在生產上廣泛應用,是加速貝類養殖業的發展的一個可行的重要方向。
致謝:感謝青島亞特蘭生物科技有限公司提供四倍體牡蠣親貝;青島嶗東海珍品良種培育有限公司郭希瑞、馬強、孫軍委等對本試驗的幫助。
[1] 王昭萍,王如才,張筱蘭.貝類多倍體育種研究及應用現狀[J].齊魯漁業,1999,16(5):32-33.
[2] 王昭萍,王如才,于瑞海.多倍體貝類的生物學特征[J].青島海洋大學學報,1998,28(3):399-404.
[3] Stanley J G,Allen S K,Hidu H.Ployploidy induced in the American oyster Crassostrea virginica withcytochalasin B[J].Aquaculture,1981,23:1-10.
[4] A llen S K,Dow ning SL,Chaiton JC,et al.Chemically and p ressure induced triploidy in the Pacific oyster,Crassostrea gigas[J].Aquaculture,1986,57:359-360.
[5] Yamamoto S,Sugawara Y,Nomura T,et al.Induced triploidy in Pacific oyster,Crassosyrea gigas,and performance of trip loid larvae[J].Tohoku Journal of Agricultural Research,1988,39(1):47-59.
[6] Yamamoto S,Sugawara Y,Nomura T.Chemical and thermal control of triploid production in Pacific oysters and mussels,with regard to controlling meiotic maturation[M].∥Hoshim,Yamashita O.Advances in invertebrate rep roduction.Amsterdam:Elsevier,1990:455-460.
[7] Desrosiers R R,Gerard A,Peignon JM,et al.A novelmethod to produce triploids in bivalvemolluscs by the use of 6-dimethylaminopurine[J].Journal of Experimental Marine Biology and Ecology,1993,170(1):29-43.
[8] Gendreau S,Grizel H.Induced trip loidy and tetrap loidy in the European flat oyster,Ostrea edulis L[J].Aquaculture,1990,90:229-238.
[9] Nell J A,Cox E,Smith IR,et al.Studies on triploid oysters in Australia.Ⅰ.The farming potential of triploid Sydney rock oysters Saccostrea commercialis(Iredale and Roughley)[J].Aquaculture,1995,137:355-358.
[10] 曾志南,陳木,林琪,等.僧帽牡蠣三倍體的研究[J].海洋通報,1994,13(6):34-40.
[11] Guo X,DeBrosse G A,A llen S K.All-triploid Pacific oysters(Crassostrea gigas Thunberg)produced by mating tetraploids and diploids[J].Aquaculture,1996,142:149-161.
[12] 王昭萍,李慷均,于瑞海.貝類四倍體育種研究進展[J].中國海洋大學學報:自然科學版,2004,34(2):195-202.
[13] 闕華勇,張國范,劉曉,等.雄性四倍體與雌性二倍體雜交培育全三倍體長牡蠣(Crassostrea gigas)的研究[J].海洋與湖沼,2003,34(6):656-662.
[14] 王冬群,李太武.6-DMAP在貝類多倍體誘導中的應用[J].河北漁業,2003(1):3-4.
[15] 田傳遠,梁英,王如才.6-DMAP誘導太平洋牡蠣三倍體-誘導因素對孵化率和D幼畸形率的影響[J].中國水產科學,2000,7(1):110-112.
[16] 王昭萍,姜波,孔令峰,等.利用四倍體與二倍體雜交規模化培育全三倍體太平洋牡蠣苗種[J].中國海洋大學學報:自然科學版,2004,34(5):742-746.
[17] 陶抵輝,劉明月,肖君澤.生物多倍體誘導方法研究進展[J].生命科學研究,2007,11(4):6-13.
[18] 于瑞海,王昭萍,施坤濤.單體全三倍體太平洋牡蠣育苗技術的研究[J].海洋湖沼通報,2008(3):115-120.
[19] Stephens L B,Downing SL.Inhibition of first polar body formation in Crassostrea gigas produces tetraploids,notmeioticI triploids[J].J Shellfish Res,1988,7(3):550-551.
[20] Supan J E,Allen S K,Wilson C A.Tetraploid eastern oysters:An arduous effort[J].J Shellfish Res,2000,19(1):655.
[21] 容壽柏,李一民,劉紹瓊,等.用冷熱休克誘導四倍體近江牡蠣[J].湛江水產學院學報,1992,12(2):18-21.
[22] Eudeline B,Allen S K,Guo X.Optimization of tetraploid induction in the Pacific oysters,Crassostrea gigas,using the first polar body as a natural indicator[J].Aquaculture,2000,187:73-84.
[23] Guo X,Allen S K.Viable tetraploid Pacific oyster(Crassostrea gigas Thunberg)produced by inhibiting polar body Iin eggs fromtriploids[J].Mol Mar Biol Biotech,1994,3(1):42-50.
[24] Guo X,Wang Y,Xu Z,et al.Chromosome set manipulation in shellfish[M].∥Burnell G,Allam G.New technologies in aquaculture improving production efficiency,guality and environmental management.Cambridge:Waodhead Publishing Ltd,2009,165-194.
[25] Que Huayong,Allen S K.Hybridization of tetraploid and diploid Crassostrea gigas(Thunberg)with diploid C.ariakensis(Fujita)[J].Journal of Shellfish Research,2002,21(1):137-143.
[26] 李云玲,孫振興,張明青.貝類雜交育種技術及其應用[J].齊魯漁業,2005,22(12):22-24.
[27] Wang Zhaoping,Guo Ximing,Allen S K,et al.Heterozygosity and body size in triploid Pacific oysters,Crassostrea gigas Thunberg,produced from meiosis II inhibition and tetraploids[J].Aquaculture,2002,204:337-348.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
