遲鈍愛德華氏菌菌蛻系統的構建
2011-01-12 06:57:28牟為管玲玉王啟要劉琴張元興
微生物學雜志 2011年3期
關鍵詞:系統
牟為,管玲玉,王啟要,劉琴,張元興
(華東理工大學生物反應器國家重點實驗室,上海200237)
遲鈍愛德華氏菌(Edwardsiella tarda,E.tarda)是一種革蘭陰性的兼性胞內寄生菌。該菌不僅是多種淡、海水養殖魚類的致病菌,同時也能感染兩棲類及哺乳類等。遲鈍愛德華氏菌是目前在水產養殖中有極大危害的病原菌,目前已在許多種海洋魚類養殖中引發病害造成了巨大損失[1-2]。它的感染能引起魚類的出血性敗血病,使其出現腹部積水腫脹、體表出血、腸內出現黏液等癥狀,稱為愛德華菌病(Edwardsiellosis)[3-5]。用化學試劑或者抗生素控制此類細菌性魚病會引起微生物的抗生素耐藥性,有害物質殘留和環境污染等影響,因此,以預防為目的的魚用疫苗已成為防止漁業病害最為有效的方法。近年來,魚用疫苗的發展相當迅速,疫苗的種類日益增多,其中多數為滅活疫苗(Killed vaccine),也有少數減毒活疫苗(Live attenuated vaccine)和亞單位疫苗(Subunit vaccine)等[4,6-7]。魚類免疫的方法主要包括腹腔或肌肉內注射,浸泡和口服。這3種方法在實用性和成本方面均有各自的優缺點。注射免疫能有效利用抗原,獲得較強的免疫效果,但易使魚受傷,且費時費力,對較小的魚不適用。浸泡免疫省時方便,但疫苗進入魚體的效率和吸收效果受到多種因素限制,并且利用細菌減毒活疫苗進行浸泡免疫時,存在菌株返毒和與野生毒株發生重組導致新毒株出現的可能,有一定的安全隱患[6,8]。菌蛻(Bacterial Ghost,BG)是革蘭陰性菌被噬菌體PhiX174裂解基因E裂解后形成的。lysisE基因并不導致宿主細胞完全裂解,而是引起細胞膜出現孔道,使得細胞內容物經由裂解孔道排出,形成不含核酸、核糖體及其他組分的幾乎完好無損的空殼細胞[9-11]。傳統的菌蛻系統是在溫度誘導系統PI/pR-cI857轉錄控制下形成E基因的可控表達,當溫度低于30℃時,熱敏感阻遏蛋白cI857的表達使基因E能被穩定抑制,細菌正常生長;溫度高于30℃時,因阻遏蛋白cI857的熱失活而使基因E開始表達,細菌裂解形成菌蛻,在42℃時細菌的裂解達到最佳狀態。鐵是一種重要而必需的過渡金屬元素,幾乎所有的生物體都需要游離鐵離子來進行呼吸作用和氧運輸等,但是過量的鐵也會造成機體損傷[12-13]。因此,微生物在進化過程中生成了多種調控系統以嚴格控制鐵元素的攝取和釋放。多數細菌鐵攝取系統在鐵豐富的環境中并不表達,主要原因是其編碼基因的-35至-10區通常存在著一個Fur蛋白結合盒(Fur box)。Fur是一種鐵依賴性的全局調控因子(Global regulator),存在于各種細菌鐵攝取系統,在大腸埃希菌中超過90種基因的表達受Fur調控。其中35種與鐵攝取相關的基因在富鐵環境中受Fur嚴格抑制,保證鐵轉運系統受到鐵濃度的嚴格調控[14-16]。本研究利用受鐵濃度調控的啟動子PyncE[15,17]來構建新型的遲鈍愛德華氏菌菌蛻系統,研究并比較其裂解效率、制備條件、菌體形態與傳統菌蛻的區別,為進一步研究高效的菌蛻疫苗奠定基礎。
1 材料與方法
1.1 材料
1.1.1 菌株與質粒E.coli Top10F'為本實驗室保藏,含有大質粒pEIB202的E.tarda EIB202菌株為本實驗室前期分離得到,所有菌株于-80℃保存于含20%甘油的LB培養基中。商業化原核表達質粒pUC18、pBV220為本實驗室前期購買保藏(Invitrogen),菌蛻質粒pPR-cI-Elysis為Lubitz W教授惠贈。
1.1.2 主要試劑本實驗所需的常規化學試劑主要采購自中國醫藥集團上海化學試劑公司。限制性內切酶SalⅠ、HindⅢ以及PCR所用DNA聚合酶(Pyrobest DNA polymerase)、DNA連接試劑盒(DNA ligation kit 2.0)均購自寶生物(TaKa-Ra),普通Taq酶預混反應液(2×Taq Marster Mix)及質粒抽提、膠回收試劑盒、DNA產物純化試劑盒均購自天根生化(Tiangen,北京)。
1.1.3 主要儀器電泳儀、瓊脂糖電泳槽、iCycler PCR反應儀以及酶標儀購自BIORAD公司;FR-200紫外與可見分析裝置購自上海復日生物工程公司;掃描電子顯微鏡(Hitachi S-2400)購自Olympos公司。
1.2 方法
1.2.1 質粒構建常規DNA技術操作參照文獻[18]進行,所用引物見表1。裂解基因lysisE通過PCR從PhiX174基因簇上得到,PR-cI857調控系統從質粒上獲得,鐵調控啟動子PyncE及其上游序列來源于E.coli TOP10基因組。用overlap PCR的方法分別將兩類啟動子與lysisE基因融合,擴增出的融合片段PR-E和PyncE-E用SalⅠ、HindⅢ酶切到表達質粒pUC18中,構建重組質粒,分別命名為LPR-E/pUC18和PyncE-E/pUC18。
1.2.2 遲鈍愛德華氏菌菌蛻制備及檢測將上文構建的2種質粒LPR-E/pUC18、PyncE-E/pUC18電轉化至E.tarda EIB202感受態細胞,得到的陽性菌株接種于含有Ampr抗性的LB液體培養基中,28℃200 r/min搖床過夜培養;次日,2重組
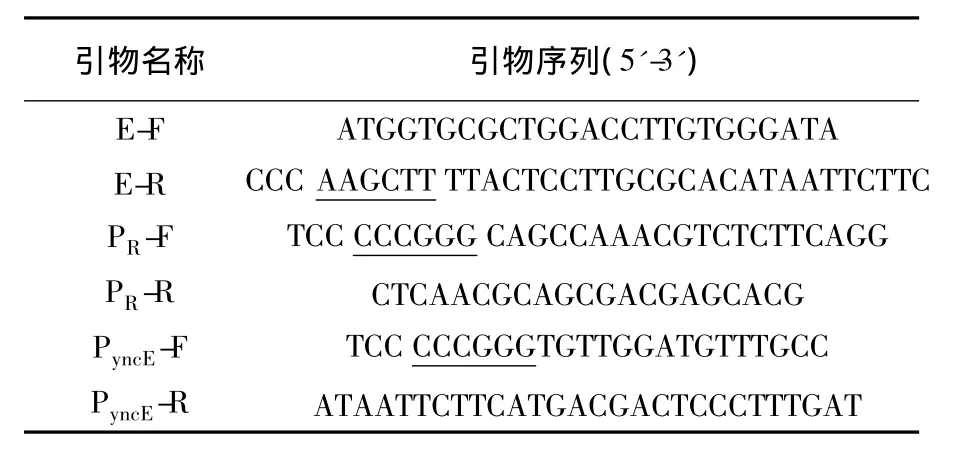
表1 所用引物列表Table 1 Primers used in this study
1.2.3 掃描電鏡觀察將誘導后生成的E.tarda菌蛻用2.5%戊二醛4℃固定2 h后用PBS洗滌3次,經乙醇逐級脫水和醋酸異戊脂置換等步驟處理后,將細胞進行冷凍干燥,用含銀離子的包埋劑處理后即可進行掃描電鏡(Hitachi S-2400)觀察。
2 結果與分析

圖1 融合片段PR-E和PyncE-E的overlap PCR擴增產物驗證Fig.1 Overlap PCR amplification of PR-E and PyncE-E
分別以菌蛻質粒pPR-cI-Elysis和E.coli Top10F'染色體DNA為模板,利用PCR擴增出裂解基因lysisE、溫度誘導啟動子PR和鐵調控啟動子PyncE(圖1),膠回收純化后的產物進行重疊延伸(overlap)-PCR,得到融合片段LPR-E和PyncEE。將融合片段經酶切SalⅠ、HindⅢ后連接到pUC18中,構建重組質粒LPR-E/pUC18和PyncEE/pUC18。將連接液電轉化入E.tarda EIB202,并對其轉化子進行菌落PCR檢測,經1%瓊脂糖電泳后,分別出現1 543 bp和874 bp的條帶,其結果跟預期的一致,測序后發現無突變堿基,說明重組質粒LPR-E/pUC18、PyncE-E/pUC18已經成功轉化進入E.tarda EIB202中。
2.2 菌蛻系統生成效率檢測
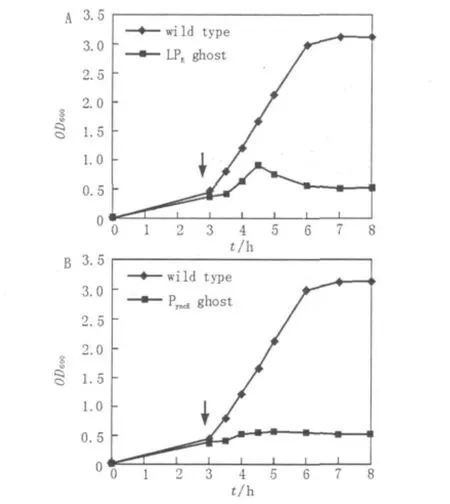
圖2 菌蛻系統對E.tarda的溶菌過程Fig.2 Growth curves of E.tarda harboring plasmidPyncE-E/pUC18 or PR-E/pUC18
將重組菌株E.tarda EIB202(LPR-E/pUC18)在28℃培養至對數中期,將菌株在42菌株分別轉接種于30 mL Ampr抗性的LB液體中,28℃200 r/min搖床培養至OD600為0.3~0.5時,將含有PR-E/pUC18的菌株移至42℃搖床中培養,在含有PyncE-E/pUC18的菌株加入鐵離子螯合劑二聯吡啶后繼續28℃培養。自此,每30 min取樣,測OD600的吸光度值,直到誘導后培養物的OD600趨于平穩為止。
2.1 菌蛻系統構建
℃環境下繼續培養,誘導lysis E表達。如圖2A所示,細菌在繼續生長一段時間后開始裂解,在溫度升高后1.5 h,可檢測到OD600開始下降,OD600的檢測數值從0.92持續下降到0.48,至誘導后4 h開始趨于平穩。同樣,將重組菌株E.tarda EIB202(PyncE-E/pUC18)在28℃培養至對數生長中期,在培養基中加入鐵螯合劑二連吡啶誘導lysisE表達。結果如圖2B顯示,誘導后菌株即停止生長。這說明E蛋白能夠在2套ghost系統中正常表達,并對宿主菌E.tarda EIB202產生致死或抑制其生長的作用。隨后的菌落涂板實驗也證實實際活菌數量與OD600所顯示情況相符。
2.3 掃描電鏡觀察
掃描電鏡觀察如圖3所示,大多數傳統菌蛻E.tarda EIB202(LPR-E/pUC18)在誘導后生成空泡狀結構,細菌的細胞兩極可見直徑之間的溶菌孔道和部分胞內物質,說明E蛋白能夠融合在遲鈍愛德華氏菌的形成特異性跨膜通道,并使得細菌內外滲透壓發生改變,從而使內含物流出。而新型菌蛻E.tarda EIB202(PyncE-E/pUC18)誘導后的細菌有一些并未全部裂解或形成孔道結構,僅有部分形成了致死的菌蛻細胞,但由于胞內壓力和部分胞內物質被溶解等原因,鐵誘導表達系統介導的菌蛻使細菌表面形態有明顯的皺縮和拉長等現象。

圖3 掃描電鏡觀察菌蛻形成情況Fig.3 Scanning electron micrograph of E.tarda ghost
3 討論
菌蛻作為新型非活性疫苗,能夠保持與活細菌相同的完好形態結構,可誘導機體產生體液免疫和細胞免疫,包括黏膜免疫,并且不具有活菌疫苗的致病作用[9]。同時,由于菌蛻保留有細菌的菌毛、纖毛,具有生物親和性,能夠進行特異的細胞和組織定位,可用作核酸和藥物的親和性載體。利用菌蛻系統作為抗原載體接收大量的外源抗原或重組蛋白,將這些抗原蛋白通過特定的膜錨定結構插入細胞膜,或填充于胞周間隙和細胞空腔中,可以獲得重組菌蛻多價苗和多聯苗[19-20]。之前的研究表明,細菌菌蛻特別適合作為黏膜免疫的疫苗運送系統,通過口服途徑也可激發體液和細胞免疫應答[21-23],但由于傳統的菌蛻生成調控系統是依靠溫度調控,因此,該系統要求在30℃以下的條件進行細菌的培養,而許多病原菌在這一溫度培養時很難達到正常的生長。因而,開發新型的可調控系統將有利于菌蛻疫苗的生產。
本研究利用細菌在生長過程中嚴格受鐵調控這一特性,挑選出一個鐵調控型啟動子PyncE,并以此為基礎構建了新型的鐵調控菌蛻系統。因為在細菌普通培養基LB中含有豐富的游離鐵離子,正常培養時受Fur蛋白調控的PyncE在豐富鐵環境下不表達裂解lysisE基因,而當培養基中加入鐵螯合劑后,因為鐵離子缺乏,PyncE開啟轉錄表達,使得細菌裂解。
與傳統菌蛻受到誘導后大量死亡不同,鐵調控菌蛻能夠讓宿主菌在誘導后保持低密度生長。
電鏡結果表明,鐵調控菌蛻除生成孔洞外,還有一些因內部裂解壓力而發生拉伸,細菌分化不完全等現象,這可能與PyncE與LPR啟動效率不同有關,說明在不同的啟動子調控下,菌蛻會出現不同的形式。目前,利用細菌菌蛻作為抗原呈遞載體已成為疫苗開發的熱點,本實驗中構建新型的調控系統,使得細菌在維持少量生存的情況下形成菌蛻,對以后遲鈍愛德華氏菌菌蛻疫苗及載體疫苗的開發和設計奠定了基礎。
[1] 單曉楓,高云航,李影.魚用疫苗研究進展[J].中國獸藥雜志,2005,39(11):19-22.
[2] Sakazaki R,Murata Y.The new group of the Enterobacteriaceae,Jap[J].J.Bacteriol,1962,17(13):616-617.
[3] Brenner DJ,Fanning GR,Steigerwalt AG.Polynucleotide sequence relatedness in Edwardsiella tarda[J].J.Syst.Bacteriol,1974,24(21):186-190.
[4] Castro N,Toranzo AE,Barja JL,et al.Characterization of Edwardsiella tarda strains isolated from turbot,Psetta maxima[J].J.Fish.Dis,2006,29(14):541-547.
[5] Chowdhury R,Wakabayshi H.Survival of four major bacterial fish pathogens in different types of experimental water[J].J.Microbiol,1990,7(16):47-54.
[6] Gudding R,Lillehaug A,Evensen O.Recent developments in fish vaccinology[J].Vet.Immunol.Immunopathol,1999,72(11):203-212.
[7] Smith GP.Filamentous fusion phage:novel expression vectors that display cloned antigens on the virion surface[J].Science,1985,228(21):1315-1317.
[8] Sang YL,Jong HC,Xu ZH.Microbial cell-surface display[J].Trends.Biotechnol,2003,21(31):45-52.
[9] Detmer A,Glenting J.Live bacteria vaccines:a review and identification of potential hazards[J].Microbial.Cell.Factories,2006,5(12):23-26.
[10] Witte A,Wanner G,Sulzner M,et al.Dynamics of PhiX174 protein E-mediated lysis of Escherichia coli[J].Arch.Microbiol,1992,157(31):381-338.
[11] Serruto D,Rappuoli R.Post-genomic vaccine development[J].FEBS.Letters,2006,580(21):2985-2992.
[12] Dill BD,Raulston JE.Examination of an inducible expression system for limiting iron availability during Chlamydia trachomatis infection[J].Microbes.Infect,2007,9(41):947-953.
[13] Ratledge C,Dover LG.Iron metabolism in pathogenic bacteria[J].Annu.Rev.Microbiol,2000,54(17):881-941.
[14] Wooldridge KG,Williams PH,Wanner G.Iron uptake mechanisms of pathogenic bacteria FEMS[J].Microbiol.Rev,1993,12(13):325-348.
[15] McHugh JP,Rodriguez F,Abdul H,et al.Global iron-dependent gene regulation in Escherichia coli.A new mechanism for iron homeostasis[J].J.Biol.Chem,2003,278(41):29478-29486.
[16] Stojiljkovic I,Baumler J,and Hantke K.Fur regulon in gramnegative bacteria.Identification and characterization of new iron-regulated Escherichia coli genes by a fur titration assay[J].J.Mol.Biol,1994,236(31):531-545.
[17] Baba-Dikwa A,Thompson D,Spencer NJ,et al.Overproduction,purification and preliminary X-ray diffraction analysis of YncE,an iron-regulated Sec-dependent periplasmic protein from Escherichia coli[J].Acta.Crystallogr.Sect.F.Struct.Biol.Cryst.Commun,2008,64(11):966-969.
[18] Sambrook J,Russell DW.黃培堂等譯.分子克隆實驗指南(第3版)[M].北京:科學出版社,2002.
[19] Eko FO,Witte A,Huter V,et al.New strategies for combination vaccines based on the extended recombinant bacterial ghost systern[J].Vaccine,1999,17(41):1643-1649.
[20] Schon P,Sehrot G,Lubitz W,et al.Two-stage model for integration of the lysis protein E of PhiX174 into the cell envelope of Escherichia coli[J].FEMS.Microbiol..Rev,1995,17(11):207-212.
[21] Huter V,Hensel A,Brand E,et al.Improved protection against lung colonization by Actinobacillus pleuropneumoniae ghosts:characterization of a genetically inactivated vaccine[J].J.Biotechnol,2000,83(21):161-172.
[22] Jalava K,Hensel A,Szostak MP,et al.Bacterial ghosts as vaccine candidates for veterinary applications[J].J.Control.Release,2002,85(17):17-25.
[23] Mayr UB,Hailer C,Haidinger W,et al.Bacterial ghosts as an oral vaccine:a single dose of Escherichia coli O157∶H7 bacterial ghosts protects mice against lethal challenge[J].Infect.Immun,2005,73(51):4810-4817.
猜你喜歡
工業設計(2022年8期)2022-09-09 07:43:20
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
裝備制造技術(2019年12期)2019-12-25 03:06:46
制造技術與機床(2019年10期)2019-10-26 02:47:06
中國洗滌用品工業(2019年4期)2019-05-11 09:27:34
鐵道通信信號(2018年5期)2018-06-28 03:06:24
家庭影院技術(2017年9期)2017-09-26 03:41:45
知識經濟·中國直銷(2017年5期)2017-06-15 20:28:19
通信電源技術(2016年6期)2016-04-20 06:21:32
