模擬輪伐期長短對杉木人工林氮動態的影響
2011-07-24 01:49:46辛贊紅接程月魏曉華BLANCOJuan周國模
浙江農林大學學報 2011年6期
關鍵詞:模型
辛贊紅, 江 洪,2, 接程月, 魏曉華, BLANCO Juan, 周國模
(1. 浙江農林大學 國際空間生態與生態系統生態研究中心, 浙江 臨安311300; 2. 南京大學 國際地球系統科學研究所, 江蘇 南京210093; 3. 不列顛哥倫比亞大學 地球與環境科學系, 不列顛哥倫比亞 基隆那V1V1V7)
森林生態系統的養分循環是生態系統的重要功能過程之一, 直接影響著森林的生產力[1-2]。 作為生命必需元素之一的氮是影響森林生態系統生產力的重要限制因素[2]。 研究一定立地條件下森林生態系統中氮元素的分配、 積累和循環, 可以進一步認識森林生態系統的功能[3]。 這對于改進森林經營策略, 促進林木速生、 豐產、 優質也有重要意義。 杉木Cunninghamia lanceolata 是中國南方最重要的用材樹種, 生長快, 材質好, 單產高, 已有1 000 多年的栽培歷史, 在中國南方林業生產中占舉足輕重的地位[4]。 近年來由于杉木連栽造成生產力日趨下降, 已嚴重影響了杉木人工林的持續經營[5-8]。 長期定位試驗是研究森林連栽或輪伐對地力影響的重要方法, 但是, 其需時長, 不確定性大, 操作起來有很大的難度。 現階段, 利用模型進行相關的模擬, 已是一個重要的發展趨勢[9-10]。 森林生態系統生物地球化學過程模擬模型是了解森林生態系統生物組分與其無機環境之間相互作用的重要工具[9]。 目前已經開發出了許多種生態系統過程模擬模型用以森林生態系統的研究[10-13], FORECAST 模型也是其中重要的一種[14]。 本研究通過運用FORECAST 模型模擬杉木人工林在不同輪伐期的生長過程, 揭示其氮循環的特點, 從而為深入了解杉木人工林在連栽條件下的養分循環機理和杉木人工林的合理經營管理提供基礎數據。
1 研究方法
1.1 FORECAST 及其氮循環的簡單示意圖
FORECAST 模型是一個以林分群體特征及林地養分循環為基礎的森林生態系統管理模型[14]。 FORECAST 模型關注的是土壤有機物和營養庫不斷變化的層面上樹木的生長狀況和林材的收獲產量。 運用該模型進行模擬可以得到土壤有機物和營養庫中各種營養元素循環變化的情況。 FORECAST 模型使用的模擬方法是將生物產量表方法和過程模擬方法相結合的混合模擬方法。 模型的驅動函數是葉氮同化率(FNE)。 FORECAST 模型氮循環的簡單示意圖1 所示。
1.2 杉木FORECAST 模型的校準和應用
FORECAST 模型的應用有4 個步驟: ①數據的收集、 輸入和驗證; ②建立生態系統模擬運行的初始條件; ③定義森林管理方式或者干擾方式; ④模擬各種管理方式并分析模型輸出結果。 前2 個步驟即為模型的參數校準。 模型的校準數據是由描述不同養分條件林地上林木(地上部分和地下部分)和小型植被按照林分生長的時間序列而逐漸累積的生物量的數據組成。 林木生物量和林分自疏率通常由樹高、 胸徑和林分密度結合具有樹種特異性的組分生物量異速生長方程得到。 在模型的養分模擬部分, 需要不同養分在各種生物量組分中的含量的數據。 模型同時還需要不同葉量所產生的遮光度的數據和樹葉對不同光照強度響應的數據。 模型校準所需的這些數據主要來自于文獻, 包括福建、 湖南等主要杉木中心產區的研究報告和江西、 廣西等氣候相似區域的杉木林的研究文獻。
第2 階段的校準需要在“設置” 模式下運行模型來建立立地初始條件。 在這個階段, 模型運行時需關閉養分反饋, 從而使表示立地條件的植被、 凋落物和土壤有機質得以積累, 這反映出歷史的積累模式。
杉木人工林的模型參數在借鑒Bi 等[15]的基礎上進行了一些調整, 數據主要來源于田大倫等編著的《杉木林生態系統》以及盛煒彤等編著的《杉木人工林長期生產力保持機制研究》。 模型中的降水營養輸入、 坡面滲流、 礦質土壤陽離子和陰離子交換能力、 腐殖質質量、 凋落物分解速率等數據來源于廖利平等[16-17]、黃志群等[18]、 丁應祥等[19]、 楊玉盛等[20]的研究結果。 樹木生物量、 死亡率和林分密度、 樹高以及林冠高度、 活組織的營養濃度的數據以楊玉盛等[21]、 肖文發等[22]、 林開敏等[23]、 劉茜等[24]、 周后盛等[25]發表的數據為基礎。
1.4 模擬方案

圖1 FORECAST 模型氮循環的簡單示意圖Figure 1 Diagram of nitrogen cycling in FORECAST model
模型的校準數據來自中國可觀測的杉木人工林范圍內的所有立地, 立地按照質量情況分為較差、 中等、 較好, 并被量化為17, 21 和27(從差到好, 基于優勢木50 a 的生長高度)來代表模型推演林木生長的相關指數。 FORECAST 模型中數量化的立地質量可以是但并不是必須基于立地指數即某一指數年齡優勢木的平均高度[26-27]。 模擬的年限為200 a; 3 種不同的輪伐期為10, 25 和50 a; 模擬的收獲方式為莖干采伐法(每次輪伐期末除去90%的樹干生物量, 殘留部分均遺留在原地)。 除采伐收獲外沒有其他干擾。
2 模擬結果
2.1 FORECAST 模型模擬結果的檢驗
模型驗證有很多方法。 本研究采用模型模擬結果與實驗測量數據對比的方法來進行驗證。 由于比較不同輪伐期(不同代數)養分氮循環的研究很少, 所以我們獲得的相關文獻數據非常有限。 根據劉愛琴等[1]的結果, 可以得到立地指數為16 的2 代11 年生和31 年生杉木人工林氮吸收量的數據。 由模擬結果可知, 3 種不同立地20 a 的優勢木高分別為: 較差立地12.1 m, 中等立地15.3 m, 較好立地19.1 m。 對照文獻地位指數, 可以選用中等立地11 a 輪伐期末和31 a 輪伐期末的氮吸收量數據與文獻數據進行比較。 模擬結果與實測數據的對比如表1。 將模擬結果與實測數據作回歸分析, 可得方程y = 1.895 6x-76.728, r2= 0.969 3**。 由此可見: 模型模擬第1 代和第2 代杉木人工林氮的吸收量有較高的可信度。
2.2 土壤有效氮的模擬結果
由圖2 可見: 3 種不同立地條件下,輪伐期長短對土壤有效氮含量有顯著的影響。 在較差立地上, 10 a 輪伐期的前5個輪伐期土壤有效氮含量較大, 下降也較快; 從第6 個輪伐期開始有小幅下降, 然后逐漸過渡到平穩。 25 a 輪伐期的第1 個輪伐期, 其土壤有效氮含量小于10 a 輪伐期第1 個輪伐期的值, 但是25 a 輪伐期的下降幅度較小, 并且在第2 個輪伐期之后就逐漸平穩。 50 a輪伐期的土壤有效氮含量高于25 a 輪伐期的土壤有效氮含量, 并且高于第2 個輪伐期之后的10 a 輪伐期的值; 50 a 輪伐期的土壤有效氮含量在各個輪伐期之間的變化不大。 中等立地和較好立地有相似的規律, 只是相比差等立地25 a 輪伐期的值要低于10 a 輪伐期的值。
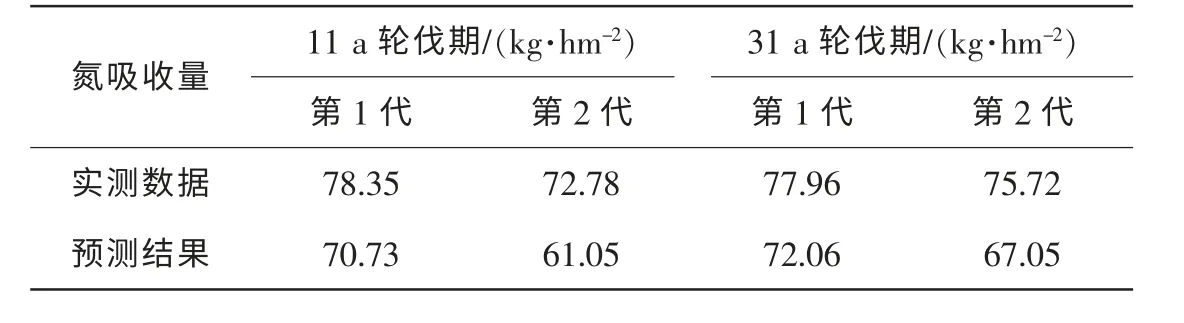
表1 氮吸收量預測數據與實測數據對比Table 1 Comparison between predicted data and observed data
2.3 氮的吸收量和歸還量
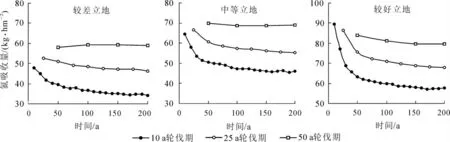
圖3 不同輪伐期氮的年均吸收量Figure 3 Uptake N in different rotations of Chinese fir plantation
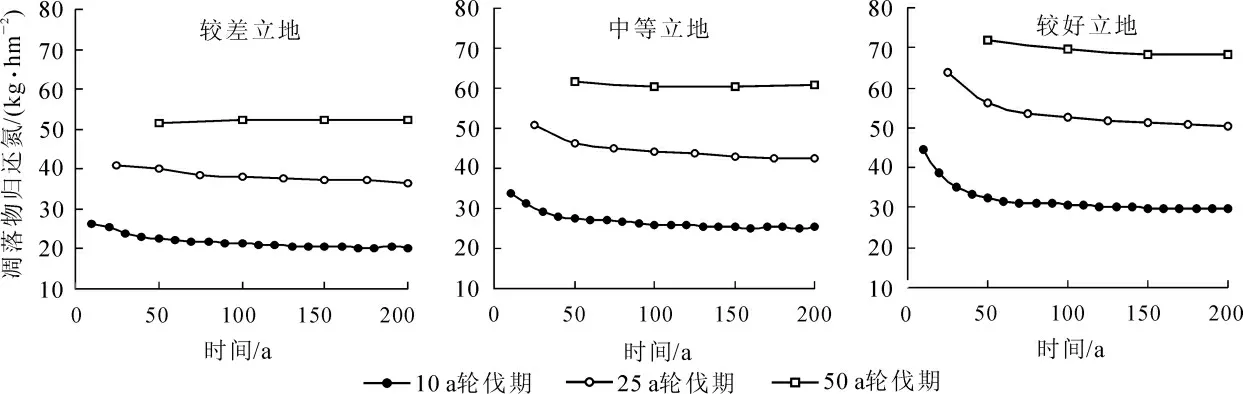
圖4 不同輪伐期凋落物形式氮的年歸還量Figure 4 Average quantity of N returned by litterfall at different rotations
由圖3 和圖4 可見: 3 種不同立地條件下, 氮的年吸收量和凋落物形式氮的年歸還量, 總體趨勢都是隨著輪伐期的增長而增加, 隨著輪伐代數的增加而減少。 10 a 輪伐期的吸收量和歸還量在前4 個輪伐期有明顯的下降, 而且下降幅度相較25 a 輪伐期要大; 50 a 輪伐期的吸收量和歸還量, 在各輪伐期之間變化不大, 趨勢相對比較平穩。 由于模擬采用的是莖干采伐的收獲方式, 枝葉和根均遺留在原地, 所以大量的氮以凋落物的形式歸還給土壤。 以中等立地為例, 不同輪伐期氮的循環系數[28](即氮的年歸還量比氮的年吸收量)分別為: 0.544, 0.766, 0.847 和0.879, 可見隨著輪伐期增長氮的循環系數增大。
2.4 土壤中氮的年淋溶損失
由圖5 可知: 杉木人工林的土壤氮淋溶損失量總體是隨著立地條件的變好而增大。 3 種不同立地條件下, 10 a 輪伐期的年平均土壤氮淋溶損失量較大, 并且在前3 個輪伐期表現尤為明顯, 3 個輪伐期之后逐漸平穩; 25 a 輪伐期的年均土壤氮淋溶損失量較小, 只在第1 個輪伐期和第2 個輪伐期之間變化比較大; 50 a 輪伐期的年均土壤氮淋溶損失量較小, 輪伐期之間的變化也不大。 總體趨勢上, 較差立地和中等立地, 土壤氮淋溶損失量都是隨著輪伐期的增長而減小; 較好立地50 a 輪伐期的土壤氮淋溶損失量要大于25 a 輪伐期的土壤氮淋溶損失量, 但是小于10 a 輪伐期的淋溶量。
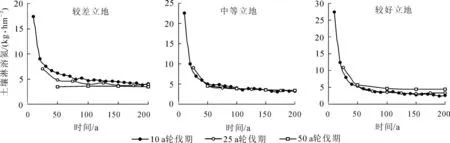
圖5 不同輪伐期土壤氮的淋溶損失量Figure 5 Chinese fir plantation soil N leaching loss at different rotations
2.5 葉片含氮量的動態
葉片含氮量是FORECAST 模型一個重要的輸出參數。 通過這個參數, 我們可以看出伴隨著林分生長, 氮元素在葉片中積累的量。 圖6 顯示了3 種立地條件下, 不同輪伐期葉片含氮量的變化趨勢。 由圖6 可知: 隨著輪伐期的增長積累在葉片中的氮逐漸增加。 10 a 輪伐期的葉片含氮量隨著立地條件變好與25 a 輪伐期的差距逐漸減小, 這可能是因為隨著立地條件的變好, 林木迅速生長, 使得10 a 輪伐期的密度優勢逐漸顯現了出來, 從而造成了葉片含氮量在總量上的增加。
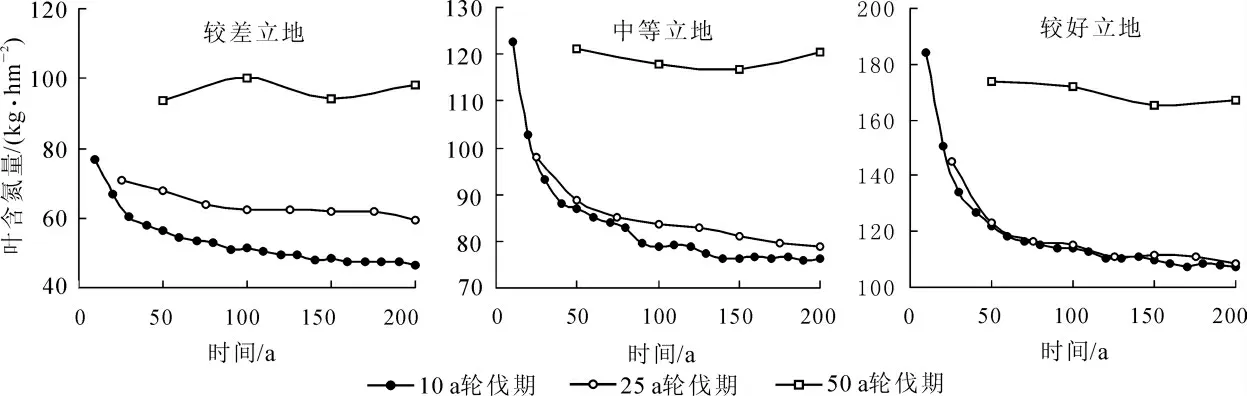
圖6 杉木人工林不同輪伐期葉片氮含量的動態Figure 6 Chinese fir plantation foliage N dynamics at different rotations
3 分析與討論
3.1 FORECAST 模型模擬的精度
FORECAST 模型通過大量森林生態系統的模擬, 已得到了較好的檢驗[29-34]。 對于杉木森林生態系統,Bi 等[15]作了實測數據與模擬結果的測試, 證明模擬結果精度較高。 本模擬的結果也證明, FORECAST 模型模擬杉木林生態系統氮循環的關鍵變量, 具有較高的模擬精度, 可以廣泛應用在有關的模擬實踐中。
3.2 不同輪伐期杉木人工林生態系統氮元素的變化趨勢
輪伐期的長短會影響杉木人工林氮元素的循環和利用。 彭長輝等[10]、 江洪等[11]運用Century 4.0 模型對加拿大北方森林和中國北方森林所做的研究顯示, 在采伐強度一定的情況下, 土壤有效氮含量在長輪伐期下最大, 并且隨著輪伐期的縮短而減小。 極短的輪伐期會造成北方森林減產達65%。 通過FORECAST 模型的模擬結果我們可以看出: 在采伐方式一定的情況下, 短輪伐期(10 a)土壤有效氮含量下降,氮吸收量降低, 長輪伐期(50 a)土壤有效氮和氮的吸收都保持較大的量并且輪伐期之間的變化較平穩。 隨著輪伐期的增長, 凋落物形式氮的年歸還量也逐漸增大。 以中等立地為例, 10, 25, 50 a 輪伐期土壤有效氮200 a 的平均值分別為96.24, 86.87 和96.07 kg·hm-2; 25 a 輪伐期相對10 a 輪伐期減少了9.7%;50 a 輪伐期與10 a 輪伐期基本持平, 相對25 a 輪伐期增加10.6%。 吸收氮的平均值分別為49.22, 58.44和69.07 kg·hm-2; 25 a 輪伐期相對10 a 年輪伐期增加了18.7%; 50 a 輪伐期相對10 a 輪伐期增加了40.3%, 相對25 a 輪伐期增加18.2%。 凋落物形式歸還氮的平均值分別為26.75, 44.76 和60.74 kg·hm-2;25 a 輪伐期相對10 a 年輪伐期增加了67.3%; 50 a 輪伐期相對10 a 輪伐期增加了127.1%, 相對25 a 輪伐期增加35.7%。 從模擬結果看: 10 a 短輪伐期下土壤氮的淋溶損失總量, 較差立地為1 167.57 kg·hm-2,中等立地為1 075.27 kg·hm-2, 較 好立 地為1 085.70 kg·hm-2; 25 a 輪伐期分別為895.05, 893.60 和924.28 kg·hm-2; 50 a 輪伐期分別為707.33, 765.98 和959.05 kg·hm-2。 10 a 短輪伐期土壤氮的淋溶損失總量均高于同等立地條件下25 a 輪伐期和50 a 輪伐期的總量。 從以上模擬結果可以看出: 輪伐期的長短對杉木人工林生態系統氮元素的循環和利用具有重要影響, 過短的輪伐期會導致土壤有效氮含量下降, 氮吸收量降低, 淋溶損失增加, 從而破壞土壤肥力, 造成林木生產力的下降。
3.3 連栽代數對杉木人工林氮元素循環和利用的影響
從模擬結果可以看出: 相同輪伐期不同代數杉木林的氮循環和利用存在著顯著差異。 隨著代數的增加, 土壤有效氮含量、 年吸收量和年歸還量均呈遞減趨勢, 而且這種趨勢隨著輪伐周期的變短而愈加明顯。 以中等立地前4 代(即前4 個輪伐期)為例, 10 a 輪伐期的第2 代、 第3 代和第4 代土壤有效氮較第1 代分別減少了17.2%, 23.6%和27.2%; 年吸收量分別減少了10.1%, 16.9%和20.4%; 年歸還量分別減少了8.1%, 14.1%和17.9%。 25 a 輪伐期的第2 代、 第3 代和第4 代土壤有效氮相較第1 代分別減少了10.6%, 13.1%和14.5%; 年吸收量分別減少了9.2%, 12.0%和13.9%; 年歸還量分別減少了8.9%,11.8%和13.4%。 可見, 隨著杉木連栽代數的增加, 土壤有效氮減少, 土壤供肥和保肥的能力下降, 吸收氮和凋落物歸還氮的量也在減少, 從而可能導致林分產量的降低。 本研究的模擬結果中, 第1 代、 第2 代和第3 代土壤有效氮的變化幅度較大, 這與盛煒彤[6]、 楊玉盛等[22]研究者的結論相似。
3.4 展望
在這次模擬工作中, 只是研究不同立地條件下輪伐期長短對杉木人工林生態系統氮動態的影響, 而沒有考慮煉山、 整地和疏伐等育林干擾。 我們將進一步完善模型, 加入煉山、 整地、 疏伐等育林措施,使模型模擬更貼近現實的生產管理, 為杉木人工林的合理經營提供更為準確的數據。
[1] 劉愛琴, 范少輝, 林開敏, 等. 不同栽植代數杉木林養分循環的比較研究[J]. 植物營養與肥料學報, 2005, 11 (2):273 - 278.LIU Aiqin, FAN Shaohui, LIN Kaimin, et al. Comparison on nutrient cycling in different generation plantations of Chinese-fir [J]. Plant Nutr Fert Sci, 2005, 11 (2): 273 - 278.
[2] 諶小勇, 潘維儔. 杉木人工林生態系統中氮素的動態特征[J]. 生態學報, 1989, 9 (3): 201 - 206.SHEN Xiaoyong, PAN Weichou.Nitrogen dynamics of Chinese-fir plantation ecosystem [J].Acta Ecol Sin, 1989, 9 (3):201 - 206.
[3] 沈國舫, 董世仁, 聶道平. 油松人工林養分循環的研究營養元素的含量及分布[J]. 北京林學院學報, 1985, 7 (4):1- 14.SHEN Guofang, DONG Shiren, NIE Daoping. Study on nutrient cycling in Pinus tabulaeformis plantation ecosystem[J]. J Beijing For Coll, 1985, 7 (4): 1 - 14.
[4] 范少輝, 盛煒彤, 馬祥慶, 等. 多代連栽對不同發育階段杉木人工林生產力的影響[J]. 林業科學研究, 2003, 16(5): 560 - 567.FAN Shaohui, SHENG Weitong, MA Xiangqing, et al. Effects of successive-generation planting on Chinese-fir plantation productivity at different development stages [J]. For Res, 2003, 16 (5): 560 - 567.
[5] 徐化成. 關于人工林的地力下降問題[J]. 世界林業研究, 1992, 5 (1): 66 - 73.XU Huacheng. On the plantation soil fertility declination [J]. World For Res, 1992, 5 (1): 66 - 73.
[6] 盛煒彤. 人工林地力衰退研究[M]. 北京: 中國科學技術出版社, 1992.
[7] 俞新妥. 杉木林地持續利用問題的研究和看法[J]. 世界林業研究, 1993, 6 (2): 80 - 86.YU Xintuo. Study and suggestions on the sustainable utilization of Chinese fir land [J]. World For Res, 1993, 6 (2):80 - 86.
[8] 馬祥慶. 杉木人工林連栽生產力下降研究進展[J]. 福建林學院學報, 2001, 21 (4): 380 - 384.MA Xiangqing.Advance in researches on productivity decline of repeating Chinese fir forests [J].J Fujian Coll For, 2001,21 (4): 380 - 384.
[9] KROS H, WARFVINGE P. Evaluation of model behaviour with respect to the biogeochemistry at the Solling spruce site [J]. Ecol Model, 1995, 83: 255 - 262.
[10] PENG Changhui, JIANG Hong, APPS M J, et al. Effects of harvesting regimes on carbon and nitrogen dynamics of boreal forests in central Canada: a process model simulation [J]. Ecol Model, 2002, 155: 177 - 189.
[11] JIANG Hong, APPS M J, PENG Changhui, et al. Modeling the influence of harvesting on Chinese boreal forest carbon dynamics [J]. For Ecol Manage, 2002, 169: 65 - 68.
[12] FERRIER R C, WRIGHT R F, COSBY B J, et al. Application of the MAGIC model to the Norway spruce stand at Solling, Germany [J]. Ecol Model, 1995, 83: 77 - 84.
[13] VERBURG P S J, JOHNSON D W. A spreadsheet-based biogeochemical model to simulate nutrient cycling processes in forest ecosystems [J]. Ecol Model, 2001, 141, 185 - 200.
[14] 接程月, 辛贊紅, 信曉穎, 等. FORECAST 模型的原理、 方法和應用[J]. 浙江林學院學報, 2009, 26 (6): 909- 915.JIE Chengyue, XIN Zanhong, XIN Xiaoying, et al.Principle, method and application of FORECAST model [J].J Zhejiang For Coll, 2009, 26 (6): 909-915.
[15] BI J, BlANCO J A, SEELY B, et al. Yield decline in Chinese-fir plantations: a simulation investigation with implications for model complexity [J]. Can J For Res, 2007, 37: 1615 - 1630.
[16] 廖利平, 楊躍軍, 汪思龍, 等. 杉木Cunninghamia lanceolata, 火力楠Michelia macclurei 純林及其混交林細根分布、 分解與養分歸還[J]. 生態學報, 1999, 19 (3): 246 - 250.LIAO Liping, YANG Yuejun, WANG Silong, et al. Distribution, decomposition and nutrient return of the fine root in pure Cunninghamia lanceolata, Michelia macclurei and the mixed plantations [J]. Acta Ecol Sin, 1999, 19 (3):246 - 250.
[17] 廖利平, 馬越強, 汪思龍, 等. 杉木與主要闊葉造林樹種葉凋落物的混合分解[J]. 植物生態學報, 2000, 24 (1):27 - 33.LIAO Liping, MA Yueqiang, WANG Silong, et al. Decomposition of leaf litter of Chinese fir in mixture with major associated broad-leaved plantation species [J]. Acta Phytoecol Sin, 2000, 24 (1): 27 - 33.
[18] 黃志群, 廖利平, 汪思龍, 等. 杉木根樁和周圍土壤酚含量的變化及其化感效應[J]. 應用生態學報, 2000, 11(2): 190 - 192.HUANG Zhiqun, LIAO Liping, WANG Silong, et al. Dynamics of phenolics content of Chinese fir stump-roots and the rhizosphere soil and its allelopathy [J]. Chin J Appl Ecol, 2000, 11 (2): 190 - 192.
[19] DING Yingxiang, TIAN Ye. A testing simulation with FOORTOON on long-term productivity of Chinese fir plantations [J]. For Study China, 1999, 1 (2): 34 - 38.
[20] 楊玉盛, 陳光水, 黃寶龍. 杉木多世代連栽的土壤水分和養分變化[J]. 南京林業大學學報, 2000, 24 (2): 25 -28.YANG Yusheng, CHEN Guangshui, HUANG Baolong. Variation in the soil water and nutrients between different rotation stands of Chinese fir [J]. J Nanjing For Univ, 2000, 24 (2): 25 - 28.
[21] 楊玉盛, 邱仁輝, 何宗明, 等. 不同栽杉代數29 年生杉木林凈生產力及營養元素生物循環的研究 [J]. 林業科學, 1998, 34 (6): 3 - 11.YANG Yusheng, QIU Renhui, HE Zongming, et al. Study on the stand net productivity and biological cycle of nutrients in the 29-year-old plantations of Chinese fir on different rotations [J]. Sci Silv Sin, 1998, 34 (6): 3 - 11.
[22] 肖文發, 聶道平, 張家誠. 我國杉木林生物量與能量利用率的研究[J]. 林業科學研究, 1999, 12 (3): 253 -258.XIAO Wenfa, NIE Daoping, ZHANG Jiacheng. Study on biomass and energy use efficiency of the stands of Cunninghamia lanceolata in China [J]. For Res, 1999, 12 (3): 253 - 258.
[23] 林開敏, 俞新妥, 何智英, 等. 不同密度杉木林分生物量結構與土壤肥力差異研究[J]. 林業科學, 1996, 32(5):385 - 392.LIN Kaimin, YU Xintuo, HE Zhiying, et al. Study on the difference of biomass structure and soil fertility in Chinese fir stands of different densities [J]. Sci Silv Sin, 1996, 32 (5): 385 - 392.
[24] 劉茜. 杉木種子營養化學成分的研究[J]. 林業科學, 1998, 34 (2): 92 - 95.LIU Qian. A study on the chemical composition of Chinese fir seeds [J]. Sci Silv Sin, 1998, 34 (2): 92 - 95.
[25] 周后盛. 杉木多代連栽地營造混交林生產力的初步研究[J]. 福建林學院學報, 1999, 19 (2): 165 - 169.ZHOU Housheng.Study on productivity of mixed forest planted on site of continuously planted Chinese fir [J].J Fujian Coll For, 1999, 19 (2): 165 - 169.
[26] KIMMINS J P, MAILLY D, SEELY B. Modeling forest ecosystem net primary production: the hybrid simulation approach used in FORECAST [J]. Ecol Model, 1999, 122: 195 - 224.
[27] KIMMINS J P. Scientific Foundations for the Simulation of Ecosystem Function and Management in FORCYTE-l1[R]. Vancouver: University of British Columbia. Faculty of Forestry. Department of Forest Sciences, 1993: 3 - 7.
[28] 方晰, 田大倫, 蔡寶玉, 等. 第2 代杉木人工林氮、 磷、 鉀的吸收、 積累和遷移[J]. 中南林學院學報, 2002, 22(2): 1 - 6.FANG Xi, TIAN Dalun, CAI Baoyu, et al. Absorption, accumulation and transportation of N, P, K elements in the second-generation Chinese-fir plantation [J]. J Central South For Univ, 2002, 22 (2): 1 - 6.
[29] MORRIS D M, KIMMINS J P, DUCKERT D R. The use of soil organic matter as a criterion of the sustainability of forest management alternatives: a modeling approach using FORECAST [J]. For Ecol Manage, 1997, 94: 61 - 78.
[30] WEI Xiaohua, LIU W, WATERHOUSE J, et al. Simulations on impacts of different management strategies on longterm site productivity in lodgepole pine forests of the central interior of British Columbia [J].For Ecol Manage, 2000,133: 217 - 229.
[31] WEI Xiaohua, KIMMINS J P, ZHOU Guoyi. Disturbance and the sustainability of long-term site productivity in lodgepole pine forests in the central interior of British Columbia: an ecosystem modeling approach [J]. Ecol Model,2003, 164: 239 - 256.
[32] SEELY B, WELHAM C, KIMMINS J P. Carbon sequestration in boreal forest ecosystem: results from the ecosystem simulation model, FORECAST [J]. For Ecol Manage, 2002, 169: 123 - 135.
[33] WELHAM C, SEELY B, KIMMINS J P. The utility of the two-pass harvesting system: an analysis using the ecosystem simulation model FORECAST [J]. Can J For Res, 2002, 32: 1071 - 1079.
[34] REICH P B, GRIGAL D F, ABER J D, et al. Nitrogen mineralization and productivity in 50 hardwood and conifer stands on diverse soils [J]. Ecology, 1997, 78: 335 - 347.
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19

- 浙江農林大學學報的其它文章
- 模擬邯鄲地區自然群落的植物景觀設計
- 浙江農林大學獲國家自然科學基金資助項目32項