彌漫性腦創(chuàng)傷后Bcl-2家族與線粒體途徑細(xì)胞凋亡的相關(guān)性分析
2011-08-20 10:40:20王金林尹立國
中國實驗診斷學(xué) 2011年12期
王金林,尹立國,李 軍
(1.唐山市工人醫(yī)院,河北唐山 063000;2.遵化市人民醫(yī)院;3.玉田縣第二醫(yī)院)
隨著凋亡機(jī)制的深入研究,線粒體在細(xì)胞凋亡過程中的中心地位逐漸得到確認(rèn),線粒體通透性轉(zhuǎn)換孔(mitochondrial permeability transition pore,MPTP)由于其在線粒體和細(xì)胞存活和死亡中的重要作用而備受廣大學(xué)者關(guān)注。近年來的研究顯示,bcl-2家族與線粒體膜通透性轉(zhuǎn)換孔關(guān)系密切。Bcl-2和Bax是Bcl-2家族中最主要的兩個成員,分別具有抑制和促進(jìn)細(xì)胞凋亡的作用,本研究以Marmarou方法建立大鼠彌漫性腦創(chuàng)傷模型,試圖通過相關(guān)性分析進(jìn)一步說明其在線粒體途徑細(xì)胞凋亡中的作用。
1 材料與方法
1.1 腦創(chuàng)傷模型的制備與分組雄性成年SD大鼠90只,體重300-350 g,隨機(jī)分為實驗對照組、顱腦創(chuàng)傷組 。每組設(shè)傷后 1 h 、3 h、6 h、24 h 、48 h、72 h 6個時相點(diǎn)。對照組每個時相點(diǎn)3只大鼠,創(chuàng)傷組每時相點(diǎn)6只大鼠。各組未達(dá)到規(guī)定時相點(diǎn)死亡的大鼠從實驗中剔除。動物置乙醚麻醉缸中,以乙醚吸入方式深度麻醉動物,麻醉后的大鼠俯臥于落體致傷海綿墊上,70%酒精消毒顱頂部皮膚,沿正中線矢狀切開頭皮,剝離骨膜,顯露人字縫與冠狀縫,將一不銹鋼墊固定在大鼠冠狀縫與人字縫之間將大鼠頭部固定于打擊模型架下的頭部固定裝置中,使重450 g,直徑18 mm的銅柱沿垂直玻璃管于1.2 m高度自由落下,撞擊大鼠顱骨頂部的鋼墊,造成大鼠中度彌漫性顱腦損傷。撞擊后即刻移開大鼠,以免銅柱反彈造成頭部二次打擊傷,給予顱腦損傷后大鼠,頭皮切口常規(guī)消毒、縫合。對照組僅給予相同的麻醉處理及沿中線矢狀切開頭皮、剝離骨膜、在冠狀縫與人字縫之間粘鋼墊,置致傷海綿墊上,但不行自由落體致傷,而后常規(guī)消毒、縫合切口.各組未到規(guī)定時相死亡的大鼠予以剔除。
1.2 細(xì)胞色素c、Bax和Bcl-2檢測標(biāo)本甲醛固定,石蠟切片,按武漢博士德提供檢測試劑盒說明書方法進(jìn)行檢測。
1.3 原位細(xì)胞凋亡檢測(TUNEL法)按Roche公司提供TUNEL檢測試劑盒進(jìn)行操作。
1.4 陽性細(xì)胞免疫反應(yīng)強(qiáng)弱采用Motic Med 6.0計算機(jī)真彩色圖象分析系統(tǒng)以平均光密度進(jìn)行半定量分析;實驗數(shù)據(jù)用SPSS11.5 for windows軟件進(jìn)行統(tǒng)計學(xué)處理。
2 結(jié)果
2.1 傷后1 h至24 hCytc表達(dá)與TUNEL陽性細(xì)胞數(shù)密切相關(guān)(r=0.958,P<0.05)(圖1)。
2.2 傷后1 h至24 h Bax和Bcl-2二者的比例與Cytc免疫反應(yīng)相關(guān)(r=0.996,P<0.01)(圖2)。
2.3 傷后1 h至72 h Bax和Bcl-2二者的比例與TUNEL陽性細(xì)胞數(shù)密切相關(guān)(r=0.925,P<0.01)(圖3)。
2.4 但Cytc表達(dá)峰值(24 h)早于Bax和Bcl-2二者的比例高峰值(48 h)(表1)。
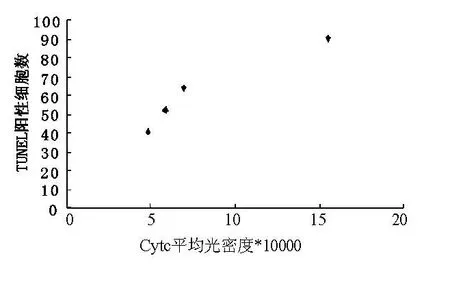
圖1 Cytc表達(dá)與TUNEL陽性細(xì)胞數(shù)的相關(guān)性
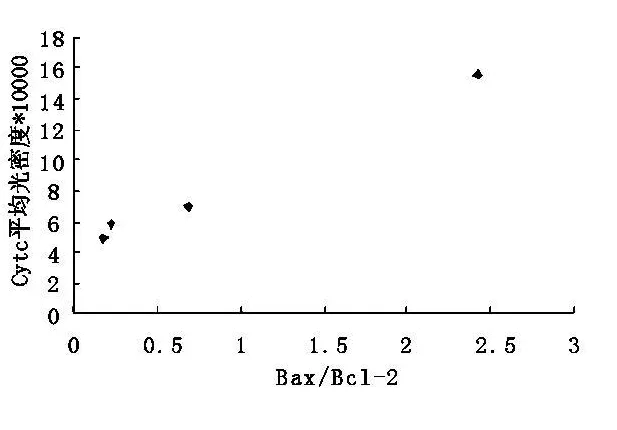
圖2 Cytc表達(dá)與bax/bcl-2的相關(guān)性
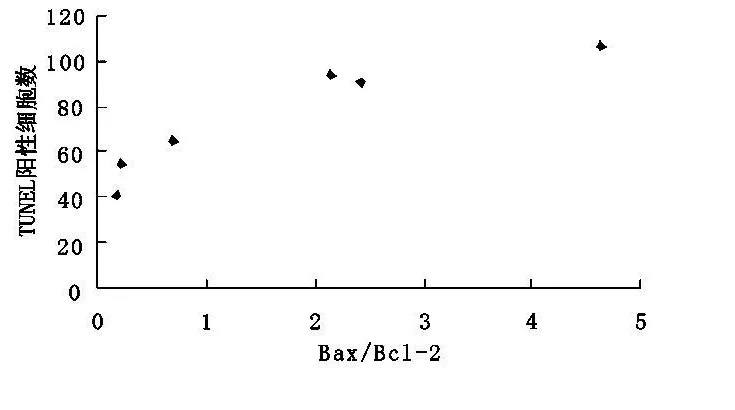
圖3 bax/bcl-2與TUNEL陽性細(xì)胞數(shù)的相關(guān)性

表1 腦創(chuàng)傷后 cyt-c、bax、bcl-2與細(xì)胞凋亡數(shù)的動態(tài)變化
3 討論
Bcl-2屬于一類新的癌基因家族,主要包括抗凋亡蛋白(bcl-2、bcl-xl、bcl-wl等)和促凋亡蛋白(bax、bak、bolt等)。近年來研究顯示Bcl-2家族蛋白對PTP的開放和關(guān)閉起著關(guān)鍵的調(diào)節(jié)作用,促凋亡蛋白bax等可以通過與ANT或VDAC的結(jié)合介導(dǎo)PTP開放,而抗凋亡蛋白bcl-2、bcl-xl等可以通過與bax競爭性的與ANT結(jié)合,或直接阻止bax與ANT、VDAC的結(jié)合來發(fā)揮其抗調(diào)亡作用[1,2]。在純化的線粒體中重組Bax、Bak具有開放PT孔的作用。可能是因為其具有與通道蛋白相似的三維空間結(jié)構(gòu),能形成足夠大的通道來轉(zhuǎn)運(yùn)ctyc,平時Bax與Bcl-2或Bcl-xl形成異二聚體,使PT處于關(guān)閉狀態(tài)。當(dāng)細(xì)胞發(fā)生調(diào)亡時,與Bcl-2,Bcl-xl相連的Bax含量增加,形成Bax同型二聚體,PT通道開放,使ctyc釋放[3]。目前大量的研究顯示細(xì)胞內(nèi)還存在著以bax為核心的不依賴與PTP復(fù)合體的凋亡調(diào)控機(jī)制。體外實驗中發(fā)現(xiàn)bax能在人工構(gòu)建的脂質(zhì)體膜上形成通過大分子物質(zhì)的孔道而不需要PTP復(fù)合體的參與,但必須依賴于bid蛋白和心肌磷脂的存在[4],提示bax可能具有不依賴PTP復(fù)合體來介導(dǎo)細(xì)胞色素c等物質(zhì)釋放的能力。而在生理狀況下bax以單體的形式于胞漿中,研究表明只有在受到凋亡信號刺激后,bax轉(zhuǎn)位到線粒體膜并暴露其BH3結(jié)構(gòu)域后,才能介導(dǎo)細(xì)胞色素c釋放等下游事件[5]與此同時,相關(guān)研究還發(fā)現(xiàn)bax單體無法在線粒體膜上形成孔道和釋放細(xì)胞色素c,只有在形成低聚物后才能介導(dǎo)細(xì)胞色素 c釋放。進(jìn)一步的研究顯示體外培養(yǎng)的HeLa細(xì)胞在外界凋亡誘導(dǎo)因子的作用下細(xì)胞內(nèi)的bax形成了兩種結(jié)合于線粒體膜的高分子量低聚復(fù)合物,而此低聚復(fù)合物中并不含有構(gòu)成PTP復(fù)合體的主要成分VDAC和ANT。還有學(xué)者發(fā)現(xiàn),發(fā)生膜轉(zhuǎn)位的bax在線粒體膜上形成了四聚體到十聚體組成的低聚復(fù)合物,同樣此低聚復(fù)合物中并不含有構(gòu)成PTP復(fù)合體的主要成分VDAC和ANT顯著,而bcl-2則可以顯著抑制此低聚復(fù)合物的形成[6]。另有研究顯示bax在凋亡信號介導(dǎo)下發(fā)生膜轉(zhuǎn)位后,形成數(shù)千個bax分子簇,bcl-xl超表達(dá)沒有阻止bax膜轉(zhuǎn)位發(fā)生,但可以抑制分子簇的形成和凋亡的發(fā)生。有此可以看出bax對細(xì)胞色素c釋放起關(guān)鍵性作用,在凋亡信號刺激下bax從胞漿轉(zhuǎn)位到線粒體膜上從而啟動了線粒體介導(dǎo)的調(diào)亡途徑。同時轉(zhuǎn)位到線粒體膜上bax和bak低聚化或者分子簇形成很可能決定凋亡是否發(fā)生。這其中,bax很可能決定凋亡發(fā)生與否的必要因素,而bak則是其調(diào)控因素[7]。抗凋亡類bcl-2蛋白可以通過阻止促凋亡蛋白在線粒體膜上形成高分子低聚物或是在線粒體膜附近形成分子簇發(fā)揮其抗調(diào)亡作用。當(dāng)然上述研究大部分限于體外實驗,我們Marmarou方法建立彌漫性腦創(chuàng)傷模型對大鼠體內(nèi)cytc、Bcl-2、Bax蛋白進(jìn)行動態(tài)觀察和相關(guān)分析,結(jié)果發(fā)現(xiàn)傷后1 h至24 h Bax和Bcl-2二者的比例與Cytc免疫反應(yīng)相關(guān)(r=0.996,P<0.01)說明bcl-2家族與cytc釋放關(guān)系密切,細(xì)胞色素c從線粒體釋放到胞漿后,在dATP/ATP存在下,首先與凋亡激活因子1(apoptosis protease activeing factor,Apaf-1)結(jié)合,形成多聚復(fù)合體[8,9]。后者充分聚合胞質(zhì)中的procaspase-9,并導(dǎo)致其自動活化[10]成caspase-9,然后啟動級聯(lián)反應(yīng),繼續(xù)活化下游的Caspase,如Caspase-3和Caspase-7,整個過程為一正反饋,活化的Caspases能對其底物特異性的切割,DNA片段化,細(xì)胞發(fā)生凋亡,并出現(xiàn)凋亡形態(tài)學(xué)改變[11,12,13]。凋亡誘導(dǎo)因子(AIF)從線粒體釋放后直接進(jìn)入胞核內(nèi),獨(dú)立的將DNA裂解引起染色體和DNA的大規(guī)模片斷化,導(dǎo)致核凋亡[14,15]。我們的相關(guān)結(jié)果也顯示cytc陽性表達(dá)與TUNEL陽性細(xì)胞數(shù)密切相關(guān),Bax和Bcl-2二者的比例與TUNEL陽性細(xì)胞數(shù)密切相關(guān),結(jié)合前人成果可以確認(rèn)腦創(chuàng)傷cytc釋放、bcl-2家族比例變化引發(fā)了細(xì)胞凋亡。同時我們也對時相變化進(jìn)行分析,結(jié)果發(fā)現(xiàn)但Cytc表達(dá)峰值(24 h)早于Bax和Bcl-2二者的比例高峰值(48 h),說明另有其它因素作用于線粒體膜通透性轉(zhuǎn)換孔引起cytc釋放。
[1]Zomora M,Granell M,Mampel T,et al.Adenine nucleotide translocase 3(ANT3)overexpression inducws apoptosis incultured cells[J].FEBS Lett,2004,563(1-3):155.
[2]Belzacq AS,Helena LAV,Guido K,ed al.The adenine nucleotide translocator in apoptosis[J].Biochimic,2002,84(2-3):167.
[3]Degli Esposti M,Dive C.Mitochondrial Membrane permeabilisation by Bax/Bak[J].Biochemical&Biophysical Communications,2003,304(3):455.
[4]Kuwana T.Mackey MR.Perkins G,et al.Bid,Bax,and lipids cooperate to form supramolecular openings in the outer mitochondrial membrane[J].Cell,2002,111(3):331.
[5]Jia L.Patwari Y.Srinivasula SM,et al.Bax translocation is crucial for the sensitivity of leukaemic cells to etoposide-induced apoptosis[J].Oncogene,2001,20(35):4817.
[6]Mikhailov V,Mikhailov M,Donna J et al.Bcl-2 prevent Bax Oligomerization in the Mitochondrial Outer Membrane[J].J Biol Chem,2001,276(21):18361.
[7]Mikhailov V,Mikhailov M,Degenhardt et al.Association of Bax and Bak Homo-oligomers in Mitochondria[J].J Biol Chem,2003,278(7):5367.
[8]Hu Y,Benedict MA,Ding L,et al.Role of cytochrome c and dATP/ATP hydrolysis in Apaf-1-mediated caspase-9 activation and apoptosis[J].EMBO J,1999,18(13):3586.
[9]Yoshida H.Apaf-1 isrequired for mitichondrial pathways of apoptosis and brain development[J].Cell,2002,94:739.
[10]Hatai T,Matsuzawa A,Inoshita S,etal.Execution of apoptosis signal-regulating kinase-1(ASK1)-induced apoptosis by the mitochondrial-dependent caspase activation[J].J Biol Chem,2000,275(34):26576.
[11]Li Jia,Srinivasa M.Srinivasula,Feng-Ting Liu,et al.Apaf-1 protein deficiency confers resistance to ctyochrome c-dependent apoptosis in human leukemia cells[J].Blood,2001,98:414.
[12]Kawane K,Fukuyama,Yoshida H,et al.Impaired thymic development in mouse embryos deficient in apoptotic DNA degradation[J].Nat Immunol,2003,4(2):138.
[13]Pawlowski J,Kraft AS.Bax-induced apoptotic cell death[J].Proc Natl A-cad Sci USA,2000,97(2):529.
[14]Patterson SD,SD,Spahr CS,Daugas E,et al.Mass spectrometric identification of proteins released frommitochondria undergoing pemeability transition[J].Cell Death Differ,2000,7(2):137.
[15]Susin SA,Daugas E,Ravagnan L,et al.Two distinct pathways leading to nuclear apoptosis[J].J Exp Med,2000,192(4):571.
