藏北高寒草地植物群落物種多樣性沿海拔梯度的分布格局
2011-08-20 08:01:36段敏杰高清竹郭亞奇萬運帆李玉娥
草業科學 2011年10期
段敏杰,高清竹,郭亞奇,萬運帆,李玉娥,
干珠扎布1,2,旦久羅布3,韋蘭亭3,西饒卓瑪3
(1.中國農業科學院農業環境與可持續發展研究所,北京100081;2.農業部農業環境與氣候變化重點實驗室,北京100081;3.西藏自治區那曲地區草原站,西藏 那曲852100)
物種多樣性是全球變化研究的重要內容之一,也是當前現代生態學和環境學研究的熱點問題之一。各種各樣的生物資源是地球上人類賴以生存的物質基礎,不論是發展中國家還是發達國家都面臨著物種多樣性的保護和持續利用的問題[1]。在很大程度上,草地生態系統的穩定性和生產力的維持依賴于草地植物群落的物種多樣性[2-3]。可以說,物種多樣性是衡量一個地區生物資源豐富程度的重要客觀指標[4-5]。物種多樣性沿海拔梯度的變化規律是生物多樣性研究的重要議題,海拔梯度的變化綜合了溫度、濕度和光照等多種環境因子,因此,在一定程度上能夠顯著地影響植物種群的空間分布格局[6-7]。眾多研究表明,物種多樣性隨著海拔梯度的變化有多種形式,其中普遍認為二者呈負相關關系,即隨著海拔高度的增加物種多樣性降低[8-9];另一種形式是“中間高度膨脹”,即隨著海拔高度的增加,物種多樣性呈先增加、后減少的趨勢[10-11]。
青藏高原被稱為世界“第三極”,具有獨特的自然地理條件,對于此研究區植物物種多樣性的研究具有重要的意義。但目前針對該區域物種多樣性海拔分布格局的研究不多,楊元合等[12]研究表明,青藏高原高寒草地植物物種豐富度隨海拔的上升呈減少趨勢,而賀連選和劉寶漢[13]對互助縣的研究表明,隨著海拔梯度的逐漸升高,多樣性指數和均勻度增加,物種豐富度先增加后減少。藏北地區位于青藏高原腹地,地域廣闊,地形復雜,氣候資源呈多樣性和復雜性的特點,平均海拔在4 500m以上,被稱為“世界屋脊”的屋脊。不同海拔高度上溫度、濕度和光照等多種環境因子的不同,使其草地物種多樣性也存在著明顯的差異。本研究以藏北地區那曲縣念青唐古拉山脈日囤布山的高寒草地為試驗樣地,通過研究植物群落物種多樣性的海拔梯度變化格局,分析藏北高寒草地植物群落特征及其物種多樣性對海拔變化的響應,為該地區高寒草地生物多樣性的保護和生態恢復提供科學依據和理論基礎。
1 材料與方法
1.1 研究區概況 藏北(那曲)地區位于西藏岡底斯山和念青唐古拉山以北的廣闊地區,83°41′~95°10′E,30°27′~35°39′N,是我國長江、怒江、瀾滄江等主要江河的發源地之一[14]。土地總面積約為44.6萬km2,占西藏自治區總面積的37.1%。該地區草地總面積約為42.1萬km2,占整個地區土地總面積的94.4%[15]。豐富的草地資源和獨特的氣候條件、地理位置等,不僅對該區畜牧業發展以及生態系統的穩定性起著舉足輕重的作用,而且對全國的江河、氣候與環境都有直接或間接的影響,甚至對全球也有不容忽視的作用[16]。那曲縣位于西藏自治區北部的青藏高原腹地,地處唐古拉山脈與念青唐古拉山脈之間。土地總面積160.44萬hm2,總草地面積為138.72萬hm2,占總面積的86.5%,是重要的畜牧業生產基地。該地區氣候寒冷干燥,多大風,屬亞寒帶季風氣候,平均氣溫-1.9℃,年降水量300~400mm,蒸發量1 799.6mm[17]。本試驗樣地選擇在藏北地區那曲縣念青唐古拉山脈日囤布山的高寒草地,其建群種主要為青藏苔草(Carex moorcroftii)、矮生嵩草(Kobresia humilis)、早熟禾(Poa annua)、紫花針茅(Stipapurpurea)、大花嵩草(K.macrantha)和鵝絨委陵菜(Potentilla anserina)等。
1.2 試驗方法 于2009年8月,在草地植被生物量高峰期采用梯度格局,對那曲縣念青唐古拉山脈日囤布山的高寒草地地上生物量及其植物群落物種多樣性沿海拔梯度變化格局進行調查分析。由山腳(海拔高度為4 523m)至山頂(海拔高度為4 673 m)沿一條樣線,每隔50m設置一個海拔梯度,共設置4個梯度,其分別為4 523、4 573、4 623、4 673m(表1),同時用GPS測量記錄樣地的經緯度。每一海拔梯度在約為50m×50m的樣地上隨機選取6個面積為0.5m×0.5m的樣方,先記錄每個樣方內各個物種的高度、蓋度和密度,然后齊地面分種收集地上生物量,稱鮮質量后再置于105℃烘箱中殺青0.5h,以70℃恒溫烘至質量不變,稱干質量。
1.3 數據處理
1.3.1 各物種重要值[18]
物種重要值(IV)= (相對蓋度+相對頻度+相對高度)/3;
相對重要值(Pi)=IV/∑IV。

表1 樣地基本信息
1.3.2 物種多樣性 本研究中,物種豐富度用0.25 m2樣方內出現的物種數表示。物種多樣性的測定采用物種豐富度指數、Shannon-Wiener多樣性指數(H)和E.Pielou均勻度指數(E)表征[19-21]。
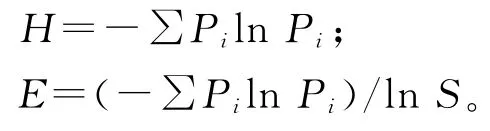
式中,S為群落物種數目,Pi為物種的相對重要值。1.4數據分析 用Excel和Access對數據進行初步整理,將各海拔梯度上所采的樣方數據計算均值,通過SAS 9.0軟件,利用單因子方差分析(ANOVA)和最小顯著差數法(LSD)對不同海拔梯度下各指標間進行比較和差異顯著性檢驗。
2 結果
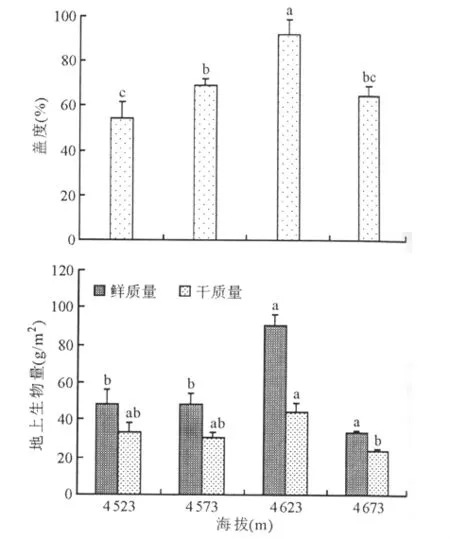
圖1 不同海拔梯度高寒草地植物群落的地上生物量及其蓋度變化
2.1 不同海拔梯度上藏北高寒草地植物群落地上生物量及其蓋度變化特征 隨著海拔的逐漸升高,草地植被群落總蓋度呈現先增加后降低的趨勢(圖1)。不同海拔梯度中,以海拔4 623m的植被蓋度最高,超過了90%,與其他處理差異顯著(P<0.05)。不同海拔梯度上,高寒草地植被地上生物量與總蓋度的變化規律相似,即先增加后逐漸減少。隨著海拔的升高,地上生物量鮮質量依次為47.73、48.00、90.40和33.07g/m2。其中在4 523與4 573m下幾乎沒有變化,而隨著海拔繼續升高,呈現先增加后降低的趨勢。海拔4 623m與前兩個梯度相比,鮮質量與干質量分別增加了88.3%~173%、33.8%~90.2%。由此可見,在藏北高海拔地區的氣候環境中,不同海拔梯度對植物群落的蓋度及其地上生物量影響顯著,而海拔高度為4 623 m時,高寒草地植物群落的蓋度和地上生物量均為最高,說明在該地區,中等海拔梯度(4 623m)水平最適宜草地植被的生長。
2.2 不同海拔梯度上高寒草地植物物種組成及其重要值的變化特征 通過對不同海拔梯度上植物物種重要值的分析,發現高寒草地植物群落物種組成及重要值存在很大的差異(表2)。在海拔4 523m處,建群種主要為青藏苔草和早熟禾等禾草類,伴生種有短穗兔耳草、海乳草以及少量的高原毛茛。由于該樣地離居民點最近,草地出現了不同程度的退化,其植被組成較單一。隨著海拔高度的增加,在海拔高度4 573m處,主要建群種為大花嵩草和矮生嵩草,次優勢種為青藏苔草和早熟禾,伴生種主要為鵝絨委陵菜、肉果草和高山紫菀。在該海拔梯度,莎草類植物優勢度明顯增加,其中出現了矮生嵩草和大花嵩草等莎草科植物。在海拔高度4 623m處,草地植物物種種類最多,主要建群種和伴生種均發生了改變。其主要建群種為矮生嵩草、紫花針茅和鵝絨委陵菜,伴生種主要為矮小藍鐘花、滇紫草和星狀風毛菊等雜類草。莎草類和雜類草的優勢度總和在該梯度的比例明顯增高。在4 673m海拔梯度下,建群種主要為矮生嵩草、鵝絨委陵菜和青藏苔草等,體現了隨海拔高度的增加,雜類草的重要值表現出明顯升高的趨勢。
2.3 藏北高寒草地植物群落物種多樣性沿海拔梯度的變化特征 不同海拔梯度上高寒草地植物群落物種豐富度、多樣性和均勻度均具有顯著差異(圖2)。在海拔相對較低的層次物種豐富度較低;隨著海拔高度的增加,物種豐富度呈先增加后減少的趨勢。即海拔從4 523m升高至4 573m時,植物物種豐富度增加,至海拔4 623m處達到最高(15種);之后隨海拔高度的繼續增加,物種豐富度逐漸降低。多樣性指數在4 623m海拔梯度下最高,為3.544;其次為海拔4 673m,其多樣性指數為2.061;相對較低的海拔4 523與4 573m 處的多樣性指數較低,分別為0.974和1.582。E.Pielou均勻度指數變化趨勢與上述規律一致,即呈現海拔4 623m(1.309)>海拔4 673m(0.895)>海拔4 573m(0.813)>海拔4 523m(0.605)的變化格局。海拔高度與物種豐富度、多樣性指數和均勻度指數之間的趨勢模擬均呈負二次函數關系(圖2),其 擬 合 系 數 r 值 分 別 為 0.825 8、0.825 2 和0.852 1。由分析可知,總體物種多樣性隨海拔高度的變化呈現“單峰”分布格局,即隨著海拔的升高,物種豐富度、多樣性和均勻度均先增加后降低,中等海拔高度(4 623m)時植物群落物種多樣性最高。

表2 不同海拔梯度上高寒草地植物群落物種組成及其重要值的變化
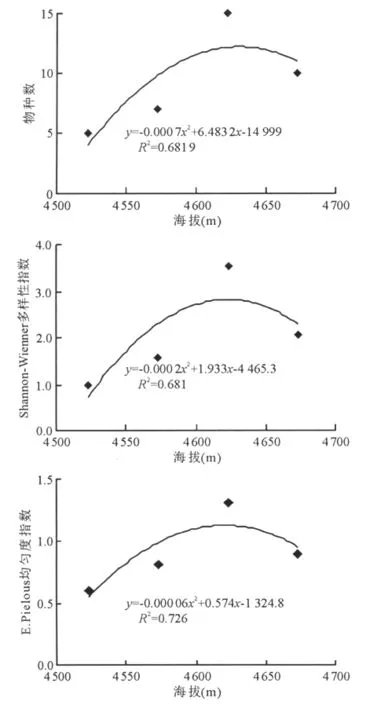
圖2 藏北高寒草地植物群落物種多樣性沿海拔梯度變化趨勢
3 討論與結論
青藏高原獨特的地理環境和氣候條件,使該區域成為一個具有全球意義的脆弱生態系統[22]。海拔高度的變化首先引起溫度、降水、大氣成分等差異,從而影響土壤水分和太陽輻射照度等,進而影響植物群落組成和物種分布[23-24],因此,研究物種多樣性的海拔梯度格局對于揭示生物多樣性的環境梯度變化具有重要的意義。本研究表明,不同海拔梯度對草地植物群落生物量及其蓋度具有顯著的影響。青海瑪沁縣高寒草甸群落的地上生物量隨著海拔的升高逐漸減少[25]。在本研究中,隨著海拔的升高,高寒草地植物群落的總蓋度、地上鮮生物量均表現出先增加后降低的趨勢。原因可能是在海拔4 523 m高度下的草地離居民點較近,受人和牲畜的影響較大,植被稀疏,組成單一,且產量很低,該處的高寒草地已出現了嚴重的退化。隨著海拔的升高,人為因素的干擾逐漸減少,草地群落優勢種植物增多,生物量也明顯增加,至4 623m海拔高度下達到最大值。隨著海拔的進一步增高,環境因素逐漸成為草地生長的主要限制因子,群落生物量及其蓋度顯著下降。綜合表明,在本研究中海拔高度4 623m水平最適應草地植物的生長。高寒草地植物群落物種組成及其重要值沿海拔梯度變化存在很大的差異。隨著海拔的升高,植被群落中出現了莎草類和雜草類植物,而且其重要值所占的比例也明顯增加。
由于影響草本植物分布的因素較多,草地植物群落物種多樣性隨海拔梯度變化的規律非常復雜,不同研究尺度上得出的結果不盡相同[4,26]。賀金生和陳偉烈[26]將物種多樣性隨海拔梯度的變化歸納為5種形式,分別為:隨著海拔的升高而降低(負相關)、先升高后降低(中間高度膨脹型)、先降低后升高、單調升高(正相關)和無規律性(無關)。在本研究中,高寒草地總體物種多樣性隨海拔變化呈現“單峰”分布格局,即隨著海拔的升高,物種豐富度、Shannon-Wiener多樣性指數和E.Pielous均勻度指數均表現出先增加后降低的趨勢,中海拔高度植物群落物種多樣性最高,即“中間高度膨脹型”,這與王國宏[27]在祁連山區的研究結論相一致。在低海拔區域,較頻繁的人為干擾、破壞和降水的限制導致該區域物種多樣性較低[28]。一般而言,隨著海拔高度的增加,氣溫直減率為每100m降低0.5~0.7℃[23]。在高海拔地區,氣溫的逐漸降低使非耐寒草本物種受到熱量的限制而減少,物種多樣性也相應降低。而在中等海拔高度,一方面,由于沒有人為干擾和破壞,草地植物群落生長旺盛;另一方面,由于具有植物所需要的適宜的水熱配置,該區域具有豐富的物種多樣性分布[10,29]。本研究中那曲縣唐古拉山脈日囤布山低海拔區域由于接近居名點,一定程度上受到人為因素的干擾,物種多樣性相對較低,因此,低海拔區域草地植被的保護對于草地生產力的維持和生物多樣性的穩定發展具有重要的意義。
在全球氣候變化背景下,研究物種多樣性隨海拔梯度的變化在一定程度上可以用于探討物種多樣性對極端氣候變化的響應與適應。基于青藏高原氣候的復雜性和脆弱性等特征,開展高寒草地恢復與生物多樣性保護等方面的研究面臨著諸多的問題和挑戰。由于受到青藏高原這一特殊生境的影響,物種多樣性沿海拔梯度變化格局可能還受到其他環境因子的影響。本研究可能還受到研究區相對海拔高度的限制,由于沒有足夠長的梯度,分布格局變化可能具有不確定的因素[30]。因此,開展不同尺度的生物多樣性的研究以及它們對全球氣候變化的響應,是未來研究的重要方向。
[1] 周興民.生物多樣性的研究與保護[J].青海環境,1996(4):19-21.
[2] Tilman D,Wedin D,Knops J.Productivity and sustainability influenced by biodiversity in grassland ecosystems[J].Nature,1996,379:718-720.
[3] Tilman D,Reich P B,Knops J,et al.Diversity and productivity in a long-term grassland experiment[J].Science,2001,294:843-845.
[4] 唐志堯,方精云.植物物種多樣性的垂直分布格局[J].生物多樣性,2004,12(1):20-28.
[5] 杜巖功,崔驍勇,葛勁松,等.三江源地區高寒草地群落特征研究[J].草業科學,2010,27(3):9-14.
[6] Gaston K J.Global patterns in biodiversity[J].Nature,2000,405:220-226.
[7] Oommen M A,Shanker K.Elevational species richness patterns emerge from multiple local mechanisms in Himalayan woody plants[J].Ecology,2005,86:3039-3047.
[8] 郝占慶,于德永,楊曉明,等.長白山北坡植物群落α多樣性及其隨海拔梯度的變化[J].應用生態學報,2002,13(7):785-789.
[9] 唐志堯,方精云,張玲.秦嶺太白山木本植物物種多樣性的梯度格局及環境解釋[J].生物多樣性,2004,12(1):115-122.
[10] Wang G H,Zhou G S,Yang L M,et al.Distribution,species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slopes of Qilianshan Mountains,Gansu,China[J].Plant Ecology,2002,165:169-181.
[11] 王國宏.祁連山北坡中段植物群落多樣性的垂直分布格局[J].生物多樣性,2002,10(1):7-14.
[12] 楊元合,饒勝,胡會峰,等.青藏高寒草地植物物種豐富度及其與環境因子和生物量的關系[J].2004,12(1):200-205.
[13] 賀連選,劉寶漢.不同海拔高度的高寒草甸植物群落多樣性的研究[J].青海畜牧獸醫雜志,2005,35(5):1-4.
[14] 高清竹,萬運帆,李玉娥,等.藏北高寒草地NPP變化趨勢及其對人類活動的響應[J].生態學報,2007,27(11):4612-4619.
[15] 甘肅草原生態研究所,西藏那曲地區畜牧局.西藏那曲地區草地畜牧業資源[M].蘭州:甘肅科學技術出版社,1991:14-33.
[16] 高清竹,李玉娥,林而達,等.藏北地區草地退化的時空分布特征[J].地理學報,2005,60(6):965-973.
[17] 西藏自治區土地管理局,西藏自治區畜牧局.西藏自治區草地資源[M].北京:科學出版社,1994.
[18] 蒙旭輝,李向林,辛曉平,等.不同放牧強度下羊草草甸草原群落特征及多樣性分析[J].草地學報,2009,17(2):239-244.
[19] Zhou Z,Sun O J,Huang J,et al.Land use affects the relationship between species diversity and productivi-ty at the local scale in a semi-arid steppe ecosystem[J].Functional Ecology,2006,20:753-762.
[20] Pielou E C.Mathematical Ecology[M].New York:John Wiley & Sons.Interscience,1977:291-311.
[21] 袁建立,江小蕾,黃文冰,等.放牧季節及放牧強度對高寒草地植物多樣性的影響[J].草業學報,2004,13(3):16-21.
[22] 曹廣民,林麗,張法偉,等.青藏高原高寒矮嵩草草甸穩定性的維持、喪失與恢復[J].草業科學,2010,27(8):34-38.
[23] Fonda R W,Bliss L C.Forest vegetation of the montane and subalpine zones,Olympic Mountains,Washington[J].Ecological Monographs,1969,39:271-301.
[24] 方精云,沈澤昊,崔海亭.試論山地的生態特征及山地生態學的研究內容[J].生物多樣性,2004,12(1):10-19.
[25] 王長庭,王啟基,龍瑞軍,等.高寒草甸群落植物多樣性和初級生產力沿海拔梯度變化的研究[J].植物生態學報,2004,28(2):240-245.
[26] 賀金生,陳偉烈.陸地植物群落物種多樣性的梯度變化特征[J].生態學報,1997,17(1):91-99.
[27] 王國宏.祁連山北坡中段植物群落多樣性的垂直分布格局[J].生物多樣性,2002,10(1):7-14.
[28] 馮建孟,王襄平,徐成東,等.玉龍雪山植物物種多樣性和群落結構沿海拔梯度的分布格局[J].山地學報,2006,24(1):110-116.
[29] 朱源,康慕誼,江源,等.賀蘭山木本植物群落物種多樣性的海拔格局[J].植物生態學報,2008,32(3):574-581.
[30] Whittaker R H,Willis K J,Field R.Scale and species richness:towards a general,hierarchical theory of species diversity[J].Journal of Biogeography,2001,28:453-470.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
