鋁脅迫對黑麥草種子萌發和幼苗生長的影響
2011-09-19 10:53:52陳志剛張紅蕊周曉紅
水土保持研究 2011年4期
關鍵詞:生長
陳志剛,張紅蕊,周曉紅,張 珂
(江蘇大學 環境學院,江蘇 鎮江 212013)
鋁是地球上含量最多的金屬元素,占地殼總質量的7.45%。同時鋁作為高價、半徑小的元素,易水解形成難溶解的氫氧化鋁,并進行相互轉化,在土壤溶液中非常活躍,尤其是在酸性條件下形成大量的Al3+、Al(OH)+2和Al(OH)2+等,從而對植物產生毒害作用,在土壤 pH=5.0或更低時,毒害作用更強[1]。在我國酸性土壤遍布南方15個省區,總面積達203萬km2,約占耕地面積的21%[2],因此,鋁毒害已成為我國酸性土壤中限制作物生長的最重要的因素之一[3],鋁毒嚴重影響到草本植物的生長。
近年來,我國對草食畜產品的需求不斷增加,因此有必要探究金屬鋁對草本植物萌發及其生長的影響。草本植物黑麥草(Lolium multif lorum),是我國南方廣泛種植的具有經濟價值的栽培牧草,具有生長迅速、根系發達、生物量大、可多次刈割,植物再生能力強等諸多優點。有報道顯示,黑麥草對重金屬具有很強的抗性,且對重金屬有富集作用[4-5],因此可被廣泛用于污染環境的生態修復中。近年來,關于黑麥草對重金屬富集的報道較多,且主要集中在生物化學或分子機制方面,但對于輕金屬鋁脅迫下黑麥草種子萌發的研究較少,而種子萌發及幼苗的生長是作物對外部環境反應的開始,也是植物最早接受金屬脅迫的階段,因此,本試驗研究不同鋁濃度對黑麥草種子萌發和幼苗生長的影響,將為酸性土壤中黑麥草的合理利用以及評價黑麥草對金屬鋁的抗逆性提供一定的理論依據。
1 材料和方法
1.1 植物材料
選取顆粒飽滿、大小均勻的多花黑麥草種子,先用0.1%的 HgCl2消毒10 min,然后用去離子水反復沖洗數次,用濾紙吸干后移入試驗容器中開展試驗。
1.2 試驗方法
試驗容器為加有一層紗布的培養皿。每個培養皿(直徑×高:90 mm×15 mm)中放置100顆種子。根據前期預試驗以及參照文獻[6-8],鋁以 AlCl3的形式加入,鋁濃度梯度設置為0 mg/L(以不添加鋁離子的蒸餾水作為對照)、10,50,100,200,300,500 mg/L共7個處理,然后用0.1 mol/L的 HCl溶液分別調節各濃度處理液 pH至4.5。每組鋁濃度分別設3組重復。隨后將裝有黑麥草種子的培養皿置于培養箱中(溫度 :25/15 ℃日/夜;光周期:16/8 h光/暗;相對濕度70%~85%;光照 558~661μmol/(m2?s),PAR)。每天補加等量的處理液使紗布保持濕潤,同時調節pH為4.5。
1.3 指標測定
1.3.1 發芽率、發芽指數測定及其計算 試驗開始2 d后統計黑麥草種子萌芽數,發芽以胚根突破種皮1 mm長為標準,連續2 d發芽種子數無增長視為發芽完全記發芽率。發芽期間,逐日統計發芽粒數,計算發芽指數。發芽率和發芽指數計算公式如下:發芽率=供試種子的發芽數/供試種子數×100%

式中 :Gt——在 t日的發芽數;Dt——相應的發芽試驗天數。
1.3.2 株高、根長測定 試驗第7天時,各處理組的種子發芽已全部完成,此時每天隨機選取培養皿中10根植株,開始用直尺測定其根長株高的變化情況,連續測定7 d。
1.3.3 質膜透性測定及其計算 試驗開始后,分別在第7天和第13天測定浸種液電導率,并以相對電導率(%)來反映種子的質膜透性 。試驗參照魏林[10]等測定浸種液相對電導率的方法:隨機取培養皿中萌發種子25粒,將其置于100 ml錐形瓶中,加入50 ml蒸餾水,于25℃條件下浸泡 2 h,用 HI8733便攜式電導率儀測定浸泡液的電導率值S1,然后將錐形瓶置于沸水浴中20 min,取出后定容至50 ml,冷卻后測定電導率值 S2,相對電導率=S1/S2×100%,重復3次,取平均值。
1.4 數據分析
試驗數據采用 Microsoft Excel軟件進行處理,并采用SPSS 16.0軟件進行統計分析。
2 結 果
2.1 鋁脅迫對黑麥草種子發芽指數和發芽率的影響
發芽指數是反映種子萌發的快慢和整齊程度的指標。發芽指數高表明該種子發芽所用時間短,發芽速度快。表1為不同鋁濃度下黑麥草種子的發芽率及其發芽指數。由表1可知,10~50 mg/L鋁濃度處理時,黑麥草種子發芽指數與對照無顯著差異(p>0.05);而當鋁濃度大于50 mg/L時,隨著鋁濃度的升高,黑麥草種子萌發的速度明顯變慢(p<0.05),當鋁處理濃度為500 mg/L時,發芽指數僅為對照的2.8%。
同時,由表 1可知,鋁濃度對黑麥草種子的發芽率有一定的影響。隨著鋁處理濃度的升高,黑麥草種子發芽率先升高后降低。當鋁濃度介于10~50 mg/L范圍內,發芽率與對照無顯著差異(p>0.05),其中10 mg/L鋁濃度處理下,發芽率最大,高于對照;之后,隨著鋁處理濃度的升高,種子萌發受到顯著抑制(p<0.05),當鋁濃度為500 mg/L時,發芽率僅為6.3%。

表1 鋁脅迫對黑麥草種子發芽指數和發芽率的影響
2.2 鋁脅迫對黑麥草幼苗株高和根長的影響
2.2.1 鋁脅迫對黑麥草幼苗株高的影響 由圖1可知,隨試驗時間的延長,株高呈上升趨勢,但隨著鋁濃度的增加黑麥草株高增長速度逐漸降低,且鋁濃度越高抑制作用越明顯。具體表現為:在試驗初始的7~9 d,50 mg/L鋁處理下株高與對照差異顯著(p<0.05),比對照降低38.8%~50.5%;而試驗的第10~13 d時,10 mg/L鋁處理顯著抑制植株生長(p<0.05),而50 mg/L鋁處理下株高比對照降低51.0%~57.0%。
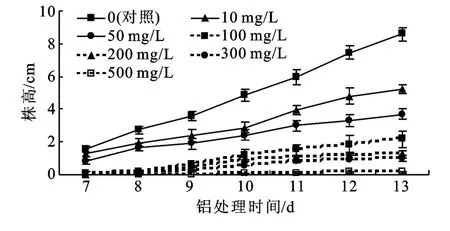
圖1 不同濃度鋁處理下黑麥草株高變化
2.2.2 鋁脅迫對黑麥草幼苗根長的影響 由圖2可知,當鋁濃度介于0~50 mg/L時,黑麥草幼苗在試驗第7天有根的生長,但隨著試驗時間的延長,10 mg/L、50 mg/L試驗組的根系增加非常緩慢,試驗第10天時,兩組幼苗根長僅增加了21.0%和16.8%;而試驗第13天時,兩組幼苗根長僅增加了26.0%和19.3%,與對照組差異顯著(p<0.05),對照組根長增加2.58倍。當鋁濃度大于50 mg/L時,黑麥草幼苗無根的生長。

圖2 不同濃度鋁處理下黑麥草根長變化
2.3 鋁脅迫對黑麥草種子質膜透性的影響
本文中黑麥草種子質膜透性采用相對電導率來表示,相對電導率高,質膜透性大,植株受害嚴重。由圖3可知,10 mg/L的鋁濃度顯著降低了黑麥草種子質膜透性(p<0.05),在鋁處理的第7天和第13天,相對電導率比對照降低44.2%和50.7%;而隨著鋁濃度的升高,相對電導率顯著增加(p<0.05),試驗第7天,50,10,200,300,500 mg/L鋁濃度處理組相對電導率較對照增加 0.7倍、1.2倍、2.4倍、2.9倍、4.2倍,且隨著試驗時間的延長,在50~100 mg/L鋁濃度處理下,各試驗組相對電導率增加顯著(p<0.05)。

圖3 鋁脅迫下黑麥草浸種液相對電導率變化
3 討 論
鋁是植物生長的非必須元素,通常以難溶性硅酸鹽或氧化鋁形式存在,對植物沒有毒害,但 p H<5時,可溶性鋁含量將增加[1],從而將影響植物種子的萌發和幼苗生長。本次試驗結果表明,10~50 mg/L的鋁處理對黑麥草種子萌發無明顯影響(p>0.05),但隨著鋁濃度的升高,種子萌發將受到顯著抑制(p<0.05)。孫冬花等[11]的研究顯示,鋁處理下秋葵(Hibiscusmoscheutos L)和兩個小麥(Triticum aestivum L)品種種子的萌發率均沒有顯著變化。而張芬琴[12]和劉鵬[13]等的研究表明低濃度的鋁促進種子萌發,高濃度鋁抑制種子萌發,這表明不同物種的種子萌發對鋁脅迫存在不同的反應機制[11]。
研究表明,植物根系是鋁毒害的最初作用點。本文研究表明,隨著鋁濃度的增加,黑麥草幼苗株高、根長呈顯著的下降趨勢(p<0.05),10 mg/L的鋁處理即可抑制黑麥草株高和根系的生長,表明黑麥草株高和根系對鋁脅迫的反應較為敏感,這與劉強[14]等關于鋁毒脅迫下油菜(Brassica campestris L)幼苗生長的研究相似。另外,本次試驗中當鋁處理濃度 ≥100 mg/L時,黑麥草根系停止生長,表明鋁對根系的抑制作用更為直接和明顯。
質膜包在細胞外面,所以又稱細胞膜,它不僅是區分細胞內部與周圍環境的動態屏障,更是細胞物質交換和信息傳遞的通道,其穩定性是細胞進行正常生理功能的基礎。逆境脅迫下,鋁毒與細胞膜作用,改變膜透性,使細胞內物質向外滲漏作用加強,用于植株正常生長發育的同化物減少,植物生長減緩,生物量下降。本研究顯示,10 mg/L鋁處理能顯著降低質膜透性(p<0.05);之后隨著鋁濃度的升高質膜透性明顯變大(p<0.05),這與王芳[15]等的研究結果相一致。而張芬琴[12]等的研究結果顯示,低濃度的鋁浸種后,苜蓿(Medicago sativa)種子的電解質外滲率微高于對照,高濃度鋁浸種后,其電解質外滲率明顯高于對照處理。Ishikawa S等的研究同樣表明:鋁毒直接作用于根系,使根系生長減慢或停止。同時,鋁毒與細胞膜作用,改變膜透性,使細胞內物質向外滲漏作用加強[16],表明不同物種的細胞對鋁脅迫的反應不同。
綜上,多花黑麥草在一定程度上具有耐鋁性,但隨著鋁濃度升高,鋁毒害程度變大。因此,對于鋁濃度較低的酸性土壤,可以考慮栽培黑麥草。但關于鋁脅迫下,黑麥草生理特性的變化及其對鋁的富集情況有待進一步研究,從而為黑麥草對金屬鋁污染的酸性土壤進行生態修復提供理論支撐。
[1] Larsen P B,Stenzler L M,Tai C Y,et al.Molecular and physiological analysis of Arabidopsis mutants exhibiting altered sensitivities to aluminum[J].Plant Soil,1997,192(1):3-7.
[2] 林咸永,王建林.植物對Al毒脅迫的適應機制[M]//張福鎖.植物營養的生態生理學和遺傳學.北京:中國科學技術出版社,1993:248-290.
[3] Taylor G J.Aluminum toxicity and tolerance in plants[J].Biological and Ecological Effects,1989,107(2):327-361.
[4] 廖敏,黃昌勇.黑麥草生長過程中有機酸對鎘毒性的影響[J].應用生態學報,2002,13(1):109-112.
[5] 徐衛紅,熊治庭,王宏信,等.鋅脅迫對重金屬富集植物黑麥草養分吸收和鋅積累的影響[J].水土保持學報,2005,19(4):32-35.
[6] Jones D L,Prabowo A M,Kochian L V.Aluminum-organic acid interactions in acid soil[J].Plant Soil,1996,182(2):229-237.
[7] Gunse B,Garzon T,Barcelo J.Study of Aluminum toxicity by means of vital staining profiles in four cultivars of Phaseolus vulgaris L.[J].Plant Physiol,2003,160(12):1447-1450.
[8] Flaten T P.Aluminum in tea-concentrations,speciation and bioavailability[J].Coord.Chem.Rev.,2002,228(2):385-395.
[9] Silva I R,Smyth T J.Differential aluminum tolerancein soybean:an evaluation of the role of organic acids[J].Physiol.Plant,2001,112(2):200-210.
[10] 魏林,梁志懷,曹福祥,等.哈茨木霉 T2-16代謝產物對花生種子活力和抗黃曲霉菌浸染能力的影響[J].中國油料作物學報,2009,31(3):370-373.
[11] 孫冬花,田秋英,張文浩.鋁對秋葵、小麥種子萌發和幼苗生長的影響[J].云南植物研究,2006,28(5):523-528.
[12] 張芬琴,于愛蘭.鋁處理對苜蓿種子萌發及其幼苗生理生化特性的影響[J].草業學報,1999,8(3):61-65.
[13] 劉鵬,徐根娣,姜雪梅,等.鋁對大豆種子萌發的影響[J].種子,2003,22(1):30-32.
[14] 劉強,龍婉婉,胡萃,等.鋁脅迫對油菜種子萌發和幼苗生長的影響[J].種子,2009,28(7):5-10.
[15] 王芳,劉鵬,徐根娣,等.鋁對蕎麥根系的影響[J].廣西植物,2006,26(3):321-324.
[16] Ishikawa S,Wagatsuma T,Ikarashi T.Comparative toxicity of Al3+,Yb3+,and La3+to root-tip cells differing in tolerance to high Al3+in terms of ionic potentials of dehydrated trivalent cations[J].Soil Sci.Plant Nutr.,1996,42(3):613-625.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
