玉米彎孢葉斑病菌ATMT轉化條件優化及轉GFP單孢的獲得
2011-10-10 02:53:26陳茂功林小虎李洪杰王曉鳴周印富
河北科技師范學院學報 2011年3期
關鍵詞:效率
陳茂功,林小虎,李洪杰,王曉鳴* ,周印富*
(1河北科技師范學院生命科技學院,河北秦皇島,066004;2中國農業科學院作物科學研究所)
由新月彎孢(Curvularia lunata(Wakker)Boed)引起的玉米彎孢葉斑病已成為一種重要的葉部病害,對我國玉米生產造成了嚴重的損失,一般會促使玉米減產20%~30%,嚴重為害時減產50%以上,甚至絕收[1~4]。目前對該病害研究主要集中在抗病品種與變異和致病機理等方面[5,6],但對該病菌在玉米葉片上的侵染及發育過程仍不十分清楚。
GFP基因作為一種活體報告基因,其應用已深入到植物病害研究中的各個方面,如對植物病原體的侵染、致病機理的研究[7,8]。本研究擬建立一套農桿菌介導彎孢菌的轉化方法,同時將GFP基因轉入到彎孢葉斑病菌中,獲得該菌轉化子,以便進行侵入機理研究。
1 材料和方法
1.1 菌株
玉米彎孢葉斑病菌(C.lunata)由中國農業科學院作物科學研究所檢疫基地保存;農桿菌菌株為AGL-1,質粒以pCAMBIAI300為骨架構建了pCAMDGFP[9],并含有潮霉素B(HyB)和卡那霉素(kan)抗性,由 Marina Franceschetti(john inner Center,UK)惠贈。
1.2 主要試劑
潮霉素B、頭孢噻肟鈉、卡那霉素、乙酰丁香酮、MES購自北京鼎國生物技術有限責任公司;其他所用試劑均購于北京萬鑫化業商貿中心。
1.3 方法
1.3.1 頭孢噻肟鈉對農桿菌抑菌濃度的測定 將農桿菌接種于LB液體培養基中,37℃搖培至OD600約為0.6后,取100 μL分別涂于含有頭孢噻肟鈉 (0,50,100,150,200 mg/L)的 PDA 培養基上,每個濃度3次重復,25℃下培養4 d后觀察農桿菌菌落的生長情況,確定頭孢噻肟鈉的抑菌濃度。
1.3.2 潮霉素B對彎孢菌菌落生長的抑制測定 在含不同質量濃度潮霉素B(0,30,50,75,100,150 mg/L)的PDA培養基上,接種直徑4 mm的菌餅,每個濃度3次重復,25℃下培養6 d,每隔36 h測量1次菌落直徑,觀察潮霉素B對菌落生長的抑制。
1.3.3 潮霉素B對彎孢菌分生孢子萌發的抑制測定 取100 μL的分生孢子懸濁液(109個/L)分別涂于含有潮霉素 B(0,30,50,75,100,150 mg/L)的 PDA 培養基上,每個濃度3次重復,25 ℃下培養48 h,觀察潮霉素B對分生孢子萌發的抑制。
1.3.4 不同轉化條件對彎孢菌ATMT轉化效率的影響 轉化方法參照趙培寶等的方法進行[10],并稍作修改。取誘導后的農桿菌菌液與萌發彎孢葉斑病菌分生孢子懸浮液等體積混合,均勻涂布于IM固體培養基的微孔濾膜上,黑暗培養一定時間后將濾膜轉移到含有200 mg/L的頭孢噻肟鈉和50 mg/L潮霉素的PDA培養基上,25℃培養7 d后將生長出來的菌落轉移到含有50 mg/L潮霉素的PDA培養基上進行二次篩選。
①孢子萌發時間。采用萌發0,6,12,24 h的分生孢子分別進行遺傳轉化,其它操作同上。
②共培養介質。分別覆有玻璃紙、微孔濾膜、定性濾紙的共培養基進行共培養,其它操作同上。③共培養溫度和時間。分別設置共培養溫度20,23,25,28℃,培養24,48,72 h,其它操作同上。
④受體材料。參照王賡等的方法制備彎孢菌原生質體[11]。分別采用孢子懸浮液、菌絲、原生質體進行遺傳轉化,其它操作同上。以上試驗均設置3次重復。
1.3.5 轉化子穩定性分析 用單孢分離的方法獲得轉化子,隨機選擇10個轉化子分別在含有50 mg/L潮霉素和不含有潮霉素的PDA平板上連續傳代5代,然后接種于含有50 mg/L潮霉素的平板上,觀察轉化子是否生長及性狀是否穩定。
1.3.6 轉化子的PCR檢測和熒光觀察 隨機挑取遺傳穩定的10個轉化子菌落,以野生型彎孢菌為對照,在PDA培養基上培養7 d后收集菌絲體。分別提取基因組DNA后進行gfp基因和潮霉素B抗性基因的 PCR檢測,程序為:95℃預變性10 min;95℃變性30 s,56℃復性30 s,72℃延伸2 min,35個循環;72℃延伸10 min;經10 g/L瓊脂糖電泳檢測。挑取轉化子的菌絲制成玻片,在Olympus熒光顯微鏡下進行熒光觀察,同時以野生型菌株為對照。
1.3.7 轉化子致病性的測定 感病自交系黃早四長至四葉期,以接種液(蔗糖:20 g/L;Tween 20:0.2 g/L;無菌水)調節病菌分生孢子濃度至1.0×109個/L,用高壓噴霧裝置噴霧接種。實驗選取5個轉化子,設置陰性對照(無菌接種液)及陽性對照(野生型菌株孢子懸液)。接種處理后黑暗保濕48 h。每天觀察植株發病情況并記錄。
2 結果和分析
2.1 頭孢噻肟鈉對農桿菌抑菌濃度的測定
農桿菌菌液涂于含有不同濃度的頭孢噻肟鈉的PDA培養基中25℃培養4 d后,在頭孢噻肟鈉質量濃度為0,50,100 mg/L的處理中均出現菌落,其他濃度無菌落出現。當頭孢噻肟鈉為150 mg/L時,在短時間內可抑制農桿菌的生長。但考慮彎孢菌ATMT的轉化周期及細菌生長特性,本次試驗選用比臨界濃度稍高的抗生素濃度,確定頭孢噻肟鈉質量濃度為200 mg/L作為病菌ATMT轉化時農桿菌抑菌濃度。
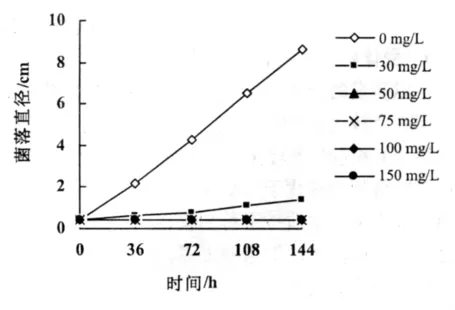
圖1 不同濃度潮霉素B對于菌落生長的抑制影響
2.2 潮霉素B最適濃度的選擇
潮霉素B可以有效地抑制真菌分生孢子的萌發。將彎孢菌的分生孢子接種于5種不同的潮霉素濃度的PDA培養基中25℃連續培養2 d。當潮霉素質量濃度達到50 mg/L以上時,分生孢子萌發受到完全抑制。將直徑4 mm的彎孢菌落接種于不同的潮霉素B濃度的PDA培養基中,25℃下連續培養6 d。當質量濃度到達50 mg/L時,菌落的生長受到了完全的抑制。本試驗確定以50 mg/L作為篩選轉化子的有效濃度(圖1)。
2.3 分生孢子萌發時間對轉化效率的影響
采用不同萌發程度的分生孢子進行轉化。利用未萌發的分生孢子進行轉化得到32個轉化子;利用萌發6 h的孢子轉化得到67個轉化子;利用萌發12 h的孢子得到106個轉化子,利用萌發24 h的孢子得到71個轉化子。萌發12 h的孢子的轉化效率是最高的,說明萌發的孢子有利于農桿菌轉化,萌發12 h的孢子是農桿菌介導轉化的最佳材料(圖2)。
2.4 共培養介質對轉化效率的影響
在定性濾紙上進行共培養轉化效率為55個轉化子/106個孢子,顯著低于在微孔濾膜和玻璃紙;微孔濾膜的轉化效率(108個轉化子/106個孢子)高于玻璃紙的轉化效率(98個轉化子/106個孢子)(圖3)。

圖2 孢子萌發時間對轉化效率的影響
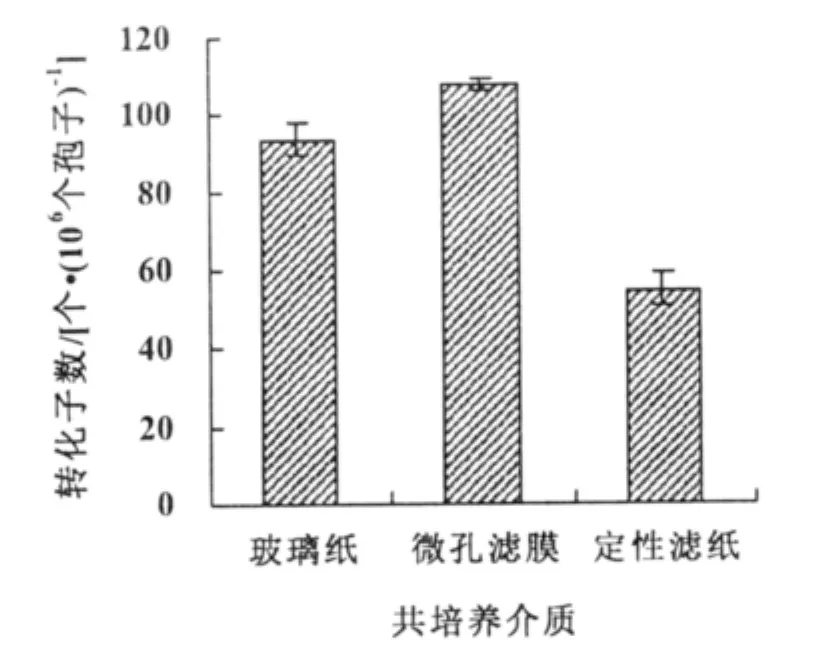
圖3 介質類型對轉化效率的影響
2.5 共培養溫度和時間對轉化效率的影響
當共培養溫度為19℃和28℃時,轉化效率較低。在共培養時間為24 h時,在19,23,25,28℃分別得到了43,64,78,58個轉化子;在共培養時間為48 h時,分別得到58,89,106,78個轉化子;在共培養時間為72 h時,分別得到47,79,67,43個轉化子。結果表明,農桿菌介導的遺傳轉化最佳條件為共培養溫度25℃,共培養時間48 h(圖4)。
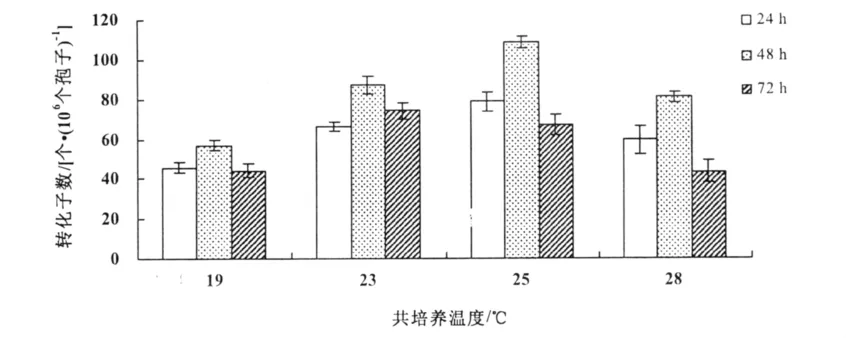
圖4 共培養時間和溫度對轉化效率的影響

圖5 受體材料對轉化效率的影響
2.6 受體材料對轉化效率的影響
以萌發的分生孢子為受體材料進行轉化,轉化效率可達108個轉化子/106個孢子,顯著高于以原生質體為受體的轉化效率(65個轉化子/106個原生質體),而以菌絲為受體材料得到的轉化子為0(圖5)。原生質體轉化效率低可能是彎孢菌原生質體不易制備,且數量較少;菌絲轉化為0可能是彎孢菌菌絲細胞壁太厚,農桿菌不易侵染,不能作為農桿菌轉化的受體材料。
2.7 彎孢菌轉化子穩定性分析
隨機選擇的10個轉化子,在PDA和含50 mg/L潮霉素的PDA上分別轉代5次后轉接到含50 mg/L潮霉素的PDA觀察菌落生長。結果表明:轉代后轉化子都能在含潮霉素B的培養基上生長,且菌落形態正常,表明轉化子在遺傳上是穩定的。
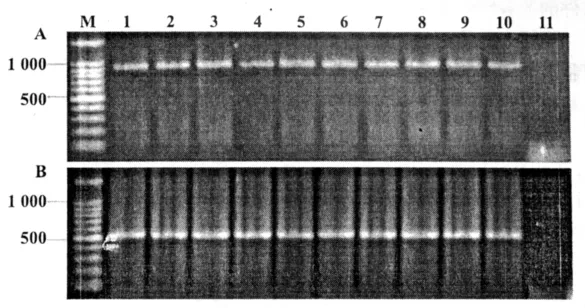
圖6 彎孢菌轉化子中gfp基因、潮霉素B抗性基因的PCR擴增A為潮霉素B抗性基因擴增產物;B為gfp基因擴增產物;M為標準分子量;1~10為轉化子的擴增產物;11為野生型對照
2.8 彎孢菌轉化子的PCR檢測
以彎孢菌轉化子和野生菌株的基因組DNA為模板進行PCR檢測,結果10個轉化子均擴增出gfp基因和潮霉素B抗性基因,而野生菌株則沒有任何擴增產物(圖6)。
2.9 彎孢菌轉化子的熒光觀察
挑取轉化子G1和野生菌株的菌絲在488 nm波長下進行熒光觀察,結果野生菌株沒有觀察到任何熒光,轉化子的菌絲和分生孢子均能發出綠色熒光(圖7)。

圖7 綠色熒光蛋白在彎孢菌轉化子中不同時期的表達
2.10 轉化子致病性的測定
接種實驗結果,野生型菌株、轉化子1和轉化子2表現出較強的毒性和侵染力,在接種后第3天,寄主植物黃早四即開始產生典型病斑,第4天癥狀明顯加重,第5天病害級別即達到3級~4級;轉化子3和轉化子4也可以在寄主植物上引起典型病斑,但是菌株侵襲力和致病力較弱,分別在第4天、第5天開始出現癥狀且發病輕,第5天病害級別為1級~2級;轉化子5則致病力完全喪失(圖8)。

圖8 玉米孢子懸濁液噴霧接種結果
3 討 論
農桿菌介導轉化是農桿菌與真菌相互作用的結果,凡能影響真菌轉化、農桿菌侵染以及轉化子再生的各種因素都會影響其轉化效率。因此,農桿菌轉化效率的提高依賴于對各種影響因子的優化和轉化條件的改善[12,13]。
不同真菌使用抗生素的濃度不同。完全抑制木霉生長的潮霉素B最低質量濃度為150 mg/L,完全抑制玉米大斑病菌的潮霉素B為100 mg/L。本研究中抑制彎孢菌生長的潮霉素B質量濃度為50 mg/L,相對于已報道的真菌抑制生長潮霉素B濃度是比較低的[14,15]。
受體材料是農桿菌轉化的關鍵步驟。原生質體、分生孢子、菌絲乃至蘑菇子實體均可作為受體材料,某些真菌以原生質體或分生孢子為受體材料均可成功轉化,而有些真菌轉化只能以原生質體作為受體材料[16~21]。本研究表明,以萌發12 h的分生孢子為受體材料,轉化效率高。萌發時間過短或過長都會降低轉化效率,這可能由于彎孢葉斑病菌分生孢子壁較厚,而萌發的芽管在一定時間內管壁較薄,農桿菌易侵染;萌發時間過長,管壁增厚,使農桿菌不易侵染所致。
共培養條件也是影響轉化的關鍵,包括共培養時間及溫度、共培養介質等。共培養時間的增加會提高轉化效率,但時間過長會導致T-DNA插入的拷貝數增加;時間過短,農桿菌侵染受體材料尚未完成[22]。過高的轉化溫度會使農桿菌的轉化功能喪失,即T-DNA插入宿主能力的丟失,導致轉化效率降低;過低的溫度會導致各種菌體活性降低[22]。對于彎孢葉斑病菌而言,在25℃黑暗條件下共培養48 h轉化效率最高。本試驗還發現,以微孔濾膜為介質轉化效率最高,可能由于微孔濾膜有較好的通透性,使彎孢葉斑病菌分生孢子與農桿菌均獲得充足營養。
本研究利用農桿菌介導的轉化系統,成功將綠色熒光蛋白標記基因插入到彎孢葉斑病菌的基因組DNA中,獲得了gfp穩定遺傳的彎孢葉斑病菌突變株,為研究該病原菌的侵染循環和致病機制提供了基礎性研究材料。通過本研究建立的彎孢菌轉化系統還可以用來進行其功能基因的研究。農桿菌介導轉化以T-DNA隨機插入受體菌基因組中,從而可能導致相應功能的喪失或減弱,進而根據表型變化來研究該基因的功能,為開展玉米彎孢葉斑病菌基因功能的研究奠定了基礎。
[1]戴法超,高衛東,王曉鳴,等.玉米彎孢菌葉斑病的初步研究簡報[J].植物保護,1996,22(4):36-37.
[2]戴法超,王曉鳴,朱振東,等.玉米彎孢菌葉斑病研究[J].植物病理學報,1998,28(2):123-129.
[3]呂國忠,劉志恒,何富剛,等.遼寧省爆發一種新病害-玉米彎孢菌葉斑病[J].沈陽農業大學學報,1997,28(1):75-76.
[4]MACRI F,LENNA P.Leaf corn blight incited by Curvularia lunata(Wakk.)Boed[J].Jounral of Plant Pathology,1974,10:27-35.
[5]XU S F,CHEN J,LIU L X,et al.Proteomics associated with virulence differentiation of Curvularia lunata in maize(Zea maydis)in China[J].J Integr Plant Biol,2007,49(4):487-496.
[6]LIU T,LIU L X,JIANG X,et al.A new furanoid toxin produced by Curvularia lunata,the causal agent of maize Curvularia leaf spot[J].Can J Plant Pathol,2009(31):22-27.
[7]汪恒英,周守標,常志州,等.綠色熒光蛋白(GFP)研究進展[J].生物技術,2004,14(3):70-71,73.
[8]吳瑞,張樹珍.綠色熒光蛋白及其在植物分子生物學中的應用[J].分子植物育種,2005,3(2):240-244.
[9]SESMA A,OSBOURN,A E.The rice leaf blast pathogen undergoes developmental processes typical of root-infecting fung[J].Nature,2004,431:582-586.
[10]趙培寶,周慶新,任愛芝,等.根癌農桿菌介導的輪枝鐮孢菌遺傳轉化及T-DNA插入突變體的篩選[J].菌物學報,2008,27(2):258-266.
[11]王賡,杜連祥.新月彎孢霉原生質體制備及再生條件的研究[J].微生物學通報,1999,26(1):21-23.
[12]方為國,張永軍,楊星勇,等.根癌農桿菌介導真菌遺傳轉化的研究進展[J].中國生物工程雜志,2002,22(5):40-44.
[13]彭姍,李靜,杜娟,等.棉花黃萎病菌遺傳轉化載體構建和農桿菌轉化[J].石河子大學學報:自然科學版,2007,25(1):34-38.
[14]MICHIELSE C B,HOOYKASS P J J,HONDEL C J J,et al.Agrobacterium-mediated transformation as a tool for functionalgenomics in fungi[J].Curr Genet,2005,48:1-17.
[15]李衛,郭光沁,鄭國锠.根癌農桿菌介導遺傳轉化研究的若干新進展[J].科學通報,2000,45(8):798-807.
[16]ABUODEH R O,ORBACH M J,MANDEL M A,et al.Genet-ic transformation of Coccidioides immitisfacilitated by Agrobacterium tumefaciens[J].The Journal of Infectious Diseases,2000,181(6):2 106-2 110.
[17]DE GROOT M J,BUNDOCK P,HOOYKAAS P J,et al.Agrobacteriumtumefaciens-mediated transformation of filamentous fungi[J].Nature Biotechnology,1998,16(9):839-842.
[18]CHI M H,PARK S Y,KIM S,et al.A novel pathogenicity gene is required in the rice blast fungus to suppress the basal defenses of the host[J].PLoS Pathogens,2009,5(4):1-16.
[19]劉朋娟,王政逸,王秋華,等.農桿菌介導的稻瘟病菌轉化及致病缺陷突變體篩選[J].中國水稻科學,2006,20(3):231-237.
[20]孫曉紅,姚泉洪,陳明杰,等.含綠色熒光蛋白及trp3iar基因的草菇表達載體的構建[J].上海水產大學學報,2006,15(1):12-16.
[21]MICHIELSE C B,RAM A F J,HOOYKAAS P J,et al.Roleof bacterial virulence proteins in Agrobacteriumme diated transformation of Aspergillus awamori[J].Fungal Genetics and Biology,2004,45(4):571-578.
[22]張震,杜新法,柴榮耀,等.根癌農桿菌介導絲狀真菌遺傳轉化因素的研究進展[J].微生物學雜志,2007,27(6):88-91.
猜你喜歡
瘋狂英語·初中天地(2021年5期)2021-07-21 02:24:28
甘肅教育(2020年14期)2020-09-11 07:57:42
中學生數理化(高中版.高考數學)(2020年5期)2020-06-02 09:19:08
商周刊(2017年9期)2017-08-22 02:57:49
遼寧經濟(2017年6期)2017-07-12 09:27:16
中國衛生(2016年9期)2016-11-12 13:27:54
時代英語·高二(2015年1期)2015-03-16 00:08:11
中國洗滌用品工業(2015年7期)2015-02-28 19:02:38
電子設計工程(2015年12期)2015-02-27 12:06:10
中國衛生(2014年11期)2014-11-12 13:11:32
