發芽蠶豆富集γ-氨基丁酸的培養液組分優化
2011-11-28 02:28:32楊潤強韓永斌顧振新
中國糧油學報 2011年11期
關鍵詞:模型
陳 惠 楊潤強 韓永斌 顧振新
發芽蠶豆富集γ-氨基丁酸的培養液組分優化
陳 惠 楊潤強 韓永斌 顧振新
(南京農業大學農業部農畜產品加工與質量控制重點開放實驗室,南京 210095)
以蠶豆為試材,研究谷氨酸鈉(MSG)、CaCl2、和VB6對發芽蠶豆谷氨酸脫羧酶(GAD)及γ-氨基丁酸(GABA)的影響,采用Box-behnken設計對發芽蠶豆富集GABA的培養液組分進行了優化,并對發芽蠶豆富集GABA的二次回歸模型進行分析。結果表明,低氧聯合鹽脅迫下,MSG、CaCl2和VB6對發芽蠶豆GAD及GABA的影響均達到顯著水平(P<0.005)。經過回歸分析建立了GABA含量對培養液組分的二次回歸模型,回歸方程的決定系數達到0.976,說明方程能很好的預測GABA富集含量的變化。蠶豆富集GABA的最適培養液組分為MSG 1.1 mg/mL、CaCl26.1 mmol/L、VB672 μmol/L,此時,GABA 富集量達到(1.98 ±0.09)mg/g DW,為對照[(1.08 ±0.01)mg/g DW]的1.83 倍。
發芽蠶豆 γ-氨基丁酸 富集 優化
蠶豆(Vicia faba L.)為豆科野豌豆屬作物,其種子富含蛋白質、淀粉、氨基酸、維生素和鉀、鈣、磷等礦物質元素[1]。蠶豆種子中豐富的蛋白質和氨基酸可為γ-氨基丁酸(GABA)富集提供充足的底物。GABA具有鎮靜安神、抗焦慮、抗驚厥、降血壓等作用[2]。通常植物組織中GABA含量較低,但受到熱激、鹽害和低氧等逆境脅迫時,其含量提高幾倍至幾十倍[3]。低氧脅迫導致植物細胞質酸化[4],使谷氨酸脫羧酶(GAD)活性提高,為GABA的合成提供了條件。鹽脅迫使植物體胞內Ca2+濃度升高[5],CaM(鈣調素)水平提高,GAD活性增加,從而促進GABA積累。因此,低氧脅迫和鹽脅迫是植物源食品富集GABA 的有效方法[6]。Li等[7]研究了溶氧量對發芽蠶豆富集GABA的影響,但關于低氧脅迫聯合鹽脅迫對蠶豆富集GABA的研究未見報道。本研究考察了蠶豆發芽過程中,低氧聯合鹽脅迫處理條件下,MSG、CaCl2和VB6對GAD活性和對GABA富集效果的影響,旨在為蠶豆發芽富集GABA提供科學依據。
1 材料與方法
1.1 材料與儀器
1.1.1 試驗材料
供試蠶豆由江蘇沿江地區農業科學研究所提供,品種為啟豆2號。
γ-氨基丁酸(GABA)標準品:購自Sigma公司,其他試劑均為國產分析純。
1.1.2 試驗儀器
Orion818型pH測試儀:Orion公司;Agilent 1200 Series高效液相色譜儀:Agilent公司;TDL-40B離心機:上海安亭科學儀器廠;HH-6型數顯恒溫水浴鍋:常州國華電器有限公司;PYX-DHS-BS型隔水電熱恒溫培養箱:上海躍進醫療器械廠;Anke GL-20G-Ⅱ冷凍離心機:上海安亭儀器廠;JA2003型電子天平:上海精密科學儀器有限公司。
1.2 試驗設計
1.2.1 單因素試驗
稱取50 g蠶豆,水清洗后,用1%的次氯酸鈉水溶液消毒30 min,然后用蒸餾水洗去殘留的次氯酸鈉。消毒后的蠶豆于蒸餾水中30℃浸泡8 h,取出吸干表面水分,置于托盤中(33±1)℃避光培養1.5 d。取出后置于具塞培養瓶(φ6 cm×9 cm)中,加入60 mmol/L的NaCl溶液(用10 mmol/L檸檬酸-檸檬酸鈉緩沖液配制,pH 3.5)200 mL,于(33±1)℃下避光培養4 d。期間,向培養液中連續通入空氣,通氣量為1.2 L/min,形成低氧聯合鹽脅迫條件。
培養液中谷氨酸鈉(MSG)為0~2.5 mg/mL、CaCl2為0~15 mmol/L、VB6為0~150 μmol/L。
1.2.2 響應面試驗
根據單因素試驗結果,以MSG濃度(A)、CaCl2濃度(B)和VB6濃度(C)3個因素與GABA含量進行響應面試驗設計(表1),優化培養液組分。通過Design Expert軟件對試驗數據進行回歸分析,預測發芽蠶豆富集GABA的最優培養液組成。以pH 3.5檸檬酸-檸檬酸鈉緩沖液培養為對照。
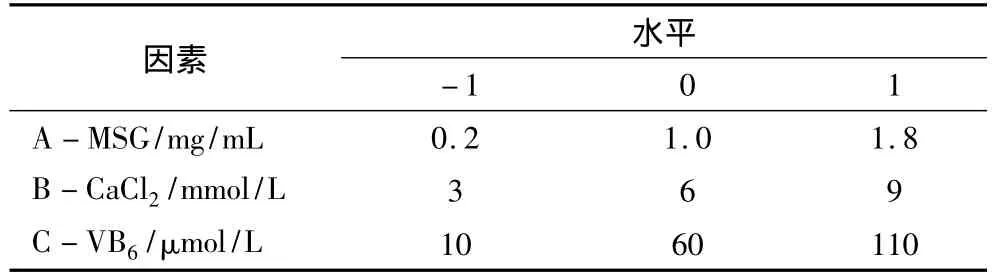
表1 響應面設計因素與水平
1.3 測定指標與方法
GABA含量:參照Bai[8]的方法測定;GAD活性:參照 Li[7]的方法測定。
2 結果與討論
2.1 MSG對發芽蠶豆GABA含量和GAD活性的影響
由表2可見,當MSG質量濃度低于1.0 mg/mL時,發芽蠶豆GABA含量隨著MSG質量濃度的增加而增加,MSG大于1.0 mg/mL時,GABA含量隨MSG質量濃度升高而降低。MSG質量濃度在1.0~2.0 mg/mL時,GABA含量差異不顯著。因此,本研究選擇MSG質量濃度為1.0 mg/mL。GAD活性變化趨勢與GABA含量變化相似。當MSG質量濃度為1.0 mg/mL時,GAD活性最高,為對照的1.43倍。谷氨酸和谷氨酸鈉(MSG)在GAD作用下轉化為GABA,其含量增多可使GABA支路中碳流量增加,并調節GAD 活性[9]。王玉萍等[10]研究表明,添加 MSG 對發芽糙米GABA的富積影響不顯著。本研究與之報道不一致,原因尚有待探討。
2.2 CaCl2對發芽蠶豆GABA含量和GAD活性的影響
植物GAD氨基酸序列中C末端存在CaM結合區,該區域與Ca2+/CaM結合后可提高GAD活性[3]。低氧條件下,添加Ca2+后,發芽粟谷中GAD活性得以提高,因而GABA含量提高[8]。由表3可知,隨著CaCl2濃度的增加,發芽蠶豆GAD活性和GABA含量均呈先升高后降低的變化趨勢。當CaCl2濃度為6 mmol/L時,GAD活性和GABA含量均達到最大值,分別為對照的1.88倍和1.09倍。這一研究結果也證實了培養液中添加適當濃度的CaCl2可提高蠶豆GAD活性和GABA含量。
2.3 VB6對發芽蠶豆 GABA含量合 GAD活性的影響
發芽蠶豆GABA含量隨VB6濃度增加呈先升后降的趨勢(表4)。
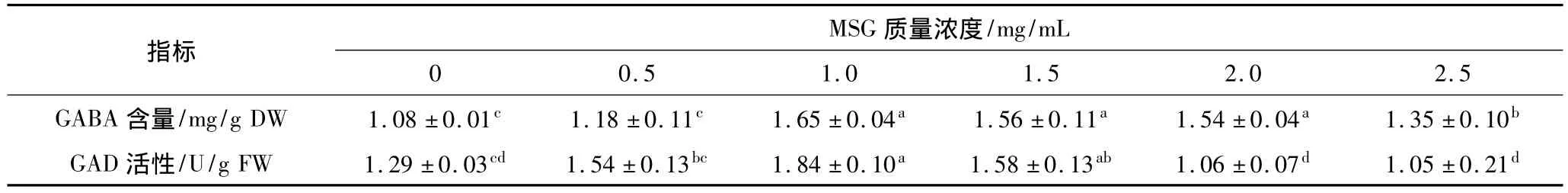
表2 MSG對發芽蠶豆GABA含量和GAD活性的影響

表3 CaCl2對發芽蠶豆GABA含量和GAD活性的影響

表4 VB6對發芽蠶豆GABA含量和GAD活性的影響
VB6濃度為60~120μmol/L時,GAD活性無顯著差異,而VB6濃度為60μmol/L時,GABA含量達到最大值,是對照的1.35倍。VB6是磷酸吡哆醛(PLP)的前體物質,而PLP是GAD的輔基,適量添加PLP可促進其與脫輔基蛋白結合而激活GAD,促進MSG脫羧生產GABA,從而增加GABA含量[11]。張磊等[11]研究發現,在米糠孵育液中添加 VB6,可使GABA含量得到提高,本研究結果與此一致。
2.4 低氧聯合鹽脅迫下發芽蠶豆GABA富集的培養液組分優化
2.4.1 模型分析
對MSG濃度(A)、CaCl2濃度(B)和 VB6濃度(C)進行了三因素三水平響應面分析試驗,試驗設計與結果見表5。利用Design Expert軟件對表5數據進行二次多元回歸擬合,得到GABA含量對編碼自變量A、B和C的二次多項回歸方程:
y=1.181 15+0.525 00A+0.011 592C+1.875 000 AC-0.000 450BC-0.291 41A2-0.028 222B2-0.000 076 6C2
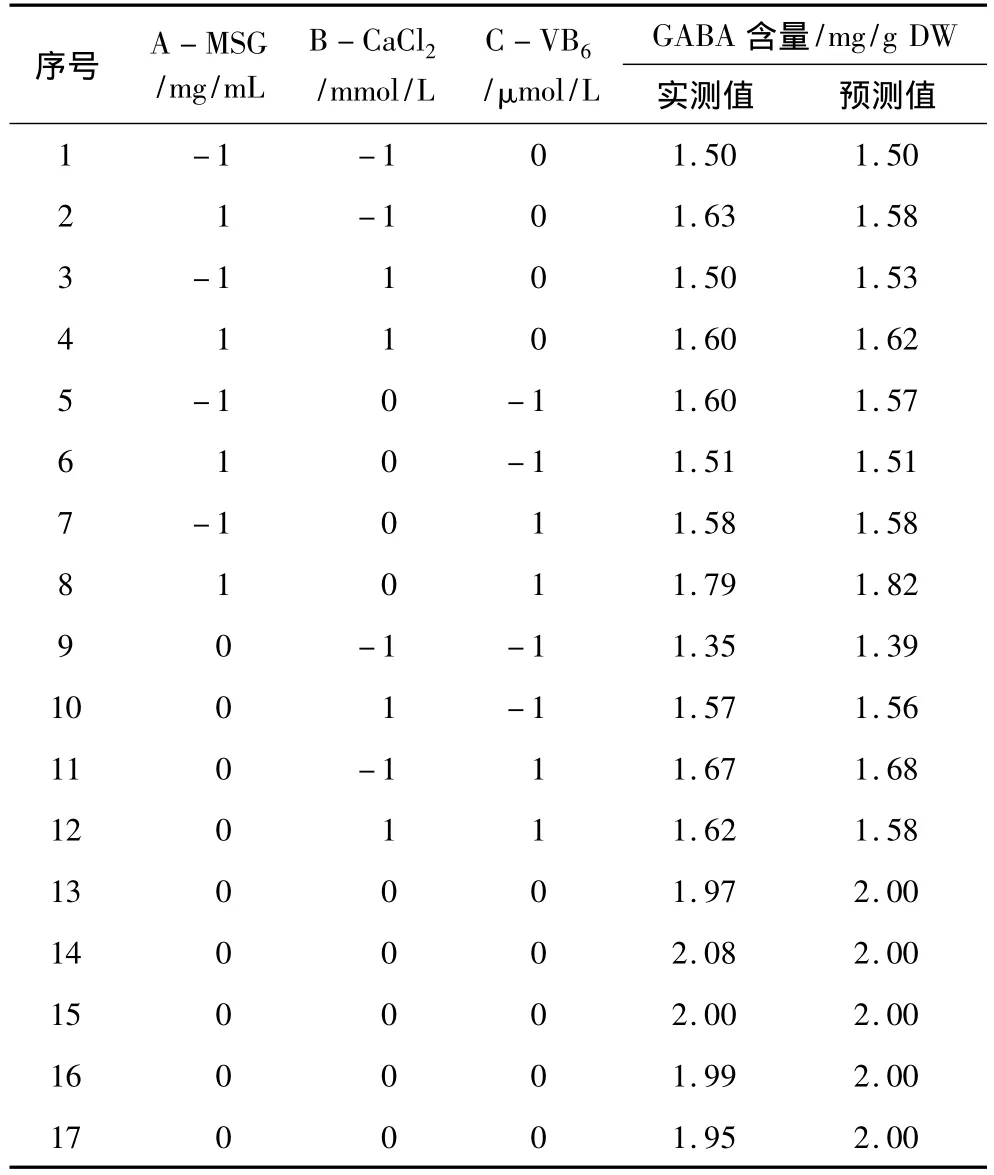
表5 中心組合試驗設計和結果
對上述回歸模型進行方差分析(表6)。結果表明,模型是顯著的(P<0.001),回歸模型的決定系數為0.976,說明該模型能夠解釋97.6%的變異。因此,可用此模型對GABA含量進行分析和預測。
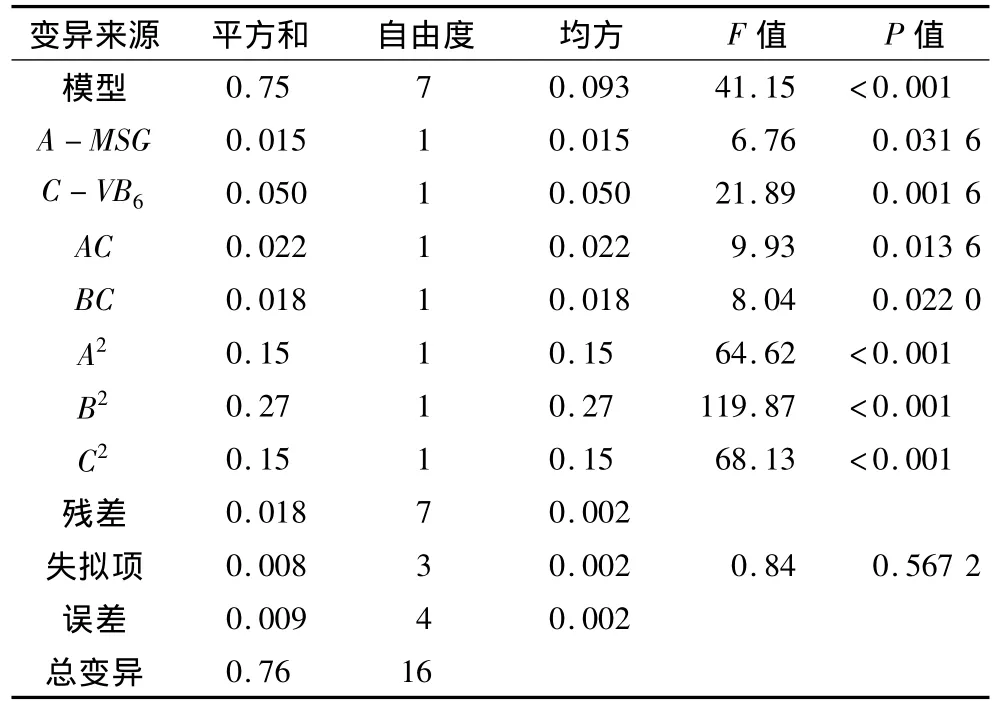
表6 回歸模型方差分析
2.4.2 發芽蠶豆富集GABA培養液組分優化
利用Design Expert軟件對表5數據進行二次多元回歸擬合,所得到的二次回歸方程響應曲面對應的等高線見圖1~圖2。
圖1顯示,CaCl2濃度為6 mmol/L時,MSG和VB6對發芽蠶豆GABA含量的交互作用顯著。因為等高線的形狀反映交互效應的強弱大小,橢圓形表示兩因素交互作用顯著[12]。VB6濃度一定時,GABA含量隨MSG濃度增加先升高后降低。VB6濃度較低(10~35μmol/L)時,圖1中等高線分布較密,表明此時MSG濃度對GABA含量的影響較大。當MSG濃度為1.1 mg/mL,VB6濃度為72 μmol/L 時,GABA含量達到最高值。MSG是GAD作用的底物,添加適量的MSG可以提高GAD活性,增加GABA含量。
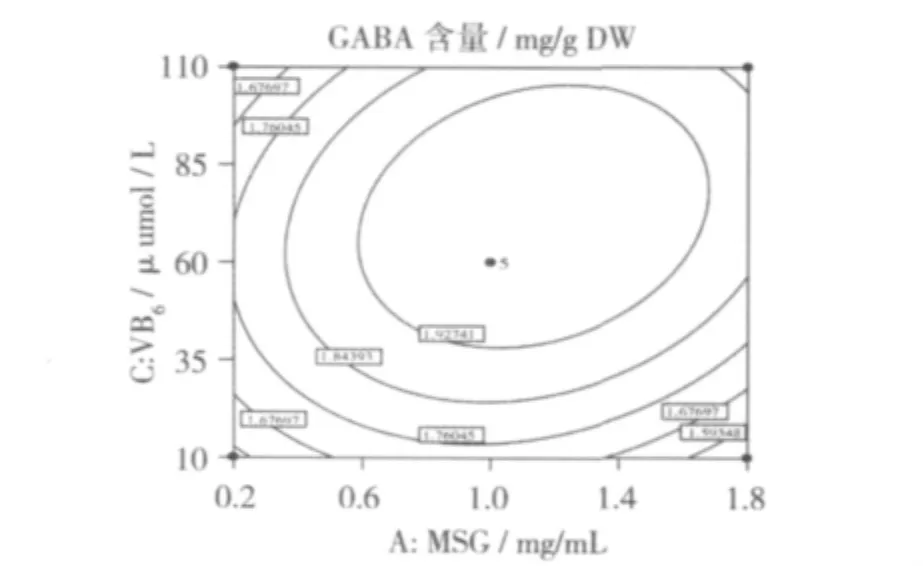
圖1 MSG和VB6的交互作用對GABA含量的影響
CaCl2和VB6對發芽蠶豆GABA含量的交互作用顯著(圖2)。與VB6相比,CaCl2濃度對GABA含量的影響較大。當CaCl2濃度一定時,GABA含量隨VB6濃度增加呈逐漸升高的趨勢。CaCl2為 6.1 mmol/L,VB6濃度為72μmol/L時,GABA含量達到峰值。由此可見,培養液添加適當濃度的CaCl2和VB6可提高發芽蠶豆中GABA含量。

圖2 CaCl2和VB6的交互作用對GABA含量的影響
Glu是GABA的前體物之一,而MSG是Glu的鹽存在形式,其在水溶液中解離后,滲入蠶豆種子,從而被發芽過程中所激活的GAD降解產生GABA;植物GAD氨基酸序列中C末端存在CaM結合區,該區域與Ca2+/CaM結合后可提高GAD活性,在足夠多的底物存在的情況下,從而提高GABA產率;PLP是GAD的輔酶,VB6與PLP結構相似,可起到輔酶功能,在蠶豆發芽過程中參與GAD的合成,提高GAD活力,促進Glu脫羧形成GABA,從而使GABA含量增加。
目前報道,GABA的形成有兩條途徑,即GABA支路和多胺氧化降解途徑[13]。其中GABA支路對發芽蠶豆GABA富集的貢獻約占70%(數據未展示)。在GABA支路中,GAD活力的提高和足夠的底物(MSG)對GABA的產生必不可少,而VB6是GAD輔酶的結構類似物,參與酶的合成來提高GAD活力。所以,MSG和VB6的加入對GABA的形成具有協同作用。作為第二信使,Ca2+通過信號傳遞的方式與VB6共同調節GAD活力,對GABA的產生具有協同作用。
2.4.3 驗證性試驗
對發芽蠶豆GABA含量的二次多項模型解逆矩陣后得出,在低氧聯合鹽脅迫培養條件下,培養液組分中MSG 質量濃度為 1.1 mmol/L、CaCl2濃度6.1 mmol/L、VB6濃度72μmol/L時,發芽蠶豆中GABA富集量的預測值為2.01 mg/g DW,實測值為(1.98±0.09)mg/g DW,是對照[(1.08 ±0.01)mg/g DW]的1.83倍,且高于隨機選擇組。相關性分析表明,實測值與預測值接近,表明實驗所擬合的模型可用來預測培養液組分和試驗響應值之間的關系(表7)。

表7 驗證試驗設計和結果
3 結論
低氧聯合鹽脅迫下,培養液中3種組分(MSG、CaCl2和VB6)對發芽蠶豆GAD活性和GABA含量的影響均達到顯著水平(P<0.005)。當MSG質量濃度為 1.1 mg/mL、CaCl2濃度為 6.1 mmol/L、VB6濃度為72μmol/L時,發芽蠶豆中GABA富集量是對照[(1.08 ±0.01)mg/g DW]的1.83 倍,達到(1.98 ±0.09)mg/g DW。
[1]Kmiecik W,Lisiewska Z,Jaworska G.Content of ash components in the fresh and preserved broad bean(Vicia faba v major)[J].Journal of Food Composition and Analysis,2000,13:905-914
[2]張暉,徐滿英.γ-氨基丁酸作用的研究進展[J].哈爾濱醫科大學學報,2006,40(3):267-268
[3]Shelp B J,Bown A W,Mclean M D.Metabolism and functions of gamma- aminobutyric acid[J].Trends in Plant Science,1999,411:446-452
[4]Fukao T,Bailey- Serres J.Plant responses to hypoxia- is survival a balancing act[J].Trends in Plant Science,2004,9:449-456
[5]Reddy V S,Reddy A S N.Proteomics of calcium- signaling components in plants[J].Phytochemistry,2004,65:1745-1776
[6]Komatsuzaki N,Tsukahara K,Toyoshima H,et al.Effect of soaking and gaseous treatment on GABA content in germinated brown rice[J].Journal of Food Engineering,2007,78(2):556-560
[7]Li Y,Bai Q Y,Jin X J,et al.Effects of cultivar and culture conditions onγ-aminobutyric acid accumulation in germinated fava beans(Vicia faba L.)[J].Journal of the Science of Food and Agriculture,2010,90:52-57
[8]Bai Q Y,Fan G J,Gu Z X,et al.Effects of culture conditions onγ-aminobutyric acid accumulation during germination of foxtail millet(Setaria italica L.)[J].European Food Research and Tech-nology,2008,228:169-175
[9]顧振新,蔣振輝.食品原料中γ-氨基丁酸(GABA)形成機理及富集技術[J].食品與發酵工業,2002,28(10):65-69
[10]王玉萍,韓永斌,顧振新,等.谷氨酸鈉和抗壞血酸對發芽糙米中GABA富積效果的影響[J].南京農業大學學報,2006,29(2):94-97
[11]張磊,白青云,曹曉虹,等.米糠富集γ-氨基丁酸的培養液組分優化[J].食品與發酵工業,2008,34(7):77-81
[12]范龔健,韓永斌,顧振新.用響應面法優化紅甘藍色素提取工藝參數[J].南京農業大學學報,2006,29(1):103 -107
[13]李巖.發芽蠶豆γ-氨基丁酸富集途徑及高效富集技術研究[D].南京:南京農業大學,2009.
Optimization of Culture Solution Compositions for γ-aminobutyric Acid Accumulation in Germinated Fava Beans(Vicia faba L.)
Chen Hui Yang Runqiang Han Yongbin Gu Zhenxin
(Key Laboratory of Agricultural and Animal Products Processing and Quality Control,Ministry of Agriculture,Nanjing Agricultural University,Nanjing 210095)
In this paper,the effects of MSG,CaCl2and VB6on glutamate decarboxylase(GAD)andγ-aminobutyric acid(GABA)of germinated fava beans were investigated.The culture solution compositions used for GABA accumulation in fava beans were optimized with Box-Behnken design.The results showed that under the condition of hypoxia combined with salt stress,the effects of MSG,CaCl2and VB6on GAD and GABA of germinated fava beans all reached significant levels(P <0.005).The quadric regression equation was established between GABA content and culture solution compositions.The R-squared reached 0.976,which meant that the equation could predict the changes of accumulation contents of GABA well.The optimum compositions of culture solution for GABA accumulation were MSG of 1.1 mg/mL,CaCl2of 6.1 mmol/L and VB6of 72 μmol/L.Under these conditions,the content of GABA was(1.98 ±0.09)mg/g DW,which was 1.83 times as much as that of the control[(1.08 ±0.01)mg/g DW].
germinated fava beans,γ - a minobutyric acid,accumulation,optimization
A
1003-0174(2011)11-0027-05
中央高校基本科研業務費專項資金項目(KYZ200 917)
2011-01-26
陳惠,女,1986年出生,碩士,食品中功能成分的富集技術
顧振新,男,1956年出生,教授,博士生導師,生物技術與功能食品
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
