Zn2+及Fe3+對嗜淀粉乳桿菌開放式發(fā)酵產(chǎn)乳酸的影響
2012-01-13 08:34:36汪群慧劉建國北京科技大學(xué)環(huán)境系北京100083
中國環(huán)境科學(xué) 2012年3期
關(guān)鍵詞:體系
鄒 惠,汪群慧,劉建國,王 爽(北京科技大學(xué)環(huán)境系,北京 100083)
乳酸是重要的生物化工產(chǎn)品,廣泛用于醫(yī)藥、食品、飲料、日用化工、化工、石油化工、皮革、卷煙工業(yè)等領(lǐng)域.其異構(gòu)體合成的L-乳酸聚合物,可生產(chǎn)能生物降解的農(nóng)用地膜及其他塑料制品,可有效解決全球的“白色污染”問題,因此,乳酸有廣闊的國內(nèi)外市場[1].
嗜淀粉乳桿菌是一種具有淀粉水解活性的乳酸細(xì)菌,它產(chǎn)生的解淀粉酶具有淀粉酶和支鏈淀粉酶雙重活性,可以直接轉(zhuǎn)化復(fù)雜的淀粉質(zhì)底物生成乳酸[2-4].餐廚垃圾是一種淀粉含量較高的有機(jī)廢物,其淀粉含量達(dá)到 41.38%~55.09%(按干重計(jì)),嗜淀粉乳桿菌的加入將有利于簡化糖化等生產(chǎn)工藝,故此類細(xì)菌在乳酸發(fā)酵工業(yè)上應(yīng)用廣泛[5-7].傳統(tǒng)乳酸發(fā)酵方式采用滅菌式的非開放式發(fā)酵,這種發(fā)酵會(huì)消耗大量電能,采用不滅菌的開放式發(fā)酵是一種理想的發(fā)酵方式[8].目前微生物發(fā)酵過程中存在著產(chǎn)酸率低,微生物對原料的利用率低,容易累積產(chǎn)生其它非目標(biāo)產(chǎn)物,生產(chǎn)周期長等問題.鋅離子、鐵離子等微量元素可影響細(xì)胞內(nèi)關(guān)鍵性酶活,當(dāng)?shù)孜镏衂n2+濃度大于1.748mg/L或Fe3+的濃度達(dá)到0.204mg/L時(shí),培養(yǎng)基中的乳酸菌生長良好,乳酸的積累量將大大增加[9-10].本課題組在前期實(shí)驗(yàn)基礎(chǔ)上,采用在開放式發(fā)酵過程中引入代謝調(diào)控物的方法,研究微量元素對乳酸發(fā)酵的影響,旨在突破產(chǎn)酸量低的瓶頸.
1 材料與方法
1.1 菌種
嗜淀粉乳桿菌(Lactobacillus amylophilus GV6),購買于中國普通微生物菌種保藏管理中心.
1.2 餐廚垃圾
取自北京科技大學(xué)鴻博園食堂,挑去骨頭、塑料袋等不可降解物后,垃圾中主要包括米飯、蔬菜、肉、蛋、豆腐、面條等.保存于-20℃的冰箱內(nèi)待用.其成分如表1所示(按干重計(jì))
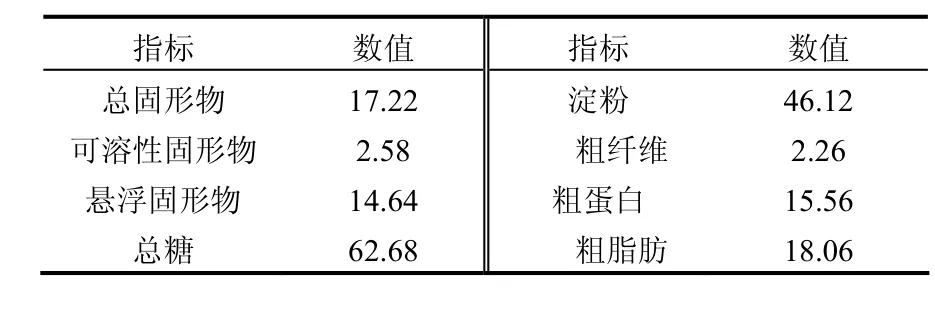
表1 餐廚垃圾成分表(%)Table 1 Composition of kitchen waste (%)
1.3 培養(yǎng)基
MRS培養(yǎng)基.121℃滅菌15min備用.
1.4 發(fā)酵方法
150g粉碎后的垃圾加入 75mL水,裝入500mL具塞玻璃瓶中(非開放式發(fā)酵在 121℃滅菌15min),通入高純氮?dú)馍蓞捬醐h(huán)境,按接種量2g(菌體濕重)/L(餐廚垃圾培養(yǎng)基)接入嗜淀粉乳桿菌(菌齡24h),37℃下發(fā)酵96h.
1.5 分析方法
取樣后 4000 r/min離心 15min,上清液經(jīng)0.45μm微孔濾膜過濾后測定.液相色譜選用日本島津公司高效液相色譜儀LC-20AT.
有機(jī)酸測定[11]為色譜柱:預(yù)柱 C185μm×26mm,分析柱 Inertsil ODS-SP 5um C186mm×250mm,檢測器UV 210nm,柱溫40℃,流動(dòng)相超純水(pH值為2.5~3.0),流速0.8mL/min.糖濃度測定[12]:分析柱為 Inertsil NH25um 250mm× 4.6mm,檢測器示差檢測器,柱溫 40℃,流動(dòng)相75%乙腈+25%超純水,流速1.0mL/min.采用峰面積外標(biāo)法定量,得到各種濃度.乙醇含量的測定:利用 SBA-40C型生物傳感器進(jìn)行測定.乙醇脫氫酶,乳酸脫氫酶測定[13]:乳酸脫氫酶是在催化丙酮酸轉(zhuǎn)化乳酸的同時(shí),將NADH氧化成NAD,使得NADH在340nm處的吸光度不斷降低,因此連續(xù)測定酶反應(yīng)過程中340nm處吸光度的變化即可算出酶活.酶活單位定義為25℃時(shí),每min氧化1umol的NADH所需的酶量;而乙醇脫氫酶是把丙酮酸轉(zhuǎn)為乙醇的關(guān)鍵性酶活,其大小是在340nm處檢測 NAD+的還原速度.酶活力單位定義為每min催化1umolNAD+還原需要的酶量.酶活力的計(jì)算公式為
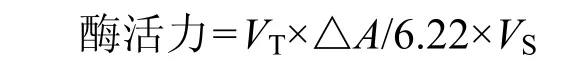
式中:VT為反應(yīng)總體積,mL;VS為樣品體積,mL;△A為每min吸光度的降低值.
將樣品置于冰浴中,200W 功率超聲波破碎20min,10000r/min離心 15min,除去細(xì)胞殘片,即得到粗酶液.每個(gè)樣品取5個(gè)平行.
2 結(jié)果與討論
2.1 利用嗜淀粉乳桿菌開放式發(fā)酵餐廚垃圾產(chǎn)乳酸的可行性
在發(fā)酵 96h后,2種發(fā)酵體系的發(fā)酵液色譜圖極其相似,說明他們的組成相差不大多,主要為乳酸、乙酸、甲酸及酒石酸.在開放式發(fā)酵下,乳酸產(chǎn)量最高值22.8g/L,占總有機(jī)酸的87.2%,而非開放式發(fā)酵下乳酸最大值僅為14.8g/L,占總有機(jī)酸的 71.5%,較開放式發(fā)酵乳酸產(chǎn)量低得多.推測在開放式發(fā)酵中,垃圾中存在各種代謝類型的微生物,使淀粉、纖維素等大分子得到不同程度的降解,提高了發(fā)酵液中乳酸細(xì)菌可利用的小分子化合物的含量,從而使乳酸產(chǎn)量增加[14],而且開放式發(fā)酵不需滅菌,節(jié)省了大量能源,故以后的實(shí)驗(yàn)均采用開放式發(fā)酵.
2.2 Zn2+及 Fe3+對開放式乳酸發(fā)酵中的相關(guān)酶活及代謝產(chǎn)物的影響
2.2.1 Zn2+及 Fe3+對發(fā)酵過程中乳酸產(chǎn)量的影響 在乳酸發(fā)酵過程中,乳酸菌經(jīng)過糖酵解途徑(EMP)利用糖后產(chǎn)生中間產(chǎn)物丙酮酸,此后丙酮酸的代謝決定了終產(chǎn)物的不同.為了獲得高產(chǎn)量的乳酸,最理想的是丙酮酸在乳酸脫氫酶(LDH)的作用下產(chǎn)生乳酸,如反應(yīng)式(1)所示,而在實(shí)際過程丙酮酸可能會(huì)在乙醇脫氫酶(ADH)的作用下被還原生成乙醇,如反應(yīng)式(2)式(3)所示,也有可能產(chǎn)生丙酸,丁酸等副產(chǎn)物,因此為提高乳酸產(chǎn)率,需在提高乳酸脫氫酶活性的同時(shí),控制副產(chǎn)物的生成,即降低乙醇脫氫酶等的活性[15].
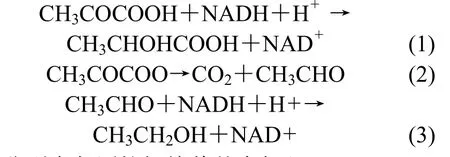
分別在餐廚垃圾培養(yǎng)基中加入0.02m mol/L ZnSO4×7H2O,0.01mmol/L Fe2(SO4)3,考察 Zn2+, Fe3+對嗜淀粉乳桿菌開放式乳酸發(fā)酵的影響由圖1可見, Fe3+的添加乳酸產(chǎn)量高達(dá)29.5g/L,較添加 Zn2+及空白樣(添加任何微量元素)的乳酸產(chǎn)量提高了 48.3%,29.4%,且添加 Fe3+的乳酸發(fā)酵L-乳酸純度為90.1%,而添加Zn2+及空白樣的L-乳酸純度分別為 82.7%,84.5%,這表明 Fe3+能促進(jìn)乳酸發(fā)酵,Zn2+卻相反.
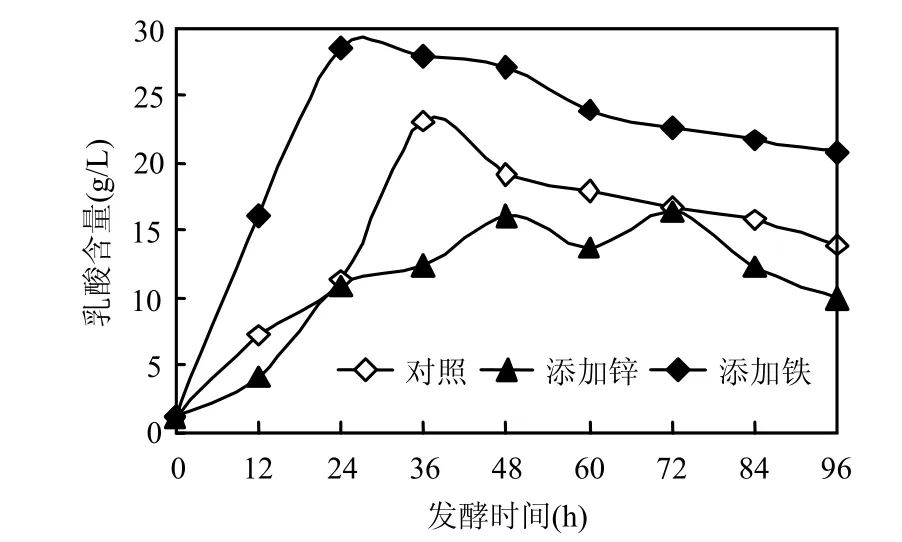
圖1 三種發(fā)酵體系中乳酸產(chǎn)量隨發(fā)酵時(shí)間的變化Fig.1 The influence on lactic acid under different microelement
2.2.2 Zn2+及 Fe3+對乙醇脫氫酶和乳酸脫氫酶酶活的影響 Zn2+,Fe3+是維持很多酶和轉(zhuǎn)錄因子功能的微量元素.它們對細(xì)胞的生長、發(fā)育和分化起至關(guān)重要的作用.鋅離子主要作為細(xì)胞蛋白、核酸、碳水化合物和脂類的共因子存在,通過調(diào)節(jié)鋅離子可影響細(xì)胞反應(yīng).
從圖2可知,添加Zn2+在整個(gè)發(fā)酵過程中抑制了乳酸脫氫酶的活性,卻提高了乙醇脫氫酶的活性,且在 48h時(shí),ADH活性達(dá)到最大值 10.75 U/mg;相反,添加 Fe3+卻抑制了乙醇脫氫酶的活性,提高了乳酸脫氫酶的活性,且在發(fā)酵進(jìn)行 48h后達(dá)到最高值11.87 U/mg,這間接說明在體系中添加 Fe3+能加快嗜淀粉乳桿菌積累乳酸.兩種離子的添加造成不同的結(jié)果,原因是Zn2+不是LDH的激活劑卻是ADH的激活劑,故其提高ADH的活性,降低了LDH的活性[16].
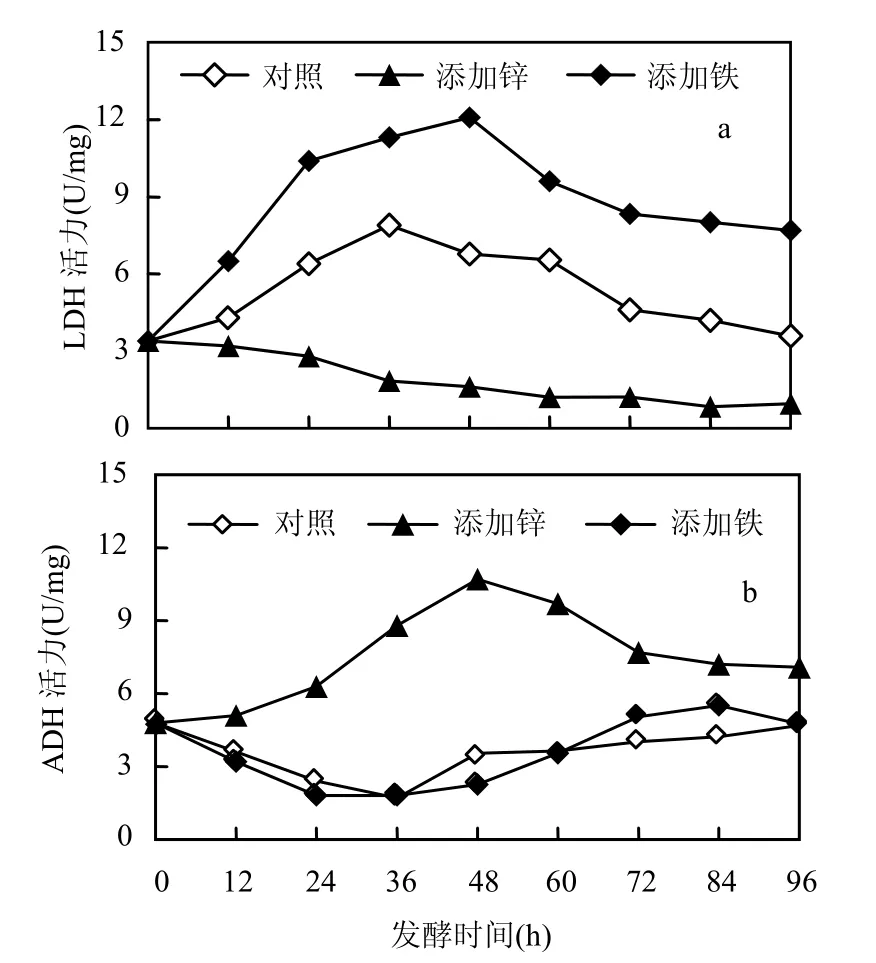
圖2 三種發(fā)酵體系中LDH和ADH隨發(fā)酵時(shí)間的變化Fig.2 The influence on LDH and ADH of microelement
加 Zn2+體系中乙醇脫氫酶(ADH)活性的提高,導(dǎo)致了乙醇的含量的增加(圖3和圖2b),而加Fe3+體系卻相反,它增加了乳酸脫氫酶(LDH)的活性而提高了乳酸的產(chǎn)量(圖1和圖2a).

圖3 不同微量元素條件下乙醇含量隨發(fā)酵時(shí)間的變化Fig.3 Variations of ethanol under different microelement
2.2.3 Zn2+及 Fe3+對發(fā)酵過程中 pH 值的影響 從圖 4可知,大量乳酸的產(chǎn)生使體系中氫離子濃度增高,進(jìn)而使得pH值逐漸降低,添加Fe3+的發(fā)酵體系pH值降得最低,約為3.39,這也能間接地反映其乳酸產(chǎn)量較其他2種條件下較高.當(dāng)發(fā)酵進(jìn)行 60h后,pH值基本不發(fā)生變化,與此結(jié)果相對應(yīng),圖2中LDH及加Zn2t體系的ADH也呈下降趨勢,說明過低的pH值抑制酶活性.
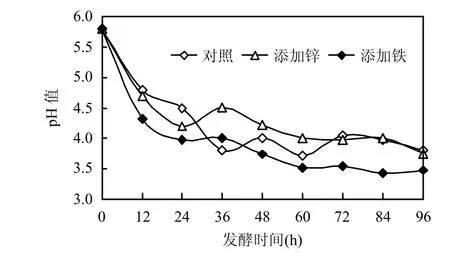
圖4 三種發(fā)酵體系中pH值隨發(fā)酵時(shí)間的變化Fig.4 Variations of pH under different microelement
2.3 Zn2+及 Fe3+對餐廚垃圾發(fā)酵過程中淀粉利用率的影響
從圖5可知,添加Fe3+的條件下,淀粉的利用率最高,發(fā)酵 84h后,淀粉含量由 46.12%降至17.34%,較添加Zn2+及未添加微量元素時(shí)利用率分別提高了 38.5%,28.1%.說明 Fe3+的加入的確能提高淀粉的利用率.
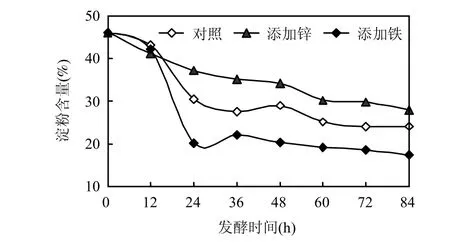
圖5 三種發(fā)酵體系中淀粉利用率隨發(fā)酵時(shí)間的變化Fig.5 Variations of starch under different microelement
2.4 嗜淀粉乳桿菌開放式發(fā)酵過程中糖組分的變化
添加 0.204mg/L Fe3+并接種嗜淀粉乳桿菌2g/L于餐廚垃圾中,在發(fā)酵37℃進(jìn)行開放式發(fā)酵,其發(fā)酵過程中各種糖組分濃度的變化如圖 6 所示.其中,葡萄糖,果糖,麥芽糖濃度先升高后降低,這可能是由于體系中存在的多糖降解成低糖,低糖最后被乳酸菌利用生成乳酸.發(fā)酵后期幾種糖濃度略有升高,這是乳酸積累到一定程度后其濃度對發(fā)酵進(jìn)行反饋抑制的結(jié)果[17],而在整個(gè)過程中乳糖的產(chǎn)量很少,且較難被乳酸菌利用.
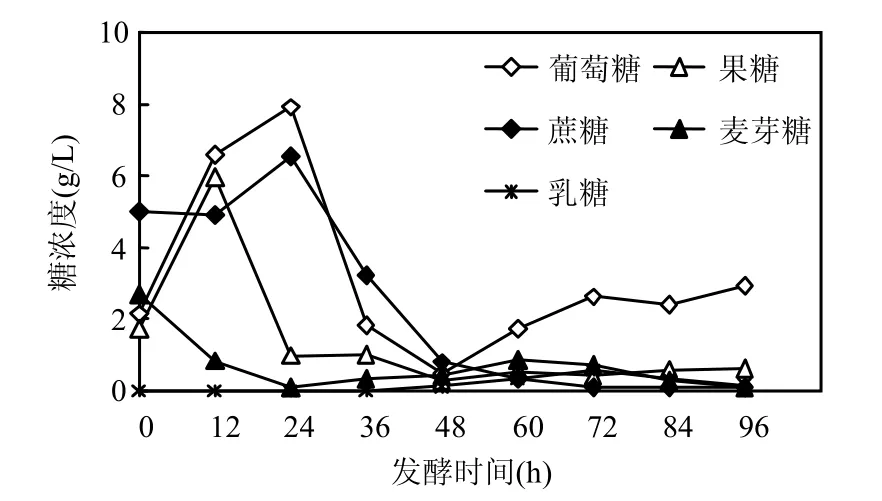
圖6 加Fe3+體系各種糖濃度隨發(fā)酵時(shí)間的變化Fig.6 Variations of sugar concentrations added with Fe3+
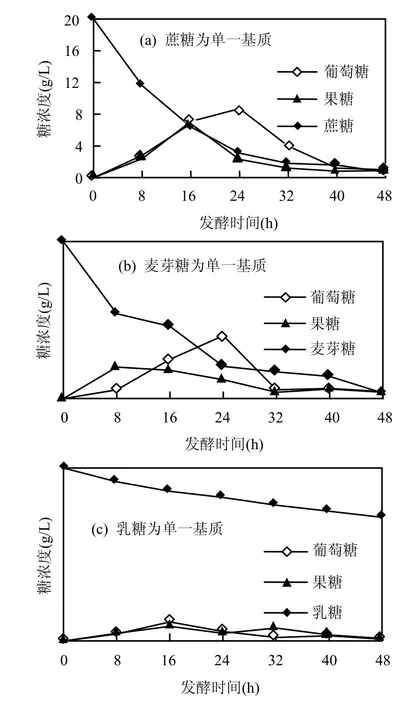
圖7 蔗糖、麥芽糖及乳糖的濃度隨發(fā)酵時(shí)間的變化Fig.7 Variations of sucrose, maltose and lactose
一般葡萄糖,果糖等單糖能直接被乳酸菌利用生產(chǎn)乳酸[2],為弄清楚二糖(蔗糖、麥芽糖及乳糖)的糖代謝途徑,將蔗糖,麥芽糖及乳糖作為單一基質(zhì)分別代替 MRS培養(yǎng)基中的葡萄糖,滅菌后在37℃下厭氧發(fā)酵48h,分析這三種糖被轉(zhuǎn)化利用的情況(圖7).圖7與圖6的結(jié)果類似,蔗糖,麥芽糖以及乳糖在發(fā)酵過程中會(huì)轉(zhuǎn)變成葡萄糖,果糖,最終被嗜淀粉乳桿菌利用,且麥芽糖和乳糖被降解的速度極大于乳糖.
3 結(jié)論
3.1 接種嗜淀粉乳桿菌的餐廚垃圾開放式發(fā)酵體系所產(chǎn)乳酸量高于非開放式發(fā)酵體系.這可能是由于大量土著菌的存在,促進(jìn)了餐廚垃圾中的高分子有機(jī)物水解成乳酸菌容易利用的低分子糖類物質(zhì).
3.2 Fe3+的加入能提高乳酸脫氫酶活性,抑制乙醇脫氫酶活性,使淀粉利用率和乳酸產(chǎn)量均高于對照體系,且能更好抑制其他有機(jī)酸的產(chǎn)生;而加Zn2+體系的乙醇脫氫酶(ADH)活性較高,導(dǎo)致副產(chǎn)物乙醇產(chǎn)量的增加,從而使淀粉利用率與乳酸產(chǎn)量低于對照體系.
3.3 在嗜淀粉乳酸菌的發(fā)酵體系中,二糖中的蔗糖和麥芽糖很容易降解成單糖(葡萄糖和果糖),從而被乳酸菌利用;而乳糖降解成單糖的速度緩慢.
[1] YUN Jong-jun,WEE Young-jung,RYU Hwa-won. Prodution of optically pure L(+)-lactic acid from various carbohydrates by batch fermentation of Enterococcus faecelis [J]. Enzyme and Microbial Technology, 2003,33:416-423.
[2] 趙國振,熊向峰,陳朝銀.解淀粉乳酸細(xì)菌在 L-乳酸發(fā)酵生產(chǎn)中的應(yīng)用 [J]. 中國生物工程雜志, 2009,29(7):134-139.
[3] Altaf M, Venkateshwar M, Srijana M, et al. An economicapp roach for L(+) 2lactic acid fermentation by Lactobacillus amylophilus GV6 using inexpensive carbon and nitrogen sources [J]. Appl. Microbiol., 2007,103:372-380.
[4] Thomsen M H, Guyot J P, Kiel P. Batch fermentations on synthetic mixed sugar and starch medium with amylolytic lactic acid bacteria [J]. Appl. Microbiol. Biotechnol., 2007,74:540-546.
[5] Muyanja C, Narvhus J A, Treimo J, et al. Isolation, characterization and identification of lactic acid bacteria from bushera: a Ugandan traditional fermented beverage [J]. Food Mierobio., 2002,80(2):201-210.
[6] Basa Janakiram Naveena, Mohammad Altaf,Kalva Bhadrayya, et al. Production of L(+) lactic acid by Lactobacillus amylophilus GV6 in semi-solid state fermentation using wheat bran [J]. Food Technol. Biotechnol., 2004,42(3):147-152.
[7] Naveena B J, Altaf M d, Bhadrayya K, et al. Direct fermentation of starch to L(+)-lactic acid in SSF by Lactobacillus amylophilus GV6 using wheat bran as support and substrate: medium optimization using RSM [J]. Process Biochemistry, 2005,40:681-690.
[8] 王旭明,汪群慧,馬鴻志,等.用乳酸細(xì)菌從有機(jī)廢棄物生產(chǎn)乳酸[J]. 現(xiàn)代化工, 2003,23(11):50-53.
[9] 潘麗軍,余 贇,鄭 志,等.米根霉乳酸脫氫酶的特性研究 [J].食品科學(xué), 2003,24(11):23-26.
[10] 封文濤,李十中,張晗星,等.微量元素在線調(diào)控 L-乳酸發(fā)酵的研究 [J]. 現(xiàn)代化工, 2009,29(2):144-146.
[11] LI Hua, HU Xian-ming, XIE Ying-feng. Using high performance liquid chromatographic method for determination of organic acids [J]. Chinese Journal of Chromatography, 1998,16(5):424-426.
[12] 王吉順,關(guān)家稅,王淑仁,等.高效液相色譜法測定葡萄酒中含糖量 [J]. 色譜, 1992,10(6):369-371.
[13] 鄭 志,姜紹通,羅水忠,等.限氧與通氧條件下米根霉ADH突變株的碳代謝特性 [J]. 中國農(nóng)業(yè)科學(xué), 2006,39(6):1228-1232.
[14] 王旭明,汪群慧.高產(chǎn)乳酸細(xì)菌發(fā)酵廚余垃圾生產(chǎn)乳酸的試驗(yàn)研究 [J]. 現(xiàn)代化工, 2005,25(Z1):148-150.
[15] Marcel H N Hoefnagel, Marjo J C Starrenburg, Dirk E Martens, et al. Metabolic engineering of lactic acid bacteria, the combined approach: kinetic modeling, metabolic control and experimental analysis [J]. Microbiology, 2002,148:1003-1013.
[16] Christopher D Skory. Isolation and expression of lactatedehydrogenase genes from Rhizopus oryzae [J]. Applied and Environmental Microbiology, 2000,66(6):2343-2348.
[17] 熊宗貴.發(fā)酵工藝原理 [M]. 北京:中國醫(yī)藥科技出版社, 2001: 69-70.
猜你喜歡
商品與質(zhì)量(2021年43期)2022-01-18 05:31:22
杭州(2020年23期)2021-01-11 00:54:42
新世紀(jì)智能(數(shù)學(xué)備考)(2020年11期)2021-01-04 00:38:16
中國外匯(2019年17期)2019-11-16 09:31:14
中國衛(wèi)生(2015年12期)2015-11-10 05:13:40
現(xiàn)代企業(yè)(2015年1期)2015-02-28 18:43:18
汽車零部件(2014年5期)2014-11-11 12:24:28
新高考·高一物理(2014年1期)2014-09-18 01:26:07
浙江人大(2014年1期)2014-03-20 16:19:53
終身教育研究(2012年4期)2012-03-25 10:41:11
