2010年中國東海夏季游離態2-苯基乙胺、腐胺、亞精胺和精胺的分布
2012-03-14 06:06:36李彩艷趙衛紅
海洋科學 2012年4期
李彩艷,趙衛紅,苗 輝
(1.中國科學院 海洋研究所 海洋生態與環境科學重點實驗室,山東 青島 266071;2.中國科學院 研究生院,北京 100049)
多胺是生物生長的調節因子,廣泛的存在于動物[1]、植物[2]、浮游生物[3-4]和海藻中[5]。內源性多胺可以與DNA、RNA和蛋白質結合,對生物的生長和發育起著重要作用[1,6-9]。外源性多胺可刺激植物細胞[10]和藻細胞的分裂增殖[11-12]。常見的多胺有腐胺、亞精胺,精胺和2-苯基乙胺。
多胺還被認為可以刺激和調控赤潮[11-13]。外源多胺可以刺激赤潮藻的生長,內源多胺對藻細胞的分裂增殖等有調節作用。腐胺和初級生產力存在正相關性也有報道,揭示了多胺在海洋氮循環中發揮著重要作用[14]。另外,多胺還可以加強藻毒素的毒性[15-16]。研究顯示多胺是赤潮發生的一個重要調控因子,但由于海水中多胺濃度較低以及測定方法的限制,我國還沒有海水中多胺濃度的報道。近年來,東海赤潮頻發,成為我國赤潮的高發區[17],東海赤潮的發生機制成為廣大海洋科學工作者研究的熱點。本文采用高效液相色譜法測定了2010年6月東海海水中的游離態多胺含量及分布,為探究多胺在東海赤潮中的作用提供科學依據和參數。
1 材料與方法
1.1 海水樣品的采集
于2010年6月8日到6月22日在長江口以及東海陸架(122.17°E~125.02°E,26.8°N~32.0°N)設置7個斷面 (圖1)。現場使用采水器分別采集表層、中層、底層水樣,表層為水面下約1 m,中層為葉綠素最大層,底層為距海底 0~3 m海水樣品經 GF/F膜(450℃灼燒4 h)過濾后儲存在棕色玻璃瓶中(450℃灼燒4 h),于-20℃冷凍保存水樣。
1.2 儀器和試劑
腐胺(putrescine,≥98%)、亞精胺(spermindine,≥98%)、精胺(spermine,≥97%)和丹磺酰氯(dansyl chloride,≥99%)均購自美國 Sigma公司;乙腈和丙酮為進口色譜純試劑(Honeywell,Burdick&Jackson公司);1,6-己二胺(1,6-diaminohexane)、碳酸鈉、氫氧化鈉、硼酸鈉、氨水、乙醚均為國產分析純;實驗用水為Milli-Q水(≥18.2MΩ)。采用Waters e2695高效液相色譜儀和Waters e2475熒光檢測器進行測定。
1.3 海水樣品游離態多胺的測定及分析方法
樣品測定前解凍,取 1 mL的海水樣品,加入70%的高氯酸,使其濃度達到 5%,4℃冰箱放置30 min。再加入 80 μL 2 mol/L 的 NaOH 溶液和 70 μL pH 9.18的硼酸,此時pH約為9,加入1mL丹磺酰氯(6 g/L丙酮)衍生劑,渦旋混勻30 s,40℃水浴避光反應45 min。加入40 μL 25%的濃氨水中止反應,靜置30 min。再加入60 μL乙腈,用0.22 μm有機針頭微孔濾膜過濾,最后采用高效液相色譜進樣分析[18]。
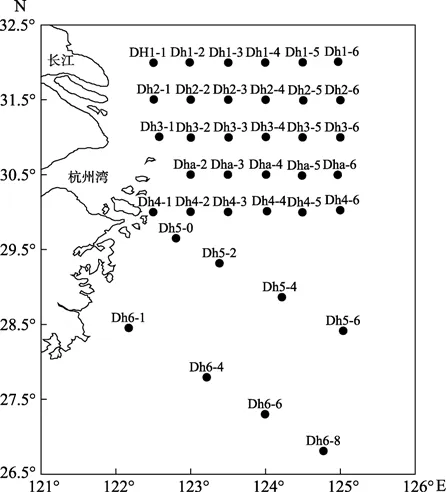
圖1 采樣站位圖Fig.1 Location of sampling stations
色譜柱為C18(150 mm×4.6 mm i.d.,5μm particle size,Agilent);熒光檢測激發波長(Ex)340 nm,發射波長(Em)515 nm;柱溫40℃;流動相A為乙腈,B為0.1 mol/L的醋酸銨(0.45 μm玻璃纖維濾膜過濾)。采用梯度洗脫,程序如下:0 min:A 35%,B 65%;0~10 min:A 35%~60%,B 65%~40%;10~15 min:A 60%~80%,B 40%~20%;15~20 min:A 80%~100%,B 20%~0;20~30 min:A 100%~35%,B 0~65%;30~35 min:A 35%,B 65%。衍生物進樣量為10 μL,流速1.0 mL/min。在該實驗條件下,2-苯基乙胺,腐胺、亞精胺、精胺得到了較好的分離,且三種胺工作曲線的相關系數都達到 0.999以上,檢測限分別為 9.3×10-11、9.6×10-11、2.8×10-10、1×10-10mol/L。
對測定后的樣品濃度采用sas9.1進行統計分析。
2 結果與分析
2.1 游離態多胺的濃度
由表1可以看出,主要的游離態多胺為腐胺和2-苯基乙胺,占總組成的比例為62%±15%和19%±11%,其次為精胺,所占比例分別為 14%±12%,亞精胺的含量最低,僅占總組成的比例的 6%±5%。不同的站位多胺的濃度變化較大。從各層總平均值看,除腐胺外,2-苯基乙胺、亞精胺和精胺在中層濃度最高。
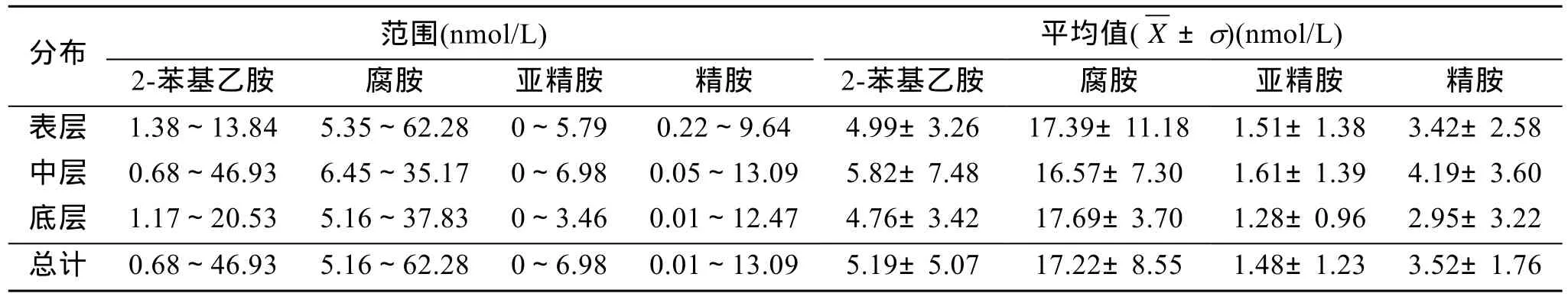
表1 海水中游離態多胺的濃度Tab.1 Concentrations of free polyamines in East China Sea
2.2 游離態多胺的大面分布
由圖2可以看出,2-苯基乙胺表層在長江口附近濃度較高,其次為杭州灣附近,各斷面濃度由高到低為 Dh1、Dh3、Dh4、Dha、Dh2、Dh5、Dh6;中層在杭州灣附近濃度較高,各斷面濃度由高到低為Dha、Dh3、Dh1、Dh4、Dh2、Dh6、Dh5;底層濃度較低,分布也較均勻,在調查區域的中東部出現高值,各斷面濃度由高到低為Dh4、Dha、Dh3、Dh1、Dh6、Dh5、Dh2。
由圖3可以看出,腐胺表層在長江口附近斷面的濃度較高,Dh2斷面濃度和其他斷面濃度差異達到顯著水平(P=0.0445),各斷面濃度由高到低依次為Dh2、Dha、Dh4、Dh5、Dh1、Dh3、Dh6;中層在長江口附近和杭州灣附近斷面濃度也較高,Dh2斷面濃度和其他斷面濃度差異達到顯著水(P=0.031),各斷面濃度由高到低依次為Dh2、Dha、Dh1、Dh6、Dh5、Dh4、Dh3;底層濃度最高,可能與生物尸體腐爛,蛋白質降解產生腐胺有關,各斷面濃度由高到低依次為 Dh2、Dha、Dh4、Dh3、Dh5、Dh1、Dh6。

圖2 2-苯基乙胺的大面分布(nmol/L)Fig.2 Distribution of free Pea (nmol/L)

圖3 腐胺的大面分布(nmol/L)Fig.3 Distribution of free Put( nmol/L)
亞精胺的濃度在四種胺中含量最低,表層在杭州灣附近南部和長江口附近斷面的中部形成高值區,各斷面濃度由高到低依次為Dh1、Dh3、Dh5、Dh2、Dh6、Dha、Dh4;中層和底層均在杭州灣附近區域出現高值區(圖4),中層各斷面濃度由高到低依次為Dha、Dh3、Dh1、Dh4、Dh2、Dh6、Dh5,底層各斷面濃度由高到低依次為Dh2、Dha、Dh4、Dh3、Dh5、Dh1、Dh6。

圖4 亞精胺的大面分布(nmol/L)Fig.4 Distribution of free Spd (nmol/L)
精胺表層在調查區域的西南部出現高值區,而長江口近岸附近濃度較低(圖5),各斷面濃度由高到低依次為 Dh6、Dh1、Dh5、Dh4、Dh3、Dh2、Dha;中層長江口附近濃度較低,而在調查區域的東部濃度較高,各斷面濃度由高到低依次為 Dh5、Dh1、Dh6、Dh3、Dh2、Dha、Dh4;底層分布和中層較為相似,在調查區域的東南部出現高值,各斷面濃度由高到低依次為 Dh5、Dh6、Dha、Dh3、Dh1、Dh4、Dh2。
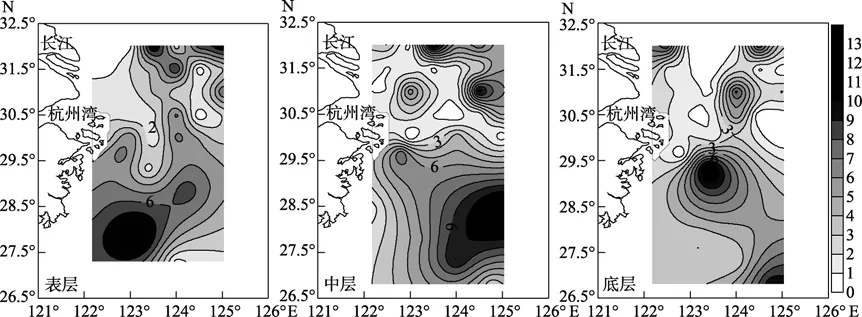
圖5 精胺的大面分布(nmol/L)Fig.5 Distribution of free Spm (nmol/L)
2.3 游離態多胺在各斷面的垂直分布
由表2可以看出,2-苯基乙胺在Dh2和Dh3斷面的垂直分布形式為中層>表層>底層;Dha和Dh6的分布形式為中層>底層>表層;Dh4和 Dh5的分布形式均為底層>中層;除Dh4和Dh5外,2-苯基乙胺在其他斷面的中層濃度均高于底層濃度。各斷面濃度的總體平均值由高到低依次為Dha、Dh3、Dh4、Dh1、Dh2、Dh6、Dh5。

表2 2-苯基乙胺在各斷面垂直分布Tab.2 The vertical distribution of Pea in 7 sections
由表3可以看出,腐胺在Dh3和Dha斷面的垂直分布形式為底層>中層>表層;Dh4和 Dh5的垂直分布形式為底層>表層>中層;Dh1、Dh2和Dh6的垂直分布形式為中層>底層。除Dh1和Dh2外,腐胺大多數斷面的底層濃度高于表層濃度。Dh2斷面的腐胺濃度平均值可達27.37 nmol/L±14.22 nmol/L,顯著高于其他斷面(p<0.001),各斷面濃度的總體平均值由高到低依次為 Dh2、Dha、Dh4、Dh1、Dh5、Dh3、Dh6。
由表4可以看出,亞精胺在Dh1、Dh3、Dh5斷面的垂直分布均為表層>中層>底層;Dh4和 Dh6的垂直分布均為底層>中層>表層;除 Dh4和 Dh6外,亞精胺大多數斷面的中層濃度高于底層。各斷面濃度的總體平均值由高到低依次為 Dha、Dh3、Dh1、Dh5、Dh6、Dh4、Dh2。
由表5可以看出,精胺在Dh1和Dha斷面的垂直分布為中層>表層>底層;Dh2、Dh4、Dh6斷面的垂直分布為表層>中層>底層;Dh3和 Dh5的垂直分布為中層>底層>表層;精胺在所有斷面的中層濃度都高于底層。各斷面濃度的總體平均值由高到低依次為 Dh5、Dh6、Dh1、Dh3、Dh4、Dh2、Dha。
3 討論
6月份海水中游離態腐胺的濃度最高,亞精胺的濃度最低。Nishibori發現日本瀨戶內海以亞精胺和腐胺為主,濃度分別為 0~4.4 nmol/L和 0~2.8 nmol/L[19]。Badini等[20]在研究亞得里亞海時發現海水中腐胺和亞精胺的濃度分別為 0~0.57 μmol/L和0~0.92 μmol/L,精胺為 0~0.02 μmol/L。2010 年4、5月份東海海水中主要的游離態多胺為精胺,由此可見,不同的海水樣品多胺的濃度差異較大,同一區域海水在不同季節多胺濃度的差異也較大。
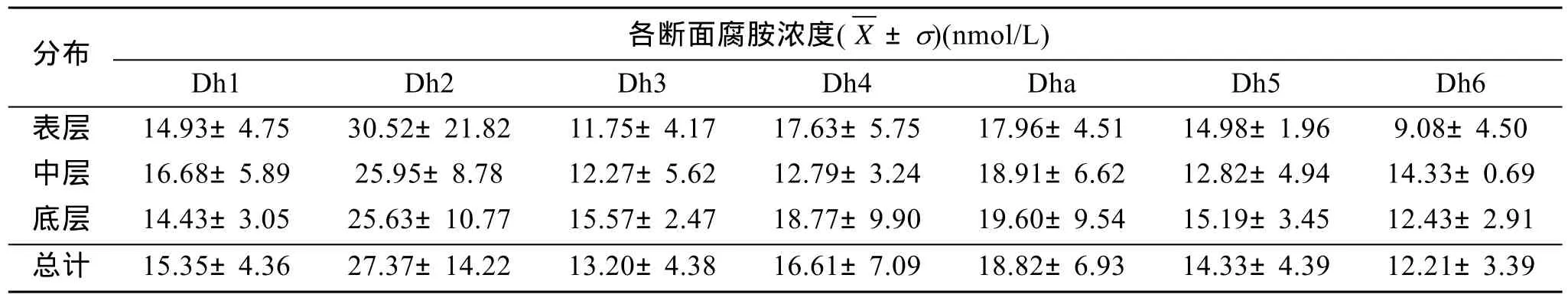
表3 腐胺在各斷面的垂直分布Tab.3 The vertical distribution of Put in 7 sections
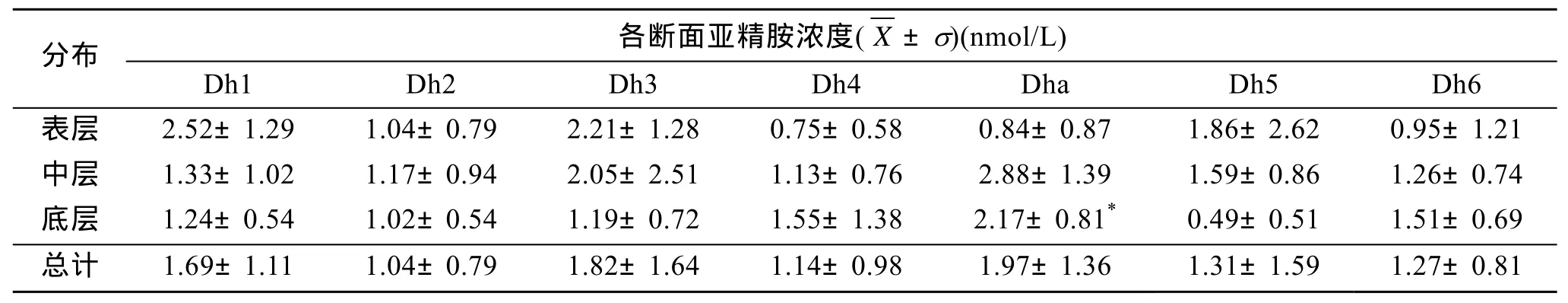
表4 亞精胺在各斷面的垂直分布Tab.4 The vertical distribution of Spd in 7 sections

表5 精胺在各斷面的垂直分布Tab.5 The vertical distribution of Spm in 7 sections
航次調查期間在 Dh1-3站位出現夜光藻赤潮,該站位亞精胺表層濃度達到 4.87 nmol/L,精胺表層和中層濃度分別為8.58 nmol/L 和11.98 nmol/L,遠高于總體平均值。對4、5月份東海赤潮高發區的調查顯示,2010年東海春季赤潮爆發區域的多胺的濃度大都較高,這與日本學者發現的瀨戶內海浮游植物大量繁殖區域多胺濃度較高一致[19]。
多胺的產生有三個條件,一是有可利用的自由氨基酸,二是有能分泌氨基酸脫羧酶的微生物存在;三是有適宜的環境條件,利于細菌的生長、脫羧酶的合成和提高脫羧酶活性[21]。氨基酸在適宜的條件下可直接轉化為腐胺,腐胺再進一步轉化生成亞精胺、精胺等。海洋環境中多胺的來源主要有以下四種途徑:①死魚及其他有機體的分解。在魚類及其他有機體死亡后,蛋白質可被微生物降解成游離態的氨基酸,經過脫羧而形成相應的多胺。②生物在代謝過程中也會產生多胺類物質,例如魚類、藻類等。③赤潮藻在消亡期會釋放大量多胺。④外源輸入。養殖廢水等富含腐敗蛋白質的廢水排入大海,能產生多種可被藻類吸收的生物活性物質,包括多胺。但多胺極不穩定,光度較強、溫度較高等,都會造成多胺的分解。另外,多胺可以被藻體等生物吸收、降解。因此海水中的多胺濃度會受到潮流、光照、溫度、營養物質、生物等多種因素的影響。
從實驗結果可以看出,2-苯基乙胺表層、腐胺和亞精胺表中底層均在長江口附近區域出現高值,這可能和長江沖淡水有關,一方面陸源物質中可能含有多胺以及多胺合成所需的氨基酸等,另一方面,該海區夏季適宜的光照和溫度,使浮游植物大量繁殖[22],生物體在代謝過程中會釋放多胺。2-苯基乙胺表中層、亞精胺表中底層及精胺的表層均在杭州灣附近區域出現高值,可能和該海區上升流的涌升帶來豐富的營養物質有關,該區域也比較適合浮游植物的生長。這兩個區域均為赤潮的高發區,由此可見,赤潮高發區夏季的多胺濃度也較高。
從總平均值看,2-苯基乙胺、亞精胺和精胺在中層的濃度最大,這可能是由于中層為葉綠素最大層,生物代謝旺盛,會像環境中釋放多胺。大多數斷面2-苯基乙胺、亞精胺和精胺的中層濃度均高于底層濃度可能也是這一緣故。腐胺在大多數斷面的底層濃度高于表層濃度,可能是海底沉積物中含有蛋白質等,蛋白質降解時產生多胺,從而導致多胺濃度較高。
4 結論
我國東海海水中 6月份腐胺的含量最高,亞精胺的含量最低,占總組成的比例分別為 62%±15%和6%±5%。從大面分布看,四種游離態多胺的高值大都出現在長江口及杭州灣附近區域。從各斷面的垂直分布看,2-苯基乙胺、亞精胺和精胺在大多數斷面的中層濃度均高于底層濃度。腐胺在大多數斷面的底層濃度高于表層濃度。夜光藻赤潮爆發處多胺的濃度相對較高。
[1]Vinci G,Antonelli M L.Biogenic amines:quality index of freshness in red and white meat[J].Food Control,2002,13(8):519-524.
[2]Minocha R,Long S.Simultaneous separation and quantitation of amino acids and polyamines of forest tree tissues and cell cultures within a single high-performance liquid chromatography run using dansyl derivatization[J].Journal of Chromatography A,2004,1035(1):63-73.
[3]Nishibori N,Nishijima T.Changes in polyamine levels during growth of a red-tide causing phytoplankton Chattonella antiqua (Raphidophyceae)[J].Eur J Phycol,2004,39(1):51-55.
[4]Nishibori N,Nishio S.Occurrence of polyamines in bloom forming toxic dinoflagellateAlexandrium tama-rense[J].Fisheries Science,1997,63:319-320.
[5]Nishibori N,Fujihara S,Nishijima L T.Changes in intracellular polyamine concentration during growth of Heterosigma akashiwo (Raphidophyceae)[J].Fisheries Science,2006,72(2):350-355.
[6]Theiss C,Bohley P,Voigt J.Regulation by polyamine of ornithine decarboxylase activity and cell division in the unicellular green alga Chlamydomonas reinhardtii[J].Plant Physiol,2002,128:1470-1479.
[7]Ober D,Gibas L,Witte L,et al.Evidence for general occurrence of homospermidine in plants and its supposed origin as by-product of deoxyhypusine synthase[J].Phytochemical,2003;62:339-344.
[8]Kotzabasis K,Senger H.Free,conjugated and bound polyamines during the cell cycle in synchronized cultures ofScenedesmus obliqus[J].Zeitschrift fur Naturforschung Teil C Biochemie Biophzsik Biologie Virologie,1994,49:181-185.
[9]Scoccianti V,Penna A,Penna N,et al.Effect of heat stress on polyamine content and proteinpattern inSkeletonema costatum[J].Marine Biology,1995,121:549-554.
[10]Smith T A.Polyamines[J].Annual Review of Plant Physiology,1985,36:117-143.
[11]Gentien P.Bloom dynamics and ecophysiology of theGymnodinium mikimotoispecics complex[J].Physiological Ecology of Harmful Algal Blooms,1998,41:155-173.
[12]Maestrini S Y,Balode M,Bechemin C,et al.Nitrogenous organic substances as potential nitrogen scources,for summer phyplankton in the Gulf of Riga,eastern Baltic Sea[J].Plankton Biol Ecol,1999,46:8-17.
[13]Iwasaki H.Growth physiology of red-tide microorganisms[J].Microbiol Sci,1984,1:179-182.
[14]Lee C,J?rgensen N O G.Seasonal cycling of putrescine and amino acids in relation to biological production in a stratifiedcoastal pond[J].Biogeochemistry,1995,29:131- 157.
[15]Legrand C,Johansson N,Johnsen G,et al.Phagotrophy and toxicity variation in the mixotrophicPrymnesium patelliferum[J].Limnol.Oceanogr,2001,46:1208-1214.
[16]Shilo M.Formation and mode of action of algal toxins[J].Bact Rev,1967,31:180-193.
[17]周名江,朱明遠.我國近海有害赤潮發生的生態學、海洋學機制及預測防治研究進展[J].地球科學進展,2006,21 (7):673-679.
[18]付敏,趙衛紅,苗輝,等.高效液相色譜法測定海水中游離態腐胺、亞精胺和精胺[J].分析化學,2010,38:1445-1449.
[19]Nishibori N,Matuyama Y,Uchida T.Spatial and temporal variations in free polyamine distributions in Uranouchi Inlet,Japan[J].Marine Chemistry,2003,82:307-314.
[20]Badini L,Pistocchi R,Bagni N.Polyamine transport in the seaweed Ulva Rigida (Chlorophyta)[J].Journal of Phycology,1994,30:599-605.
[21]Silla S M.Biogenic amines:their importance in foods[J].International Journal of Food Microbiology,1996,29:213-231.
[22]周名江,朱明遠,張經.中國赤潮的發生趨勢和研究進展[J].生命科學,2001,13(2):53-59.
