一株水解大豆蛋白細菌的鑒定及其酶學(xué)性質(zhì)研究
2012-04-13 12:30:32李娟娟
中國釀造 2012年12期
任 丹,陳 曦,李娟娟,馮 鎮(zhèn)*
(東北農(nóng)業(yè)大學(xué) 乳品科學(xué)教育部重點實驗室,黑龍江 哈爾濱 150030)
我國傳統(tǒng)發(fā)酵大豆食品資源豐富,主要有豆醬、豆豉、醬油和腐乳等[1]。許多大豆發(fā)酵食品多以傳統(tǒng)的方式進行生產(chǎn),因而存在生產(chǎn)周期長的問題。過長的成熟期會導(dǎo)致較高的生產(chǎn)成本。為了縮短傳統(tǒng)發(fā)酵食品的成熟周期,國內(nèi)外學(xué)者做了大量研究,主要途徑為提高成熟溫度、使用經(jīng)過修飾的發(fā)酵劑、添加輔助發(fā)酵劑或是直接添加蛋白酶,而在這些途徑中,使用蛋白酶來縮短發(fā)酵周期被認為是最簡單和行之有效的方法[2-5]。研究表明,對于具體的發(fā)酵食品而言,使用非固有發(fā)酵劑雖然可以達到促進成熟的目的,但往往改變了產(chǎn)品的特征質(zhì)構(gòu)與風(fēng)味[6]。對于外源酶的使用也存在相同的問題。因此,為了不改變傳統(tǒng)發(fā)酵食品的特征質(zhì)構(gòu)與風(fēng)味,研究者往往是從發(fā)酵食品中篩選具有良好發(fā)酵特性的固有發(fā)酵劑菌株或是利用來源于固有發(fā)酵劑菌株的酶,進行發(fā)酵食品的促熟研究。發(fā)酵食品特征風(fēng)味的形成是一個復(fù)雜生物化學(xué)反應(yīng)的綜合結(jié)果,包括由發(fā)酵微生物導(dǎo)致的糖酵解、脂肪水解和蛋白水解。對于發(fā)酵食品的風(fēng)味和質(zhì)構(gòu)特性而言,蛋白水解無疑是最重要的生化過程[7]。因此,發(fā)酵食品成熟的實質(zhì)之一是參與發(fā)酵的微生物所分泌蛋白酶作用于蛋白質(zhì)的結(jié)果。微生物所分泌的蛋白酶水解食品基質(zhì)中的蛋白質(zhì)產(chǎn)生游離氨基酸和肽,同時賦予發(fā)酵食品特征的風(fēng)味和質(zhì)構(gòu)。因此,研究參與食品發(fā)酵微生物所分泌蛋白酶的酶學(xué)性質(zhì)對于微生物及其蛋白酶的應(yīng)用至關(guān)重要。
在前期工作中,從我國傳統(tǒng)發(fā)酵腐乳中篩選出一株可以水解大豆蛋白的細菌,本研究是在此基礎(chǔ)上對該菌進行分子生物學(xué)鑒定。就酶制劑應(yīng)用于傳統(tǒng)大豆發(fā)酵食品而言,由于純酶制劑的價格較高,因此限制了其的應(yīng)用[8]。而粗酶的價格相對低廉更利于其推廣應(yīng)用。因此,在本研究中對該菌蛋白酶粗酶的酶學(xué)性質(zhì)進行了研究,目的是為通過酶法促進大豆發(fā)酵食品的成熟奠定基礎(chǔ)。
1 材料與方法
1.1 材料與試劑
菌株F1來源于東北農(nóng)業(yè)大學(xué)食品學(xué)院保藏。菌株的活化培養(yǎng)基為胰蛋白酶消化大豆肉湯(TSB)(Becton,Dickinson and Co.,Sparks,USA)培養(yǎng)基。菌株的產(chǎn)酶培養(yǎng)基為含6%(w/v)脫脂乳粉的胰蛋白酶消化大豆肉湯(TSB)培養(yǎng)基。其他生化試劑均為分析純。
1.2 儀器與設(shè)備
Bia-Rad電泳儀:美國Bia-Rad有限公司;T6型紫外可見分光光度計:北京普析通用儀器有限責(zé)任公司;TDL-40B型離心機:上海安亭科學(xué)儀器廠;DK-98-11A型電熱恒溫水浴鍋:天津市泰斯特儀器有限公司;ZWY-2102C/1102C型搖床培養(yǎng)箱:上海智城分析儀器制造有限公司;BX51-P型顯微鏡:奧林巴斯。
1.3 方法
1.3.1 形態(tài)結(jié)構(gòu)觀察及分子生物學(xué)鑒定
形態(tài)結(jié)構(gòu)觀察參照文獻[9]。將經(jīng)過活化的菌株F1接種于滅菌的TSB液體培養(yǎng)基中,30℃搖瓶發(fā)酵12h。試劑盒法提取細菌的總DNA,以總DNA為模板進行PCR反應(yīng),擴增細菌的16s rDNA序列,測定該序列并進行同源性分析(http://www.ncbi.nlm.nih.gov/BLAST/)。所用的引物為細菌通用引物[27f(5′-AGAGTTTGATC CTGGCTCAG-3′)和1512r(5′-ACGGCTACCTT GTTACGACT-3′)]。
1.3.2 粗酶液的制備及蛋白酶分子量的測定
將經(jīng)過活化的菌株F1接種于產(chǎn)酶培養(yǎng)基中進行培養(yǎng),30℃培養(yǎng)3d。4℃、12000r/min離心10min,除去菌體得粗酶液,4℃保存?zhèn)溆谩5鞍酌阜肿恿康臏y定通過聚丙烯酰胺凝膠電泳的方法[10]。采用不連續(xù)系統(tǒng),5%濃縮膠,12%分離膠,電泳緩沖液為Tris甘氨酸緩沖溶液(pH 8.0),加樣液與載樣液等量混合,煮沸2min~3min,考馬斯亮蘭染色。將粗酶液與上樣緩沖溶液混合,在進行電泳之前樣品不經(jīng)歷任何加熱處理。電泳后將凝膠浸沒于含有2.5%(v/v)Triton X-100的100mmol/L glycine-NaOH(pH 9.0)緩沖溶液中,緩慢搖動30min以除去SDS。用100mmol/L glycine-NaOH緩沖溶液(pH 9.0)清洗凝膠3次,以除去Triton X-100。在40℃的條件下,將凝膠在含有1%(w/v)酪蛋白的100mmol/L glycine-NaOH緩沖溶液(pH 9.0)中放置40min。最終用考馬斯亮藍R-250對凝膠進行染色。
1.3.3 溫度對蛋白酶活性和穩(wěn)定性的影響
溫度對蛋白酶活性的影響,在50mmol/L Tris-HCl的緩沖溶液(pH 7.0)中,分別在20℃、30℃、40℃、50℃、60℃和70℃時測定蛋白酶的活性,在進行試驗前將酶樣品和含有底物的50mmol/L Tris-HCl緩沖溶液(pH 7.0)培養(yǎng)到預(yù)定的溫度。對于蛋白酶熱穩(wěn)定性的研究,將酶樣品分別在20℃、30℃、40℃、50℃、60℃和70℃的條件下培養(yǎng)1h,然后在標準的條件下測定殘留的酶活力,未進行熱處理的酶樣品作為空白。
1.3.4 pH值對蛋白酶活性和穩(wěn)定性的影響
在pH 5.0~13.0的范圍研究pH值對蛋白酶活性的影響,應(yīng)用濃度為50mmol/L檸檬酸鈉(pH 5.0)、磷酸鉀(pH 6.0~8.0)、Tris-HCl(pH 7.0~9.0)、硼酸鈉(pH 9.0~10.0)和甘氨酸-氫氧化鈉(pH 11.0~13.0)的緩沖溶液。pH值對蛋白酶穩(wěn)定性的影響,將蛋白酶在上述不同pH值的緩沖溶液中于30℃的條件下培養(yǎng)1h,在標準的條件下測定殘留的蛋白水解活力。
1.3.5 NaCl和乙醇對蛋白酶活性和穩(wěn)定性的影響
在NaCl濃度為0~15%(w/v)和乙醇濃度為0~8%vol的條件下研究NaCl和乙醇對酶活力的影響,在標準的條件下測定酶活力。NaCl和乙醇對蛋白酶穩(wěn)定性的影響,酶在30℃的條件下于上述不同濃度的乙醇和NaCl溶液中培養(yǎng)1h。在標準的條件下測定殘留的酶活力。未經(jīng)NaCl和乙醇處理的酶樣品作為空白。
1.3.6 金屬離子對蛋白酶活性的影響
10mmol/L MgCl2、CaCl2、KCl、ZnCl2、FeSO4和MnCl2被用來研究金屬離子對蛋白酶活力的影響。在進行試驗之前,酶的樣品與金屬鹽在30℃培養(yǎng)60min。在標準的條件下測定殘留的酶活性。未經(jīng)金屬鹽處理的酶樣品作為空白。
1.3.7 蛋白酶活力的測定
蛋白酶活力的測定標準參照參考文獻[11],以酪蛋白為底物。
2 結(jié)果與分析
2.1 形態(tài)結(jié)構(gòu)觀察及分子生物學(xué)鑒定
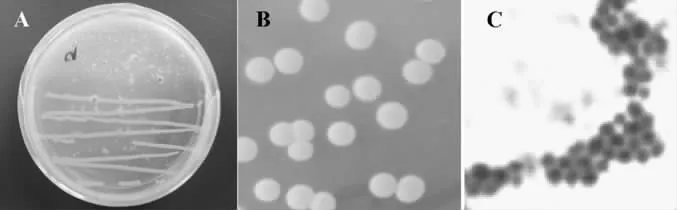
圖1 菌株F1水解大豆蛋白與菌落及菌體形態(tài)圖Fig.1 Hydrolyzed soy protein,colony and cell morphology of F1
從我國傳統(tǒng)的腐乳中篩選出一株對大豆蛋白有較強水解活性的細菌F1,在大豆蛋白瓊脂培養(yǎng)基中產(chǎn)生的水解現(xiàn)象見圖1A。菌株F1的菌落和菌體的形態(tài)學(xué)特征見圖1B和圖1C。菌株F1在TSB平板上菌落光滑、濕潤、橙黃色、邊緣整齊。采用革蘭氏染色顯微鏡下觀察,細胞為球形、無芽孢、過氧化氫酶陰性、革蘭氏陽性。菌株F1的16S rDNA測序結(jié)果見圖2,在Genebank中進行序列的同源性比對,與模式菌株Staphylococcus carnosusATCC 51365T(AB009934)的16S rDNA序列的同源性為99.7%,因此確定菌株F1為肉葡萄球菌(Staphylococcus carnosus)。肉葡萄球菌是凝固酶陰性球菌的一種常見菌株(Coagulase-negative cocci,CNC),這類細菌廣泛的存在于各種發(fā)酵食品中[12]。因CNC具有亞硝酸鹽和硝酸鹽還原酶活性,因此可以抑制脂質(zhì)氧化和食品的腐敗。CNC具有較強的蛋白水解作用,可以將食品中的蛋白質(zhì)水解為肽和游離氨基酸,而這些小分子物質(zhì)是許多發(fā)酵食品的重要風(fēng)味物質(zhì)或是風(fēng)味物質(zhì)的前體,同時賦予發(fā)酵食品特殊的質(zhì)構(gòu)和風(fēng)味[13]。因此這類細菌作為食品發(fā)酵劑而被廣泛應(yīng)用于發(fā)酵食品的生產(chǎn)。

圖2 菌株F1的16S rDNA序列Fig.2 16S rDNA sequence of strain F1
2.2 粗酶液的制備及蛋白酶分子量的測定
按實驗方法中的方法進行粗酶液的制備,以酪蛋白為底物在標準的條件下進行蛋白酶活力的測定,粗酶液的蛋白水解活力為83.87U/mL。SDS-PAGE 顯示該蛋白酶的分子量約為45ku。用酪蛋白作底物,在電泳圖中出現(xiàn)一條透明的條帶,說明該酶具有較強的蛋白酶活性。
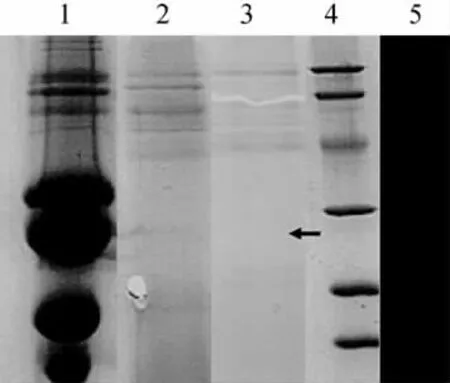
圖3 蛋白酶分子量的測定Fig.3 Electrophoretic analysis of protease molecular weight
2.3 溫度對蛋白酶活性和穩(wěn)定性的影響
溫度對蛋白酶催化活性和穩(wěn)定性影響結(jié)果見圖4。實驗結(jié)果表明,蛋白酶在30℃~50℃時(相對酶活力>50%)表現(xiàn)出較高的催化活性,蛋白酶的最佳催化溫度為40℃,此時的酶活力為92U/mL。就酶的穩(wěn)定性而言,當(dāng)酶在不同溫度經(jīng)歷1h的處理時,蛋白酶在20℃~50℃時表現(xiàn)出較高的酶活力(殘留蛋白酶活力>80%),而當(dāng)溫度為70℃時相對殘留酶活力僅為2%,因此該酶具有較低的耐熱性。多數(shù)發(fā)酵豆制品的發(fā)酵溫度為30℃,在本研究中的蛋白酶在30℃~50℃時顯示出較高的催化活力和穩(wěn)定性,因此,該蛋白酶適合于應(yīng)用通過提高發(fā)酵溫度或者是酶法生產(chǎn)發(fā)酵豆制品而促進發(fā)酵豆制品的成熟。此外,該酶具有較低的耐熱性,可以通過較高的熱處理溫度使其失活,從而防止其在應(yīng)用過程中造成過度成熟。

圖4 溫度對酶活力和穩(wěn)定性的影響Fig.4 Effect of temperature on the activity and stability of the protease
2.4 pH值對蛋白酶活性和穩(wěn)定性的影響
pH值對蛋白酶活力和穩(wěn)定性的影響見圖5。實驗結(jié)果表明,蛋白酶在pH 7.0~11.0(相對酶活力>50%)表現(xiàn)出較高的催化活力,最佳催化pH值為10.0,此時的蛋白酶活力為133U/mL。就pH值的穩(wěn)定性而言,當(dāng)酶在不同pH值條件下經(jīng)歷1h的處理時,在pH 7.0~11.0(殘留相對酶活力>58%),實驗結(jié)果表明,該蛋白酶為堿性蛋白酶,并且在較寬的pH范圍內(nèi)表現(xiàn)出較強的催化活性和穩(wěn)定性。本研究結(jié)果表明,該蛋白酶對pH值表現(xiàn)出的催化活性和穩(wěn)定性的特點對于許多發(fā)酵大豆食品是非常重要的,因為多數(shù)發(fā)酵大豆食品為堿性發(fā)酵食品,如kinema、Natto、Hawaijar、Thua nao、douche和sufu[14-16]。因此,就蛋白酶對pH值表現(xiàn)出的催化活性和穩(wěn)定性而言,該酶適合于應(yīng)用在大豆發(fā)酵食品中通過酶法或輔助手段進行促熟。
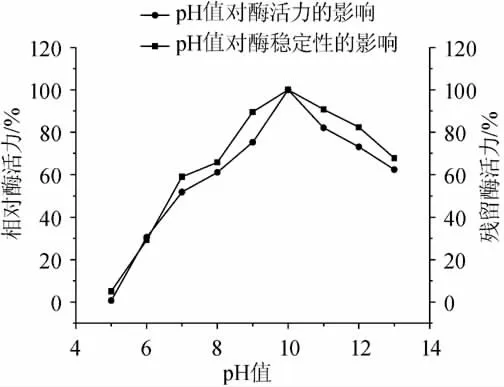
圖5 pH值對酶活力和穩(wěn)定性的影響Fig.5 Effect of pH value on the activity and stability of the protease
2.5 NaCl和乙醇對蛋白酶活力和穩(wěn)定性的影響
NaCl對蛋白酶穩(wěn)定性和活力的影響見圖6。實驗結(jié)果表明,當(dāng)NaCl的濃度為0~1%時,蛋白酶的活力明顯增加。當(dāng)NaCl的濃度大于1%時,隨著NaCl濃度的不斷增加酶活力明顯下降。當(dāng)NaCl的濃度為9%時,蛋白酶的相對活力為66%,其蛋白酶的催化活力為33U/mL。就酶的穩(wěn)定性而言,當(dāng)NaCl的濃度小于9%時,蛋白酶相對殘留活力大于48%。乙醇對蛋白酶穩(wěn)定性和活力的影響見圖7,實驗結(jié)果表明,該蛋白酶對乙醇表現(xiàn)出較高的活力和穩(wěn)定性,當(dāng)乙醇的濃度為8%vol時,蛋白酶的相對活力為76%。就酶的穩(wěn)定性而言,當(dāng)乙醇的濃度為8%vol時,其蛋白酶相對殘留活力仍為70%。研究結(jié)果表明,該酶對乙醇具有較強的耐受性。
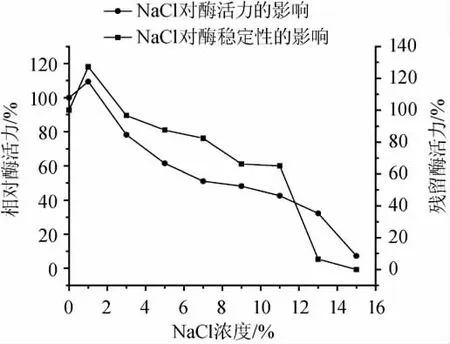
圖6 NaCl對酶活力和穩(wěn)定性的影響Fig.6 Effect of NaCl on the activity and stability of the protease
在發(fā)酵食品的成熟過程中,微生物所分泌的蛋白酶水解發(fā)酵基質(zhì)中的蛋白質(zhì)產(chǎn)生肽和氨基酸,但由微生物產(chǎn)生的蛋白酶可能會被一些鹽或者是添加劑所抑制,因此研究酶對具體的添加劑的耐受性是十分必要的[17]。該蛋白酶對NaCl和乙醇表現(xiàn)出較強的耐受性,這是非常重要的特性對于在發(fā)酵豆制品中的應(yīng)用,如sufu、miso、koji和soy sauce等,為了獲得較好的風(fēng)味和抑制一些治病菌的生長和繁殖,這些大豆發(fā)酵食品中往往在配料中加入較高濃度的NaCl和乙醇[18]。

圖7 乙醇對酶活力和穩(wěn)定性的影響Fig.7 Effect of ethanol on the activity and stability of the protease
2.6 金屬離子對蛋白酶活性的影響
幾種金屬離子對蛋白酶活性的影響見附表。在實驗考查的6種金屬離子中Mg2+、Ca2+、K+、Fe2+和Zn2+增強了蛋白酶的催化活性,而Mn2+減弱了蛋白酶的催化活性。許多金屬離子可以影響蛋白酶的催化活性,超過75%的酶需要金屬離子作為激活劑才能表現(xiàn)出催化活力[19]。本實驗的研究結(jié)果為該蛋白酶在大豆發(fā)酵食品工業(yè)中的應(yīng)用過程中通過金屬離子來調(diào)節(jié)酶的催化活力奠定了基礎(chǔ)。

附表 金屬離子對蛋白酶活力的影響Attached table Effects of metal ions on the activity of the protease
3 結(jié)論
本實驗對篩選出的一株對大豆蛋白具有良好水解能力的細菌進行了鑒定并對其粗酶的酶學(xué)性質(zhì)進行了研究。16S rDNA測序結(jié)果表明該菌與模式菌株Staphylococcus carnosusATCC 51365T(AB009934)的16S rDNA序列的同源性為99.7%,因此確定菌株F1為肉葡萄球菌。電泳法測得菌株F1產(chǎn)蛋白酶的分子量約為45ku。菌株F1產(chǎn)蛋白酶的最適反應(yīng)溫度和pH值分別為40℃和10.0,在20℃~50℃和pH 7.0~11.0 范圍內(nèi)較穩(wěn)定。Ca2+、Mg2+、K+、Fe2+和Zn2+對粗酶的催化活性有增強作用。粗酶對較高濃度的NaCl和乙醇表現(xiàn)出較好的穩(wěn)定性。
[1]DAE YK,JAMES WDIII,HYUN JK,et al.Antidiabetic effects of fermented soybean products on type 2 diabetes[J].Nutr Res,2010,30(1):1-13.
[2]CATHERINE T,AGNIESZKA K,SANDRA H,et al.Glutamate dehydrogenase activity:a major criterion for the selection of flavour-producing lactic acid bacteria strains[J].Antonie Van Leeuwenhoek,2002,82(4):271-278.
[3]ANJANI K,KAILASAPATHY K,PHILLIPS M.Microencapsulation of enzymes for potential application in acceleration of cheese ripening[J].Int Dairy J,2007,17(1):79-86.
[4]BRONCANO JM,TIMON ML,PARRA V,et al.Use of proteases to improve oxidative stability of fermented sausages by increasing low molecular weight compounds with antioxidant activity[J].Food Res Int,2011,44(9):2655-2659.
[5]KILCAWLEY KN,NONGONIERMA AB,HANNON JA,et al.Evaluation of commercial enzyme systems to accelerate Cheddar cheese ripening[J].Int Dairy J,2012,26(9):50-57.
[6]MIRNA MF,MARION E,ANDREA S,et al.Bacterial communities associated with the production of artisanal Istrian cheese[J].Int J Food Microbiol,2010,142(1):19-24.
[7]RANDAZZO CL,PITINO I,RIBBERA A,et al.Pecorino Crotonese cheese:Study of bacterial population and flavour compounds[J].Food Microbiol,2010,27(3):363-374.
[8]趙德安.酶制劑應(yīng)用于傳統(tǒng)大豆發(fā)酵食品釀造的探討[J].中國釀造,2007(4):58-61.
[9]凌代文,東秀珠.乳酸細菌分類鑒定及實驗方法[M].北京:中國輕工業(yè)出版社,19991.
[10]薩姆布魯克丁,弗里齊EF,曼尼阿蒂斯T,金冬雁譯.分子克隆實驗指南[M].北京:高等教育出版社,1993.
[11]KEMBHAVI AA,KULKARNI A,PANT A.Salt-tolerant and thermostable alkaline protease fromBacillus subtilisNCIM No.64[J].Appl Biochem Biotech,1993,38(1):83-92.
[12]LING G,KYEUNG HC,JONG-HOON L.Analysis of the cultivable bacterial community in jeotgal,a Korean salted and fermented seafood,and identification of its dominant bacteria[J].Food Microbiol,2011,28(1):101-113.
[13]KALLIOPI R,LUCA C.New developments in the study of the microbiota of naturally fermented sausages as determined by molecular methods:A review.Int J Food Microbiol,2006,108(2):255-267.
[14]NOUT MJR,BAKSHI D,SARKAR PK.Microbiological safety of kinema,a fermented soya bean food[J].Food Control,1998,9(6):357-362.
[15]JEYARAM K,MOHENDRO SINGH W,PREMARANI T,et al.Molecular identification of dominant microflora associated with‘Hawaijar’-A traditional fermented soybean(Glycine max(L.))food of Manipur,India[J].Int J Food Microbiol,2008,122(3):259-268.
[16]KATEKAN D,ARUNEE A,EKACHAI C,et al.Free-amino acid profiles ofthua nao,a Thai fermented soybean [J].Food Chem,2011,125(2):342-347.
[17]RUIZ-RAMIREZ J,ARNAU J,SERRA X,et al.Effect of pH,NaCl content and proteolysis index on the relationship between water content and texture parameters inbiceps femorisandsemimembranosusmuscles in dry-cured ham[J].Meat Sci,2006,72(2):185-194.
[18]BREWER MS.Encyclopedia of food microbiology[M].Urbana:Academic Press,2000.
[19]TUNGA R,BANERJEE R,BHATTACHARYA BC.Some studies on optimization of extraction process for protease production in SSF[J].Bioprocess Eng,1999,20(6):485-489.
