赤眼鱒線粒體D-loop和Cyt b基因序列的對比分析*
2012-05-10 06:44:48楊慧榮趙會宏蒙子寧林權卓
中山大學學報(自然科學版)(中英文) 2012年5期
關鍵詞:分析
楊慧榮,趙會宏 ,蒙子寧 ,劉 麗 ,林權卓
(1.華南農業大學動物科學學院,廣東 廣州 510642;2.中山大學水生經濟動物研究所暨廣東省水生經濟動物良種繁育重點實驗室,廣東 廣州 510275)
赤眼鱒Squaliobarbuscurriculus屬鯉科Cyprinidae,雅羅魚亞科Leuciscinae,赤眼鱒屬Squaliobarbus,俗稱紅眼棒、野草魚、參魚,是廣布中國主要水系的野生經濟魚類[1]。赤眼鱒因其肉質細嫩,味道鮮美,具有抗病力強,食性雜,生長較快等特點,成為我國重要的經濟魚類之一。目前,有關赤眼鱒的研究主要集中于資源狀況、基礎生物學、養殖和人工繁殖方面[2-3],而關于其群體遺傳學與基因多樣性研究則鮮有報道,僅2008年楊太有等[4-6]用RAPD和ISSR技術對丹江口水庫、青龍湖和宿鴨湖3地理群體進行了遺傳多樣性分析。
線粒體DNA(Mitochondrial DNA,mtDNA)具有母性遺傳、進化速度快、核苷酸替代率高等特點,已成為魚類進化生物學和群體遺傳學研究的重要遺傳標記[7-8]。mtDNA基因組內不同的區域進化速度存在差異,適合不同水平的進化研究。在mtDNA上,位于tRNAPro和tRNAPhe基因之間的D控制區(D-loop)是整個mtDNA上序列和長度變異最大的區域[9],也線粒體基因中進化最快的部分,適用于種群水平差異的檢測,也可用于種間分析[7-10]。而Cytb基因是線粒體DNA上的蛋白質編碼基因,其進化速度適中,適合種群水平差異的檢測,是探討種間和種內遺傳分化程度的良好標記[11-13]。本研究開展了珠江水系和長江水系不同水域赤眼鱒mtDNA D-loop和Cytb基因序列的對比分析,旨在研究這兩種遺傳標記對赤眼鱒遺傳多樣性狀況和群體遺傳結構的差異,探討mtDNA D-loop和Cytb基因對于群體遺傳多樣性分析的優缺點,進而為針對不同研究材料、不同研究目的,選擇不同的分析方法提供技術上的指導,使得結論更客觀真實。
1 材料與方法
1.1 材料
赤眼鱒樣品于2005年6月分別采自長江水系的宜昌和武漢江段(編號為YC群體、WH群體),珠江水系梧州和新豐江段(編號為WZ群體、XF群體)。YC群體取自宜昌巴東縣的官渡口,屬于長江上游干流巫峽的起點;WH群體取自武漢市境內江段,屬于長江中游水域;WZ群體和XF群體分布代表了珠江水系的西江和東江流域。共計24個個體,均為野生群體。活體運回實驗室后取背部肌肉并置于超低溫冰箱,取樣后的赤眼鱒用福爾馬林固定保存。
1.2 方法
基因組DNA的提取參考《分子克隆實驗指南》[14]的方法略加改進。PCR反應在Biometra PCR儀上進行。D-loop和Cytb引物由Invitrogen生物技術有限公司合成,D-loop引物序列如下:DL1:5'-ACCCCTGGCTCCCAAAGC-3',DH2:5'-ATCTTAGCATCTTCAGTG-3'[15]。Cytb引物序列如下:L14724:5'-GACTTGAAAAACCACCGTTG-3',H15915:5'-CTCCGATCTCCGGATTACAAGAC-3'[12]。反應總體積50 μL,其中10×Taq Buffer 5 μL,Taq Polymerase 0.5 μL (5 U·μL-1),dNTPs(2.5 mmol·L-1)4 μL,Mg2+(25 mmol·L-1)2.5 μL,引物1和2(10 umol·L-1)各2 μL,模板DNA 50 ng,無離子超純水35.5 μL。擴增條件為:94 ℃預變性2 min后,再進行35個循環,每一循環包括:94 ℃ 45 s,58 ℃ 45 s,72 ℃ 1 min;最后72 ℃延伸7 min。每次PCR反應均設不含模板DNA的空白對照。擴增產物經w=1.5%TBE瓊脂糖凝膠電泳分離,EB染色,紫外燈光下檢測、拍照。
1.3 測序與數據分析
產物純化和序列測定由上海基康公司(Gene core Co.)完成,通過雙向測序獲得赤眼鱒D-loop和Cytb序列,經SeqManII (DNASTAR Inc)軟件拼接并賦予人工校對后,用CLUSTAL-X程序對所獲得的序列進行比對[16],采用軟件DAMBE統計單倍型并作圖,檢測轉換、顛換是否達飽和。群體的遺傳多樣性指標單倍型多樣性(Haplotype diversity,H)和核苷酸多樣性(Nucleotide diversity,π)由ARLEQUIN[17]統計軟件計算獲得,群體間的分化指數(F-statistics,FST)[18]的計算在ARLEQUIN中完成,群體間的基因流[19]由公式Nm=[(1/FST)-1]/2計算得出,群體遺傳變異的分子方差分析(AMOVA)[20]在ARLEQUIN中完成。采用MEGA4.1軟件[21]統計DNA序列的堿基組成,轉換/顛換、插入/缺失位點數,單突變位點,簡約信息位點和平均轉換/顛換率;利用Kimura雙參數模型計算個體間、群體內和群體間的遺傳變異率;用NJ法進行聚類分析,構建聚類關系樹。
2 結 果
2.1 D-loop和Cyt b基因序列信息
由軟件DAMBE統計可知:在24個598 bp的D-loop基因序列中共檢測出18個單倍型。由MEGA4.1軟件對D-loop基因序列進行分析,檢測到18個單突變位點,34個簡約信息位點數;在67個突變位點中:轉換位點40個,顛換位點15個(其中有2個位點即發生了轉換,又發生了顛換),插入/缺失位點14個,轉換明顯高于顛換,平均的轉換發生頻率是顛換的3.192。24個個體D-loop基因片段中,T、C、A、G堿基的平均含量分別為35.1%,17.5%,33.7%,13.7%,其中,A+T的含量(68.8%)明顯高于C+G的含量(31.2%)。同樣,在24個752 bp的Cytb基因序列中共檢測出18個單倍型,檢測到4個單突變位點,68個簡約信息位點數;在72個突變位點中:轉換位點60個,顛換位點15個(其中有3個位點即發生了轉換,又發生了顛換),無插入/缺失位點,轉換明顯高于顛換,平均的轉換發生頻率是顛換的4.636。T、C、A、G堿基平均含量分別為26.4%,29.8%,29.3%,14.5%,其中,A+T的含量(55.7%)略高于C+G的含量(44.3%)。對比分析可知:Cytb除簡約信息位點百分比大于D-loop外,多態位點百分比和單突變位點百分比均小于D-loop(表1)。
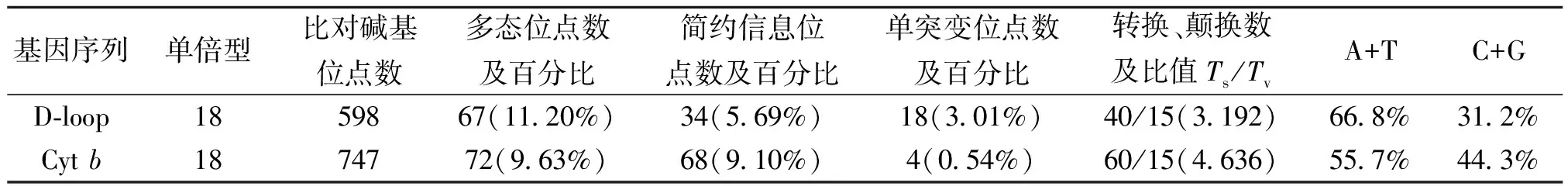
表1 赤眼鱒線粒體D-loop 及Cyt b堿基組成及變異分析
2.2 D-loop和Cyt b基因序列的遺傳變異分析
用軟件MEGA4.1統計出兩水系4個不同水域赤眼鱒各個體間的轉換/顛換數及遺傳變異率。D-loop序列中,各個體間的轉換位點為0~24,顛換位點為0~8,均為轉換大于顛換。兩大水系4個不同群體個體間的遺傳變異率在0~5.93%之間,赤眼鱒不同個體與青魚的遺傳變異率在19.09%~20.43%之間。4個不同群體赤眼鱒(WZ、XF、WH和YC)群體內的遺傳距離分別是0.012 0、0.016 2、0.009 9和0.004 8,群體間的遺傳距離值為0.007 0~0.049 1,總平均遺傳距離為0.029 3(不含外群)。Cytb序列中,各個體間的轉換位點為0~51,顛換位點為0~13,均為轉換大于顛換。兩大水系4個不同群體個體間的遺傳變異率在0~8.78%之間,而赤眼鱒不同個體與青魚的遺傳變異率在11.61%~12.48%之間。4個不同群體赤眼鱒(WZ、XF、WH和YC)群體內的遺傳距離分別是0.010 9、0.007 3、0.003 8和0.005 2,群體間的遺傳距離值為0.004 3~0.082 7,總平均遺傳距離為0.045 5(不含外群)(表2)。
4個群體的D-loop和Cytb序列的單倍型多樣性(H)、平均核苷酸差異數(K±SD)、核苷酸多樣性(π)見表3。4個群體間的FST及Nm值見表4。從表4可以看出:武漢群體與新豐群體及梧州群體的遺傳差異顯著,宜昌群體與新豐群體及梧州群體的遺傳差異顯著,而同一水系內的不同群體差異不顯著。把4個群體作為一個組進行群體間的分子變異等級分析(AMOVA)(表4),結果表明:不管D-loop,還是Cytb,群體間都存在顯著性遺傳差異(FST=0.689 77或0.871 94,P=0.000 0<0.05)。
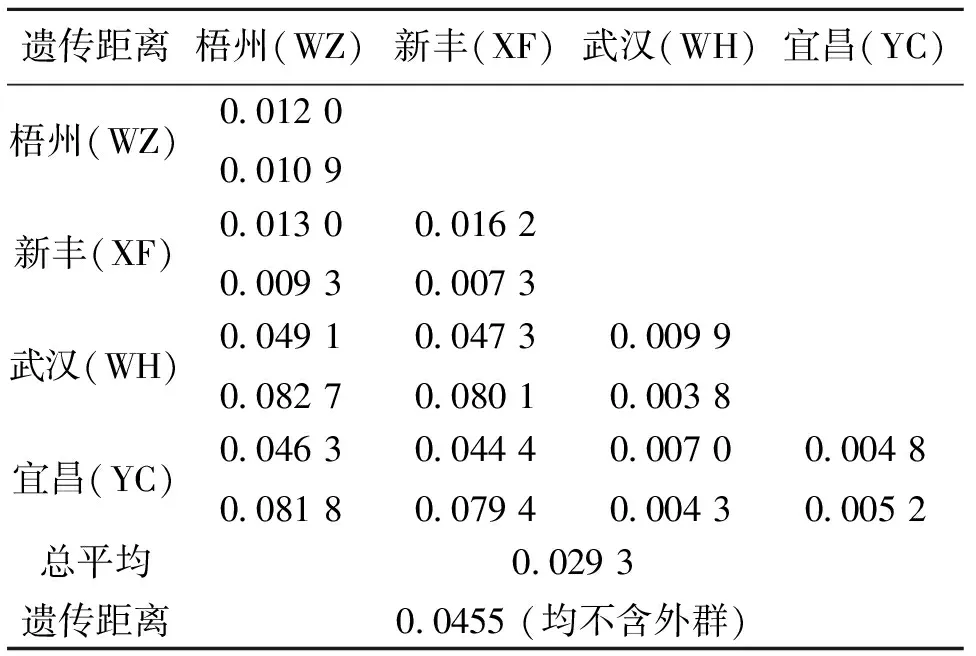
表2 兩大水系4個不同水域赤眼鱒D-loop和Cyt b基因序列群體內(對角線上)、群體間(對角線下)以及總平均(最后行)遺傳距離1)
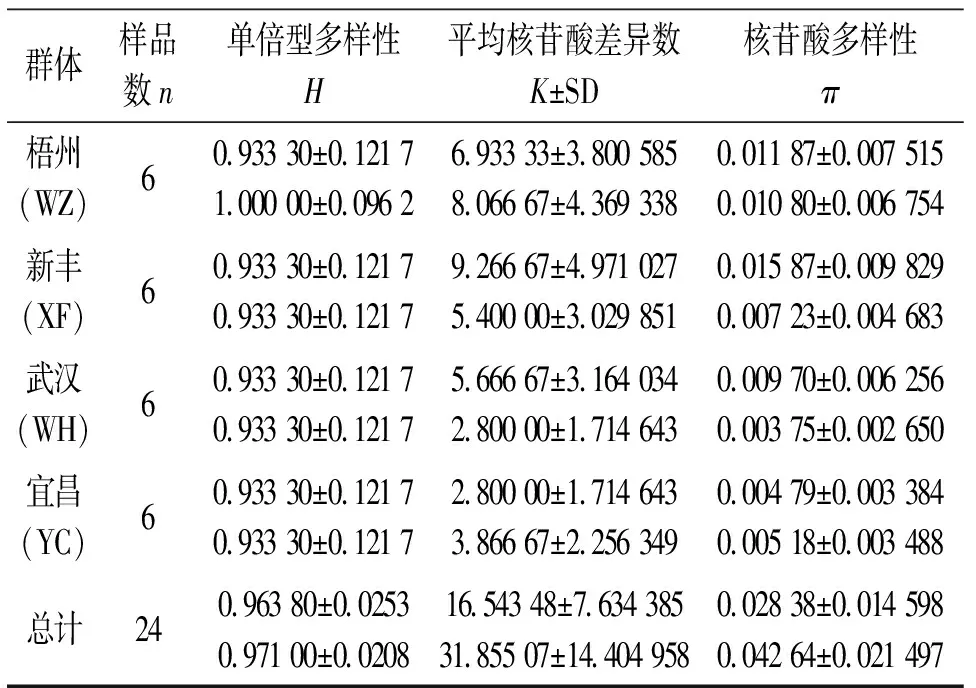
表3 4個赤眼鱒群體相關遺傳多樣性指標1)
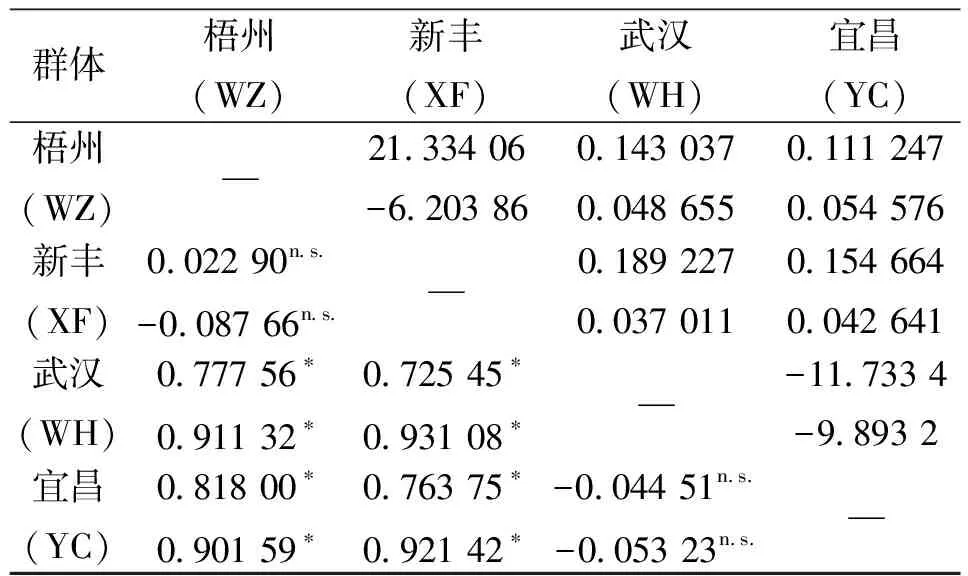
表4 群體間的FST(對角線下方)和Nm(對角線上方)1)

表5 群體間的遺傳變異的分子變異等級分析(AMOVA)1)
2.3 分子系統樹
以青魚MylopharyngodonpiceusD-loop和Cytb基因序列為外群,用MEGA4.1中的“NJ”法構建分子系統樹,其拓撲結構見圖1。用Bootstrap 1000給出各支的置信度,最高為100,最低為17。兩大水系4個不同水域赤眼鱒24個個體的系統樹明顯分為兩支:珠江12個個體聚為一支,長江12個個體聚為另一支,且都有較高的置信度,各大水系內的不同水域個體卻混雜在一起。
3 討 論
3.1 D-loop和Cyt b對赤眼鱒遺產多樣性的分析
單倍型多樣性(H)、平均核苷酸差異數(K±SD)和核苷酸多樣性(π)是反映物種遺傳變異水平的3個重要參數,赤眼鱒D-loop和Cytb的H、K和π值分別為0.963 8、16.543 48、0.028 38和0.971 0、31.855 07、0.042 6,與相同技術對光唇裂腹魚[22-25]的研究結果相比(表5),本研究中赤眼鱒各遺傳多樣性參數值較高。另外,根據堿基組成的基本信息,赤眼鱒具有較高的單倍型檢出率(數)、多態位點百分比(數)以及簡約信息位點百分比(數)[22-26]。所以,無論是D-loop,還是Cytb,兩者都很相似地得出赤眼鱒總體較高遺傳多樣性的研究結果。由于物種特定的遺傳結構、分類地位、生態習性及淡、海水魚類在基因交流方面的不同,可能導致它們彼此之間在遺傳分化水平上存在差異,但上述比較,在一定程度上能夠間接反映出赤眼鱒的遺傳多樣性狀況。
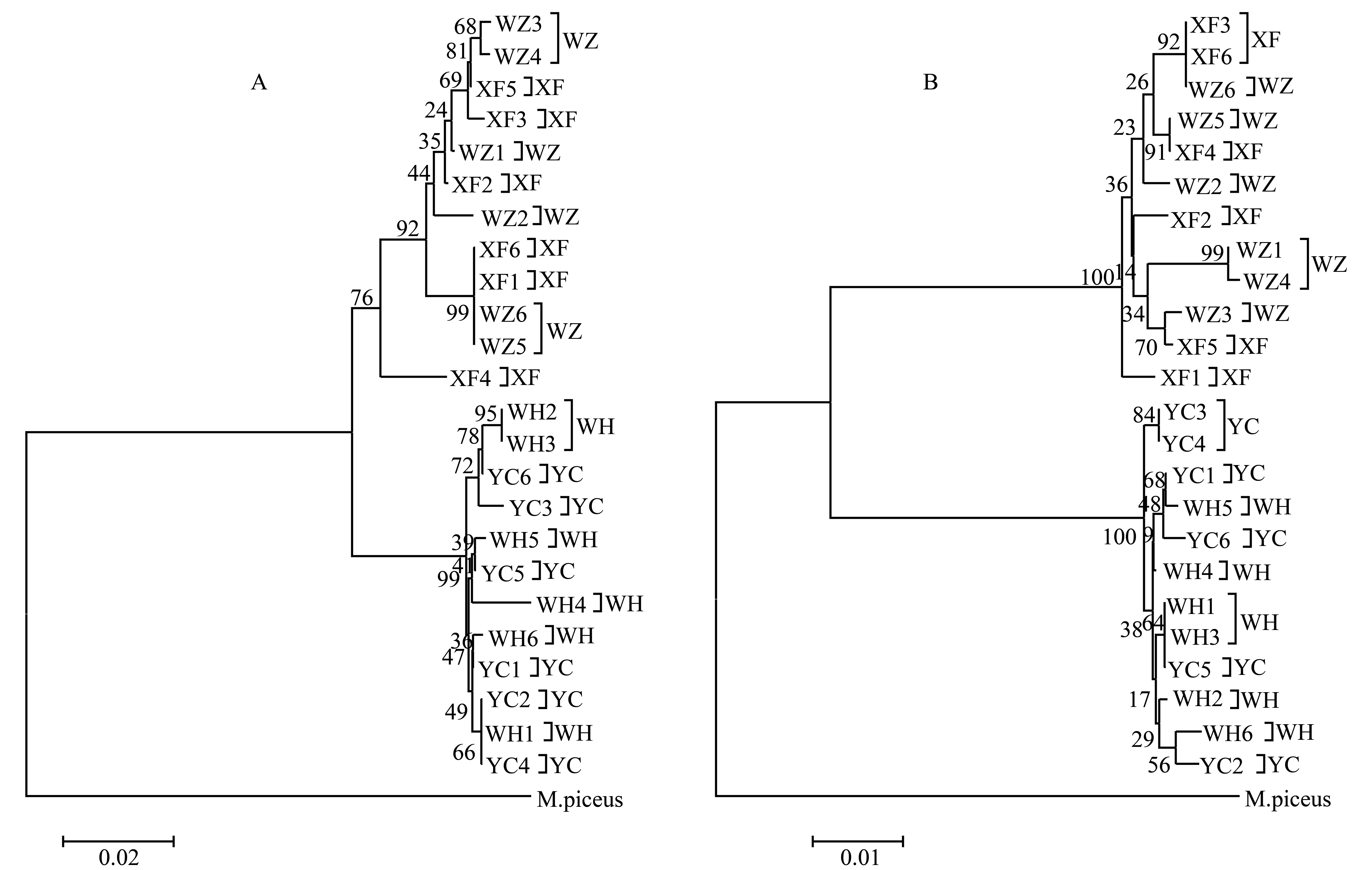
圖1 兩大水系4個不同水域赤眼鱒D-loop(A)和Cyt b(B)基因序列的NJ分子系統樹(數值表示1 000次重復抽樣的百分比)
一般來說,物種遺傳多樣性的高低與其適應能力、生存能力和進化潛力密切相關,遺傳變異是有機體適應環境變化的必要條件[27-28]。雖然赤眼鱒具有較高的遺傳多樣性水平,但由表2、4可以看出,D-loop和Cytb很一致的表明:長江和珠江水系間存在明顯的遺傳變異,可同一水系內群體間的遺傳變異和遺傳多樣性并不十分豐富。分子變異分析(AMOVA)結果也顯示:4個群體總的遺傳分化指數為0.689 77和0.871 94(P<0.001),其中68.98%、87.19%遺傳變異存在于群體間,群體內的遺傳變異率對總的遺傳變異貢獻較小。所以,在同一水系內部,赤眼鱒的遺傳多樣性狀況并不十分樂觀,加強對現有資源的科學管理和保護、恢復有效種群大小、豐富遺傳多樣性是保護赤眼鱒自然資源的有效舉措。
3.2 D-loop和Cyt b基因的變異位點比較
在所研究的不同水域赤眼鱒個體D-loop序列中檢測出了67個突變位點,多態位點的比例高達11.2%,再一次驗證了D-loop序列的高進化速率。其中插入/缺失位點14個,在檢測的Cytb中就沒有發現插入/缺失位點,在其他的研究中也證實了該觀點[29,30],這主要是由于Cyt b用于編碼蛋白質,如有插入/缺失的發生很容易被淘汰;而D-loop片段為非編碼基因,主要與線粒體基因的調控有關,不受蛋白功能上的需要和三聯體密碼的限制,所以插入/缺失突變后比較容易被保留。在D-loop和Cytb的變異位點中,轉換和顛換比值分別為3.192和4.636,通過軟件DAMBE分析所得的轉換、顛換對遺傳距離圖可知:D-loop和Cytb的轉換和顛換數沒有達到平臺效應,即沒有達到飽和狀態,說明供統計分析的數據準確可靠,序列中的堿基替代均具有系統發育意義[31],可以用來作為赤眼鱒種內的遺傳分析。統計每個個體兩兩之間的轉換和顛換數,每條序列的轉換均大于顛換,Gerlach等[32]的研究成果也證實了該觀點,其原因仍不清楚,需要進一步研究。在D-loop和Cytb基因堿基組成中,A+T含量均高于C+G含量,且D-loop基因中A+T含量要高出Cytb近11.1%,這與其他的研究結果相一致[33,34]。
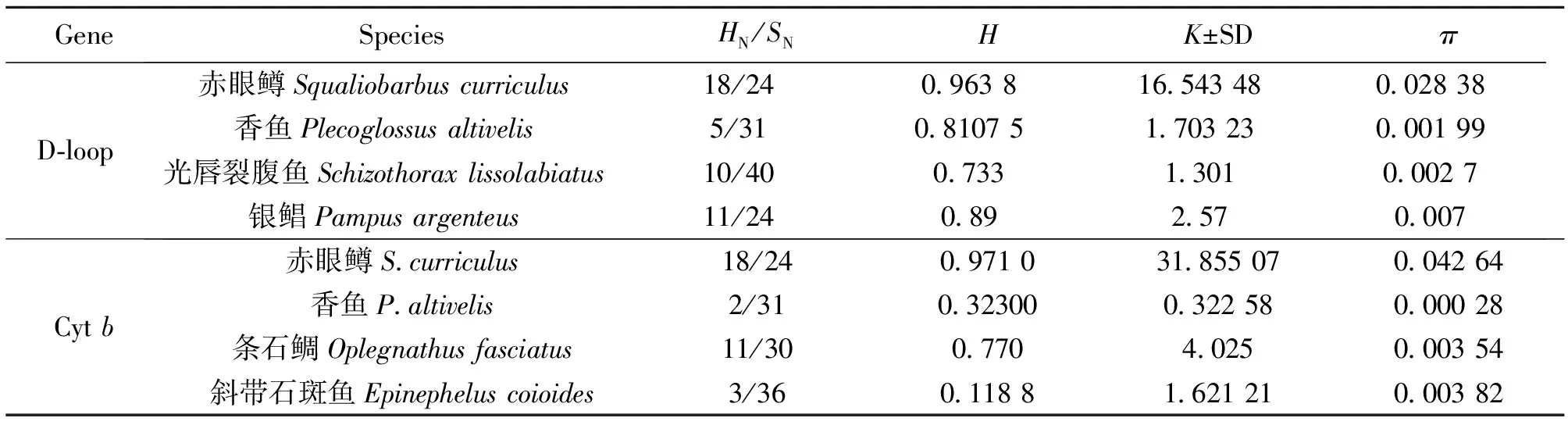
表6 不同物種相關遺傳多樣性指標比較1)
3.3 D-loop和Cyt b基因的進化比較
線粒體基因組的特性使它已成為進化生物學和群體遺傳學研究中重要的分子標記。目前,研究最多的是Cyt b、12S rRNA、16S rRNA、COⅠ基因和D-loop區[22-26,30,34-35]。本研究中,盡管D-loop和Cytb很一致的表明長江和珠江水系赤眼鱒存在明顯的遺傳分化,但同一水系內群體間的遺傳變異并不豐富,且D-loop和Cytb基因之間也存在差異。由群體間的遺傳距離值和FST值(表2和表4)可以看出,對于同一水系群體間(如WZ-XF,YC-WH)的遺傳差異較小,Cytb則更小,對于不同水系群體間(如WZ-YC和WH,XF-YC和WH)的遺傳差異較大,Cytb則更大,呈現出“小則越小,大則越大”的態勢。分析其原因,可能與D-loop和Cytb進化速率密切相關。
有研究表明,線粒體不同基因的進化速率存在差異[37]。D-loop區由于不編碼蛋白,受到的選擇壓力最小,所以進化速度最快[23,36]。Cytb基因由于編碼線粒體細胞色素b蛋白,其進化速度適中,適合研究種內到種間甚至科間的系統發育關系,近年來廣泛地應用于爬行類、魚類等的系統發育、種類鑒別等的研究[38-40]。蒙子寧等[26]認為,石斑魚線粒體細胞色素b基因序列在種內變異極小、變異位點數和平均核苷酸差異數等均非常低;但該基因序列在石斑魚種間卻存在較大的核苷酸差異,UPGMA系統樹更是以100%的置信度將斜帶石斑魚和赤點石斑魚區分開。由此可見,Cytb基因具有保守和變異的雙重特性、保守性決定了該基因不適合做石斑魚的種內遺傳變異分析;但其變異性使得該基因適合于種以上水平的種類鑒定。李娜等[23]的研究中,將中國香魚與日本香魚的Cytb基因比對后,僅發現2個變異位點,說明Cytb基因在香魚種內的變異很小,所以作者認為其不適合作為種內遺傳分析的標記。而在D-loop區發現17個變異位點,說明香魚與其他脊椎動物一樣線粒體D-loop區的進化速度較快。
據此分析,是否可以推測:Ctyb在遺傳變異顯著(或遺傳分化明顯)的群體中變異較大,更適合種間或以上階元系統進化分析,而D-loop在遺產差異不顯著的群體中變異相對較大,適合同種不同地理居群的多樣性分析?
由于各種遺傳分析方法都有優缺點,因此在實踐應用中,需根據研究材料、目的不同,選擇不同的分析方法,且最好是幾種方法同時運用,將結果相互印證,以便使得到的結論更客觀,更真實。
參考文獻:
[1]楊干榮,黃宏金.雅羅魚亞科[M] // 伍獻文.中國鯉科魚類志.上海: 上海科學技術出版社,1982: 52-53.
[2]李桂峰,康裕財,孫際佳,等.酵母多糖對赤眼鱒非特異性免疫技能的影響[J].中山大學學報:自然科學版,2003,42(4): 55-58.
[3]何學福,陽清發.嘉陵江西河赤眼鱒的生長研究[J].西南師范大學學報:自然科學版,1997,22(6): 680-685.
[4]楊太有,關建義,陳宏喜,等.丹江口水庫赤眼鱒(Squaliobarbuscurriculus)遺傳多樣性的RAPD和ISSR分析[J].海洋與湖沼,2008,39(2): 157-161.
[5]楊太有,關建義,陳宏喜.三個地理群體赤眼鱒遺傳多樣性的ISSR分析[J].水生生物學報,2008,32(4): 529-533.
[6]楊太有,關建義,陳宏喜,等.三個地理群體赤眼鱒遺傳多樣性的RAPD分析[J].四川動物,2008,27(5): 783-784.
[7]郭新紅,劉少軍,劉巧,等.魚類線粒體DNA研究新進展[J].遺傳學報,2004,31(9): 983-990.
[8]肖武漢,張亞平.魚類線粒體DNA的遺傳與進化[J].水生生物學報,2000,24(4): 384-391.
[9]LEE W J,KOCHER T D.Complete sequence of a sea Lamprey (Petromyzoamarinus) mitochondrial genome: Early establishment of the vertebrate genome organization [J].Genetics,1996,139: 873-887.
[10]葛隴利,赫崇波,高祥剛,等.利用mtDNA控制區序列分析斑點叉尾鮰的遺傳多樣性[J].水產科學,2007,26(10): 547-550.
[11]MOMMSEN H.Biochemistry and molecular biology of fishes[M].London: Elsevier Science Publishiers,1993: 1-38.
[12]XIAO Wuhan,ZHANG Yaping,LIU Huanzhang.Molecular systematics of Xenocyprinae(Telestei: Cyprinidae): taxonomy,biogeography,and coevolution of a special group restricted in east Asia[J].Mol Phylogenet Evol,2001,18: 163-173.
[13]韓虎峰,代應貴.珠江水系光唇裂腹魚可渡河種群mtDNA D-loop序列多態性分析[J].中國水產科學,2010,17(1): 143-149.
[14]SAMBROOK J,RUSSELL D.分子克隆實驗指南 [M].2版.金冬雁,黎孟楓,譯.北京: 科學出版社,1996: 464-469.
[15]唐瓊英,劉煥章,楊秀平,等.沙鰍亞科魚類線粒體DNA控制區結構分析及系統發育關系的研究[J].水生生物學報,2005,29(6): 645-653.
[16]THOMPSON J D,GIBSON T J,PLEWNIAK F,et al.The Clustal_X windows interface: flexible strategies for multiple sequences alignment aided by quality analysis tools[J].Nucleic Acids Research,1997,25 (24): 4876-4882.
[17]SCHNEIDER S,ROESSLI D,EXCOFFIER L.ARLEQUIN,version2000: a software for population genetics data analysis[M].Geneva: University of Geneva,2000.
[18]WRIGHT S.The interpretation of population structure byF2statistics with special regard to systems of mating[J].Evolution,1965,19: 395-420.
[19]NEI M.Evolution of human races at the gene level[M] // BONNE T B,COHEN T,GOODMAN R M,et al.Human Genetics,Part A: The unfolding genome.New York: Alan R Liss,1982: 167-181.
[20]EXCOFFIER L,SMOUSE P E,QUATTRO J M.Analysis of molecular variance inferred from metric distances among DNA heliotypes: application to human mitochondrial DNA restriction data[J].Genetics,1992,131: 479-497.
[21]KUMAR S,TAMURA K,JAKOBSEN I B,et al.MEGA2.0: molecular evolutionary genetics analysis software[M].Tempe: Arizona State University,2001.
[22]韓虎峰,代應貴.珠江水系光唇裂腹魚可渡河種群mtDNA D-loop序列多態性分析[J].中國水產科學,2010,17(1): 143-149.
[23]李娜,陳少波,謝起浪,等.閩浙地區香魚線粒體Cytb基因和D-loop區序列多態性分析[J].遺傳,2008,30(7): 919-925.
[24]彭士明,施兆鴻,陳超,等.根據mtDNA D-loop序列分析東海銀鯧群體遺傳多樣性[J].海洋科學,2010,34(2): 28-32.
[25]孫鵬,尹飛,彭士明,等.條石鯛線粒體 COⅠ和 Cytb序列的遺傳變異分析[J],水產學報,2011,35(5): 327-333.
[26]蒙子寧,楊麗萍,吳豐,等.斜帶石斑魚、赤點石斑魚RAPD和線粒體Cytb基因序列變異分析[J].中山大學學報:自然科學版,2007,46(1): 75-80.
[27]CONRAD M.Adaptability:The significance of variability from molecular to ecosystem[M].New York: Plenum Press,1983.
[28]FERAL J P.How useful are the genetic markers in attempts to understand and manage marine biodiversity[J].Journal of Experimental Marine Biology and Ecology,2002,268: 121-145.
[29]唐瓊英,楊秀平,劉煥章.刺鲃基于線粒體細胞色素b基因的生物地理學過程[J].水生生物學報,2003,27(4): 352-356.
[30]BILLINGTON N,HEBERT P D N.Mitochondrial DNA diversity in fishes and its implications for introductions[J].Canadian Journal of Fisheries and Aquatical Sciences,1991,48(suppl.1): 80-94.
[31]KOCHER T D,WILSON A C.Sequence evolution of mitochondrial DNA in humans and chimpanzees: Control region and a protein-coding region[M] // OSAWA S ,HONJO T.Evolution of life: Fossils,molecules and culture.Tokyo: Springer-Verlag,1991: 391-413.
[32]GERLACH G,MUSOLF K F.Fragmentation of landscape as a cause for genetic subdivision in bank voles[J].Conservation Biology,2000,14: 1066-1074.
[33]TANG Qiongying,LIU Huanzhang,MAYDEN R,et al.Comparison of evolutionary rates in the mitochondrial DNA cytochromebgene and control region and their implications for phylogeny of the Cobitoidea (Teleostei: Cypriniformes)[J].Molecular Phylogenetics and Evolution,2006,39: 347-357.
[34]孫希福,高天翔,竇存銀,等.江豚線粒體Cytb和D-loop片段序列分析及其分子系統學研究[J].南方水產,2009,5(5): 24-31.
[35]梁華芳,徐曉鵬,黃志堅,等.中國沿海龍蝦屬8種龍蝦COI基因序列的分子系統學研究[J].中山大學學報:自然科學版,2011,50(6): 94-98.
[36]黃志堅,徐曉鵬,唐晶晶,等.淡水魚類線粒體DNA D-loop基因的引物設計和應用[J].中山大學學報:自然科學版,2009,48(4): 84-88.
[37]MEYER A.Evolution of mitochondria DNA in fishes[M]//MOMMSEN T P,HOCHACHKA P W.Biochemistry and Molecular Biology of Fishes,Vol.2.New York: Elsevier,1993: 1-38.
[38]周繼亮,張亞平,黃美華,等.蝮亞科蛇線粒體細胞色素b基因序列分析及其系統發育[J].動物學報,2001,47(4): 361-366.
[39]FARIAS I P,ORTI G,SAMPAIO I,et al.The cytochromebgene as a phylogenetic marker: the limits of resolution for analyzing relationships among cichlid fishes[J].J Mol Evol,2001,53(2):89-103
[40]項方,鄒記興,鄧鳳姣,等.用細胞色素b部分序列研究斑馬魚的分子分類與系統發育[J].動物學雜志,2004,39(5): 13-18.
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
財經界(學術版)(2015年20期)2015-12-23 09:20:13
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
