地塞米松和飼糧能量水平對肉仔雞能量采食及神經肽Y基因表達的影響
2012-06-06 09:47:36磊宋志剛孔焦洪超
動物營養學報 2012年1期
劉 磊宋志剛孔 雪 焦洪超 林 海
(山東農業大學動物科技學院,泰安 271018)
隨著養殖業逐步向規模化、集約化發展,養殖過程中存在的諸多應激因素亦因之受到重視,如高溫、擁擠、免疫接種、有害氣體等。面對應激,機體會產生一系列生理及神經內分泌的應答反應,表現出食欲、能量存儲以及體內能量流動模式的變化,動物的采食量及采食飼料的種類受到影響,最終造成生產性能及胴體品質的改變。研究應激對家禽食欲的影響機制對促進家禽生產和提高飼料效率意義重大。應激反應不僅降低了家禽生產性能,也影響到了能量的沉積和利用效率。皮質酮處理后的肉仔雞能量利用效率顯著降低[1],大量能量以脂肪形式被儲存[2]。而高能飼糧能夠緩解應激帶來的一些不利影響,如高能飼糧顯著提高了應激肉仔雞的采食量和體增重[3]。Zulkifli等[4]發現應激能夠激活家禽的下丘腦-垂體-腎上腺(HPA)軸,導致糖皮質激素的大量分泌,糖皮質激素與其受體結合后,能通過改變相關基因的轉錄和蛋白的表達來調控機體能量資源的重新分配。地塞米松(DEX)是一種人工合成的糖皮質激素,它能與糖皮質激素受體特異性的結合。相比其他人工合成的糖皮質激素而言,DEX作用時間較長,更能代表體內的糖皮質激素的作用模式[5]。Qi等[6]用1 mg/kg的 DEX注射大鼠,發現大鼠脂肪酸和葡萄糖代謝紊亂。本實驗室在肉仔雞上經過長期試驗確定按2 mg/kg DEX對肉仔雞進行長期(3~7 d)注射,可以成功誘導肉仔雞的糖脂代謝發生改變[7-9]。
神經肽Y(NPY)廣泛分布在中樞神經系統和外周組織,尤其在下丘腦弓形核(ARC)中高度表達,其促進攝食的功能不斷受到人們的廣泛關注。NPY神經元從ARC投射到腹內側核(VMN)、背內側核(DMN)和室旁核(PVN),形成了密集的NPY網絡,此網絡在攝食調控中起著重要的作用[10-11]。中樞注射 NPY 可提高鼠、雞、羊和豬的采食量。糖皮質激素受體也在家禽的下丘腦中廣泛的分布[12],而糖皮質激素是否會影響下丘腦中的NPY表達,尚未見報道。本試驗通過腹部注射DEX來模擬應激模型,旨在研究肉仔雞在應激狀態下對不同能量水平飼糧的偏嗜性,同時初步研究應激影響家禽采食行為的作用機理。
1 材料與方法
1.1 試驗設計
體重相近的180只23日齡的雄性愛拔益加(AA)肉雞隨機分成6組,每組3個重復,每個重復10只雞。試驗采用2×3析因設計,因素為DEX[處理(注射DEX 2 mg/kg)、未處理(注射等劑量生理鹽水)]和飼糧能量水平[高能(HE)、低能(LE)以及高能、低能自由采食(H-LE)]。H-LE飼糧在1組中同時放2個料槽,分別盛有HE飼糧和LE飼糧。預試期5 d,正試期7 d,為28~34日齡,每天在08:00和10:00之間進行注射,期間所有的雞自由飲水。DEX購自山東魯抗辰欣藥業有限公司,批號為H37021969,濃度為3.85 mg/mL。試驗飼糧組成及營養水平見表1。
1.2 試驗方法
1.2.1 數據記錄和樣品采集
每天記錄肉仔雞的體重和采食量,并計算采食量、體增重、耗料增重比和耗能增重比。34日齡注射后空腹2 h,每個重復隨機選取3只雞(每個處理 9只),翅下靜脈竇采血,肝素抗凝,3 000 r/min離心10 min,分離血漿,-20℃保存待測;然后將肉仔雞屠宰,剝取下丘腦,于液氮速凍后-80℃冰箱保存以備抽提總RNA;剝離腹脂和腸道(包括十二指腸、空腸和回腸,并去除腸道內容物),稱重并計算腹脂率和腸道指數。
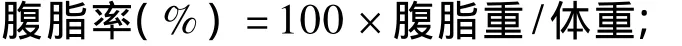
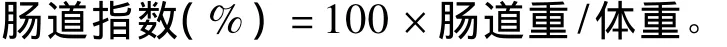
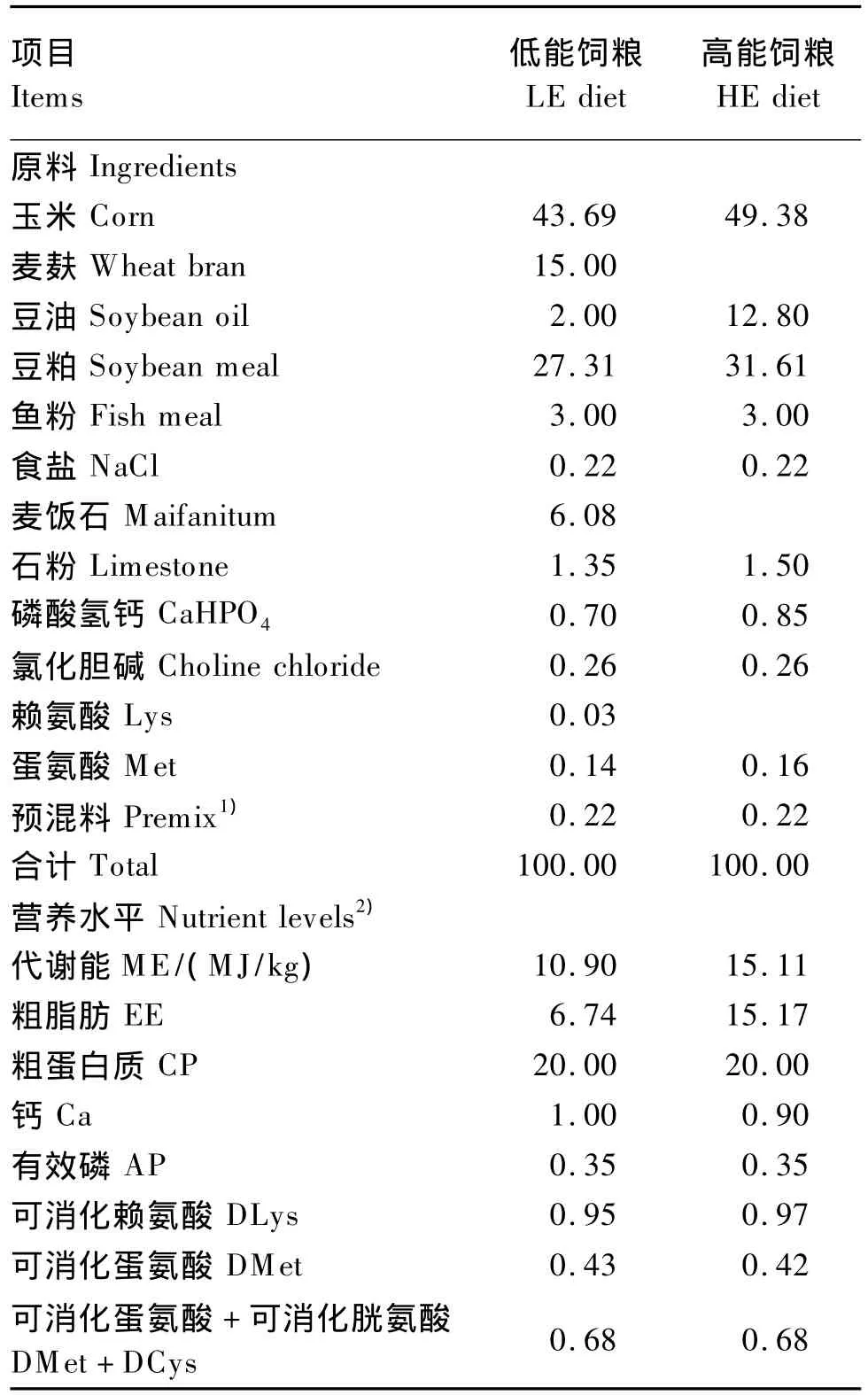
表1 試驗飼糧組成及營養水平(風干基礎)Table 1 Composition and nutrient levels of experimental diets(air-dry basis) %
1.2.2 血漿指標的測定
血漿中葡萄糖、尿酸和甘油三酯濃度采用VITALAB MICRO半自動生化儀測定,試劑盒購自南京建成生物技術研究所。
1.2.3 下丘腦NPY基因表達量測定
下丘腦總RNA用異硫氰二胍鹽法提取,利用瓊脂糖凝膠電泳和生物分光光度計(Biophotometer plusone,德國Eppendorf)分別檢測總RNA的質量和濃度。按照TaKaRa RNA PCR反轉錄試劑盒操作說明進行反轉錄。反應體系為:500 ng總RNA、5 mmol/L MgCl2、1 μL RT buffer、1 mmol/L dNTP、2.5 U 逆轉錄酶(AMV)、0.7 nmol/L oligo d(T)和 10 U Ribonuclease inhibitor,加 DEPC 水至10 μL。42 ℃反應 40 min,99 ℃ 滅活 5 min,降低到5℃ 5 min。
cDNA合成以后進行熒光定量PCR。磷酸甘油醛脫氫酶(GAPDH)(上游:5'-ACATGGCATCCAAGGAGTGAG-3', 下 游:5'-GGGGAGACAGAAGGGAACAGA-3')和NPY的引物(上游:5'-GAGGCACTACTCAACCTCATCAC-3',下 游:5'-TGTTTTCTGTGCTTTCCCTCAA-3')由上海生工生物技術有限公司合成。通過對混合樣品進行標準曲線測定確定引物的質量及最佳的稀釋濃度。反應體系為 20 μL:10 μL SYBR Premix Ex TaqTM(2 × )、上游和下游引物(10 μmol/L)各0.4 μL、0.4 μL ROX Reference Dye Ⅱ(50 × )、2 μL cDNA模板和6.8 μL去離子水。PCR反應程序為:95.0℃預變性10 s;95.0℃變性5 s,60℃延伸和退火34 s,40個循環。
參照 Livak 等[13]的方法用 2-ΔΔCt法定量目標基因相對表達量,以GAPDH作為參照基因進行校正[7]。
1.3 數據統計分析
試驗數據用平均值±標準誤表示,DEX和飼糧能量水平的互作效應采用SAS 9.1.3統計軟件ANOVA進程進行雙因子方差分析,以P<0.05為存在互作效應。差異顯著者用Duncan氏法進行多重比較,以P<0.05為差異顯著。
2 結果
2.1 DEX和飼糧能量水平對肉仔雞生產性能的影響
由表2可知,DEX處理極顯著降低了肉仔雞的采食量和體增重(P<0.01),極顯著提高了耗料增重比和耗能增重比(P<0.01)。飼糧能量水平對肉仔雞的體增重(P<0.05)、耗料增重比(P<0.05)和耗能增重比(P<0.01)有顯著影響;其中采用LE飼糧體增重顯著低于其他2種飼糧(P<0.05),耗料增重比顯著低于HE飼糧(P<0.05);采用HE飼糧耗能增重比極顯著高于其他2種飼糧(P<0.01),采用LE飼糧極顯著低于其他2組(P<0.01)。DEX、飼糧能量水平對耗料增重比(P <0.05)和耗能增重比(P <0.01)的影響存在顯著的互作效應。
2.2 DEX和飼糧能量水平對肉仔雞血漿生化指標的影響
由表3可知,DEX處理極顯著提高了肉仔雞血漿葡萄糖、甘油三酯和尿酸濃度(P<0.01),其中葡萄糖濃度提高了68.35%,尿酸濃度提高了208.47%;飼糧能量水平對血漿葡萄糖、甘油三酯和尿酸濃度影響不顯著(P>0.05);DEX、飼糧能量水平對血漿葡萄糖、甘油三酯和尿酸濃度的影響無顯著互作效應(P>0.05)。
2.3 DEX和飼糧能量水平對肉仔雞腸道指數、腹脂率和下丘腦NPY基因表達量的影響
由表4可知,DEX處理顯著提高了肉仔雞的腸道指數(P<0.05),極顯著提高了腹脂率(P<0.01)。飼糧能量水平對腹脂率有極顯著影響(P<0.01),對腸道指數無顯著影響(P>0.05);采用LE飼糧腹脂率極顯著低于其他2種飼糧(P<0.01)。DEX、飼糧能量水平對腸道指數和腹脂率的影響無顯著互作效應(P>0.05)。
DEX處理對肉仔雞下丘腦NPY基因表達量的影響不顯著(P>0.05)。采用H-LE飼糧NPY基因表達量顯著高于LE飼糧(P<0.05)。DEX、飼糧能量水平對肉仔雞下丘腦NPY基因表達量的影響無顯著互作效應(P>0.05)。
3 討論
3.1 DEX和飼糧能量水平對血漿生化指標的影響
家禽的肝臟是脂類生成的主要器官[14],DEX能夠促進肝臟內的脂類合成[8],肝臟內合成的甘油三酯被運輸到血液中,血液甘油三酯濃度得到提高。急性應激時,HPA軸通過與腎上腺素、胰高血糖素和交感神經系統的互作,提高血糖濃度,滿足機體重要器官對葡萄糖的需求。在長期應激反應中糖皮質激素具有重要作用,其作用主要表現為脂肪和肌肉組織的動員和肝臟糖異生過程的加強,導致血糖濃度和游離脂肪酸濃度升高[15-16]。本試驗結果表明,在DEX作用下,肉仔雞體內的血糖濃度顯著提高,增加了糖原的異生。Lin等[15]研究表明飼糧中添加30 mg/kg的皮質酮增強了肉仔雞機體蛋白質的分解代謝;Menconi等[17]在大鼠和小鼠肌小管的體外試驗中發現,DEX不僅抑制了蛋白質的合成代謝,還促進了它的分解代謝,在本試驗中,DEX提高了血漿中的尿酸濃度,與前人的研究一致。DEX對生長后期的肉仔雞處理7 d,血漿葡萄糖、甘油三酯和尿酸濃度都有相應變化,提示DEX已經改變了體內糖脂代謝過程。
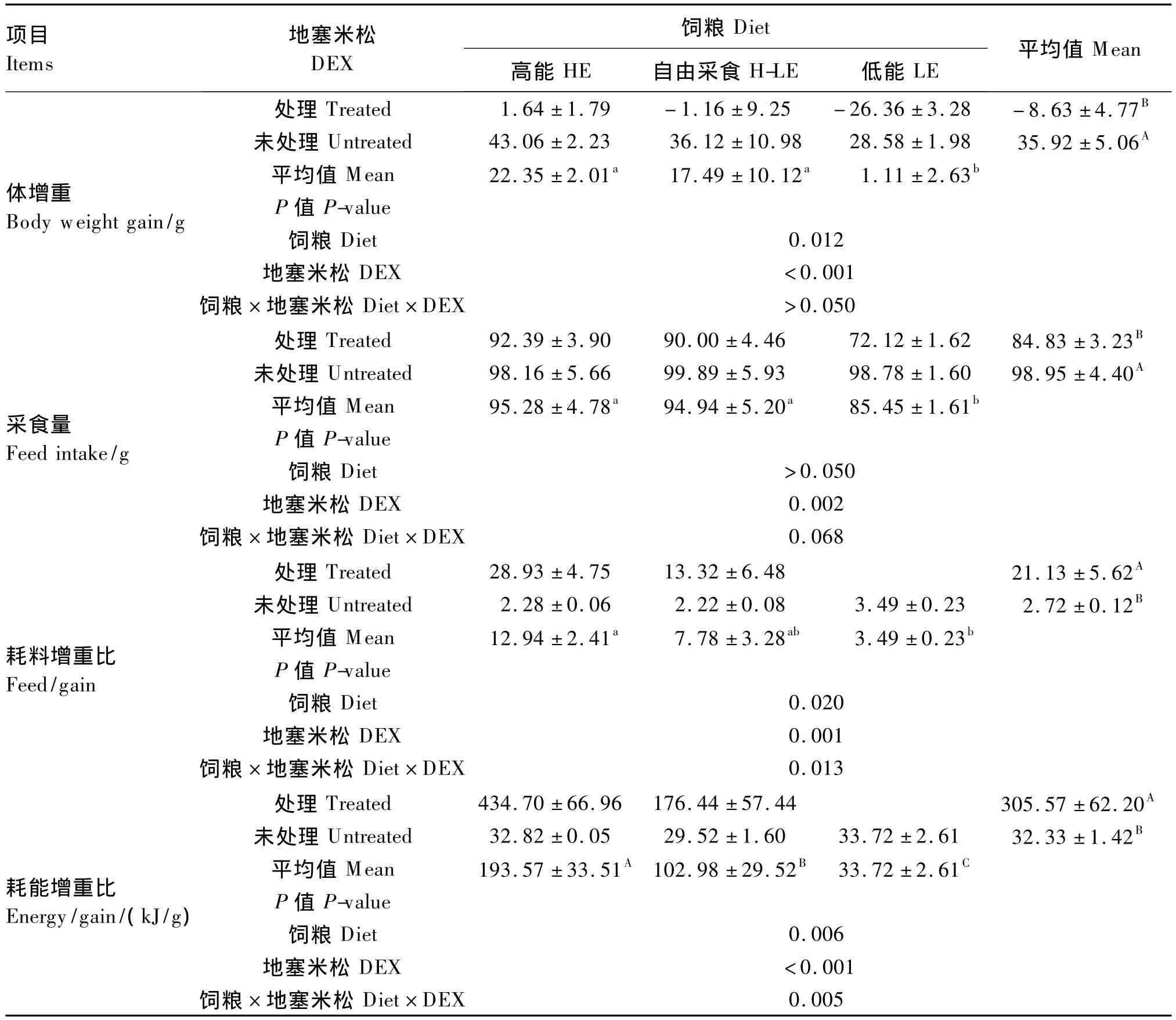
表2 DEX和飼糧能量水平對肉仔雞生產性能的影響Table 2 Effects of DEX and dietary energy level on performance of broiler chickens
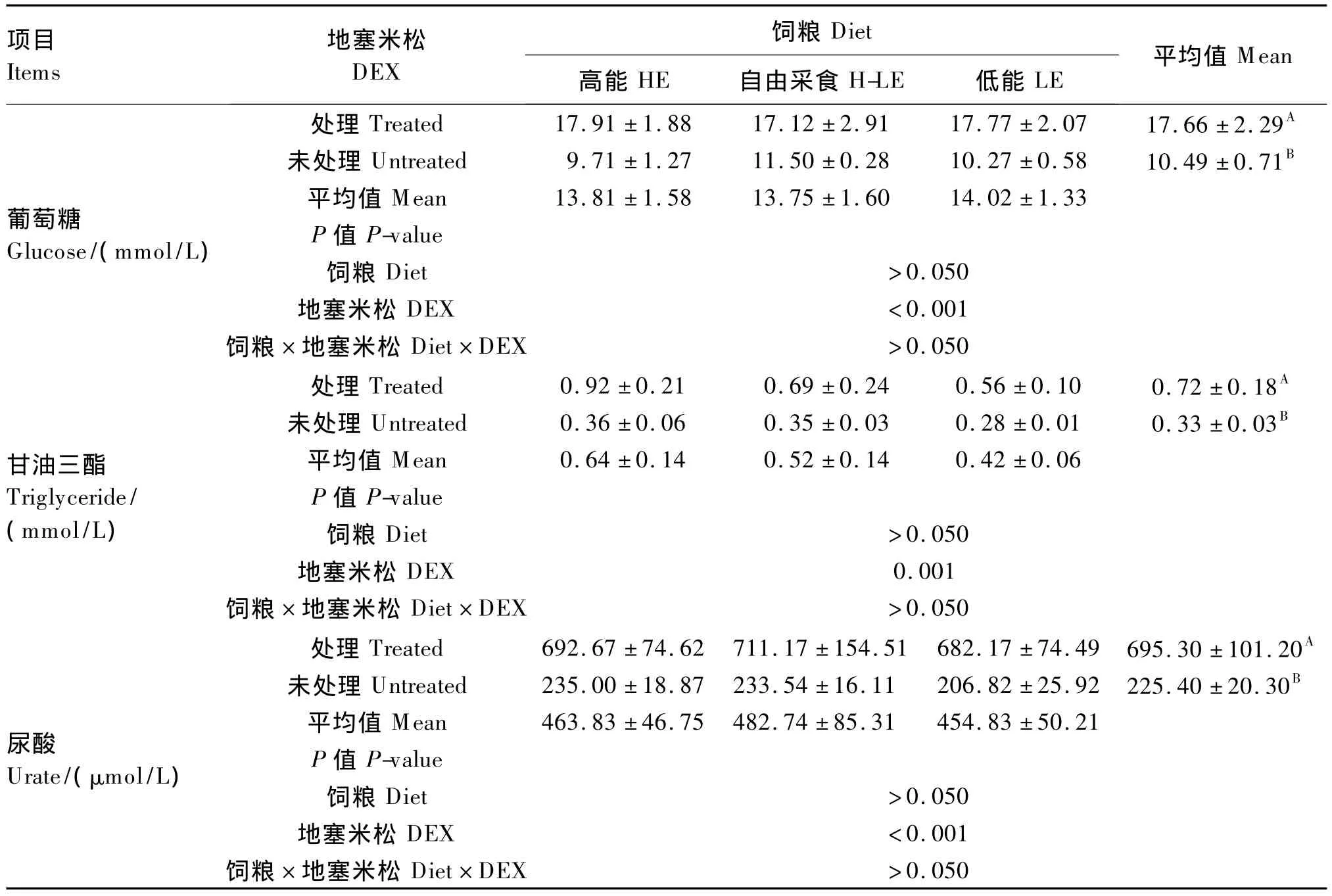
表3 DEX和飼糧能量水平對肉仔雞血漿生化指標的影響Table 3 Effects of DEX and dietary energy level on plasma biochemical indices of broiler chickens
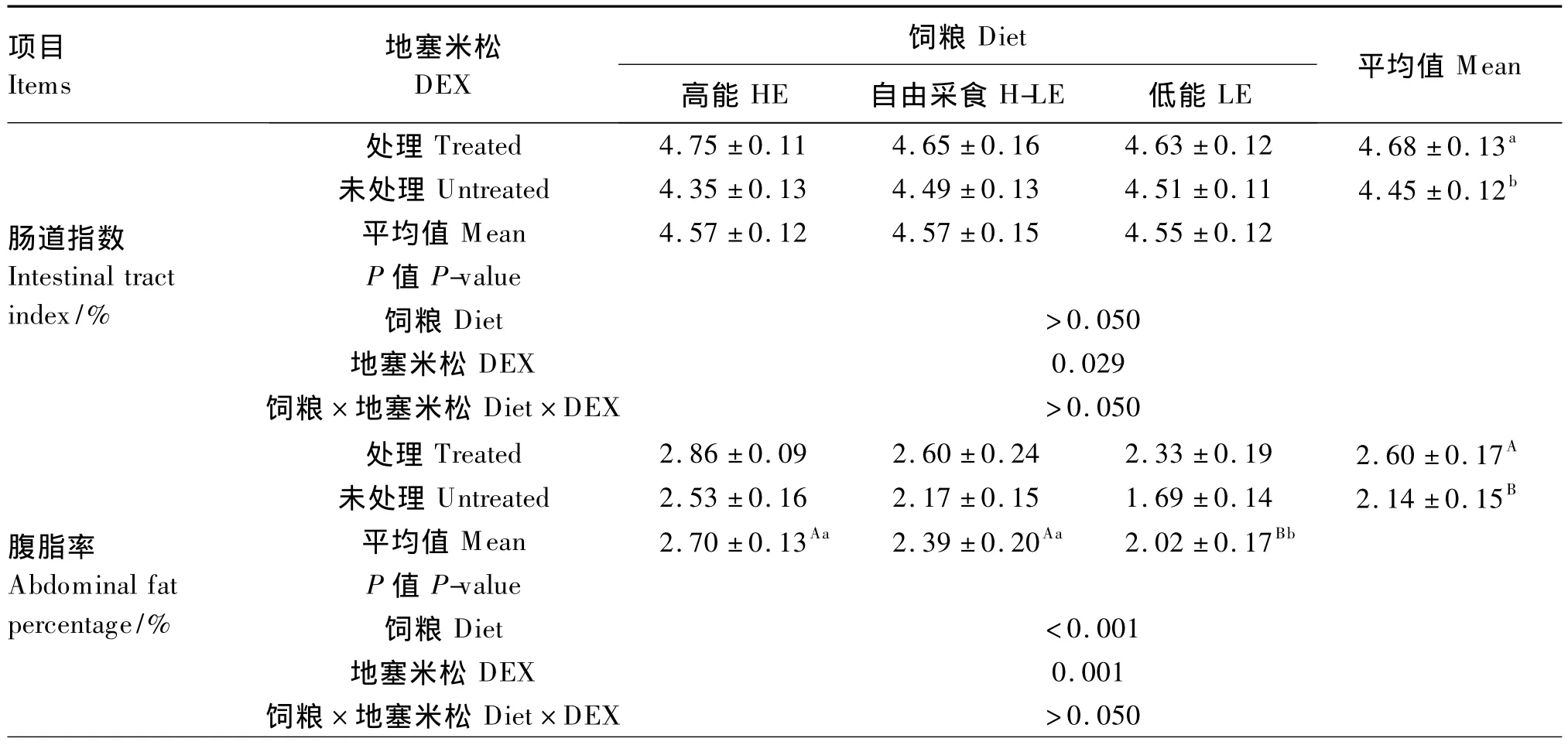
表4 DEX和飼糧能量水平對肉仔雞腸道指數、腹脂率和下丘腦NPY基因表達量的影響Table 4 Effects of DEX and dietary energy level on intestinal tract index,abdominal fat percentage and gene expression level of NPY in the hypothalamus of broiler chickens
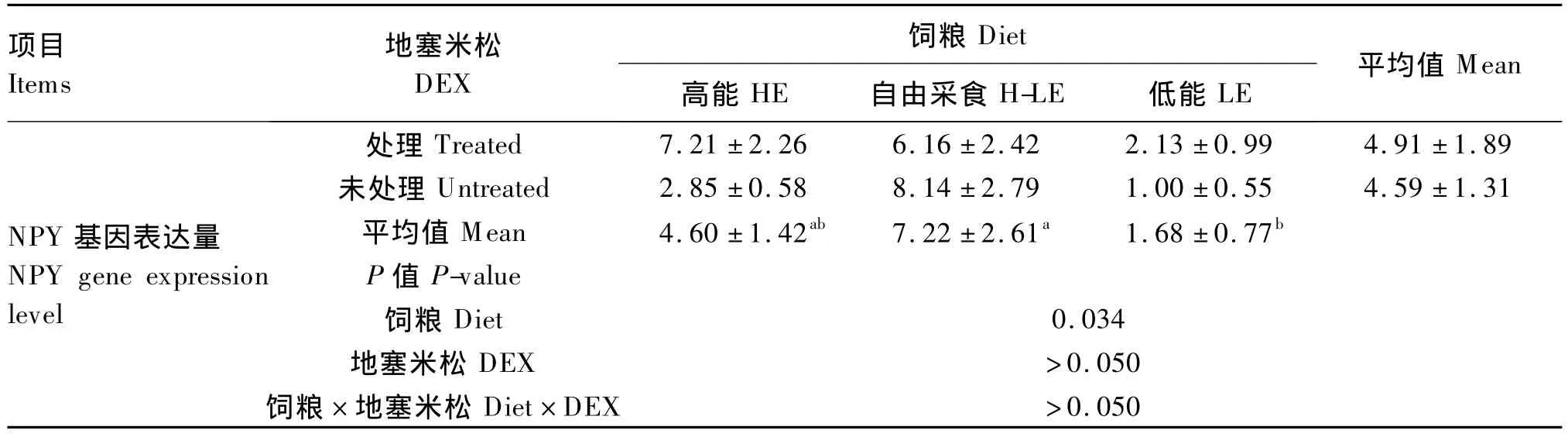
續表4
3.2 DEX和飼糧能量水平對能量采食的影響
機體通過控制采食量來調控機體的能量平衡,當應激或運動發生時,機體的能量消耗增加,采食量也隨之增加,使得耗料增重比降低。用外源導入方式進行的研究已證實皮質酮可導致家禽體重和飼料利用率降低[15,18]。Bartov 等[19]研究發現高濃度的糖皮質激素會引起肉仔雞負氮平衡,造成骨骼肌萎縮從而影響生長。Hayashi等[20]進一步研究發現糖皮質激素對肌肉生長的抑制作用歸因于骨骼肌蛋白質合成率的降低或蛋白質降解率的增加,同樣的結果也出現在本試驗中,DEX處理的肉仔雞體重不但不增加反而出現了降低的趨勢。據報道,高水平的糖皮質激素可增強小鼠腸系膜區域的脂肪沉積[21]。本試驗發現,DEX處理增強了肉仔雞腹部脂肪沉積,這與前人的研究結果一致[18,22]。可能的解釋是應激抑制了肌肉的發育并促進體內的脂肪沉積,從而表現為能量的分配由肌肉組織向脂肪組織轉移,造成了能量的浪費,使能量利用效率和飼料利用效率顯著降低。Elwinge等[23]已證明雞脂肪沉積量受飼糧的影響。高脂肪飼糧引起的脂蛋白脂酶活性變化,具有組織特異性,能夠提高脂肪組織脂蛋白脂酶的活性,并降低肌肉組織脂蛋白脂酶的活性,從而導致甘油三酯優先在脂肪組織沉積[24]。由此提示,HE飼糧可能會提高肉仔雞脂肪組織脂蛋白脂酶的活性,從而促進肉仔雞腹部等處的脂肪沉積,表現出耗能增重比低的現象。皮質酮處理后,家禽通過改變攝入食物類型或飼糧偏嗜性來試圖緩解[25]。在哺乳動物上的研究顯示,糖皮質激素可提高體內總能量的攝入量,并呈劑量關系;皮質酮處理使腎上腺切除后的小鼠采食更多的脂肪[26]。在本試驗中,DEX、飼糧能量水平對采食量的影響無顯著互作效應,但DEX注射下,動物對高能飼糧的采食量顯著增加,與前人研究結果一致。
胃腸容積是哺乳動物和家禽采食量的一個限制因素[27],哺乳動物的食道、胃、小腸中均存在著牽張感受器,采食后能激活迷走神經,從而興奮下丘腦飽感中樞,使動物停止采食。在本試驗中,DEX對腸道指數的影響可能是因為DEX降低了體增重,或是DEX處理造成能量以脂肪形式儲存,使機體對能量的需求增加,導致腸道的長度和重量增加以提高對營養物質和能量的攝取。
3.3 DEX和飼糧能量水平對下丘腦NPY基因表達量的影響
家禽的采食量是受多方面因素調控的,如體內一些激素(胰島素和瘦素)、食欲因子(NPY、縮膽囊素和饑餓素等)、環境以及營養水平,這些信號在下丘腦的核團中進行整合,共同完成對攝食行為的調節[28]。糖皮質激素通過其受體參與采食和能量平衡的中樞調控[16]。有證據顯示在NPY神經元上有糖皮質激素的受體,NPY的基因編碼區上游也攜帶糖皮質激素的應答片段[29]。體外和體內研究結果表明,糖皮質激素能夠激活NPY神經元的ARC-PVN路徑,使NPY合成量增加,并對ARC神經元表達 NPY 受體起正調節作用[30-31]。糖皮質激素還可能直接影響NPY基因表達,從而增加NPY的合成。Shimizu等[32]通過體外試驗發現糖皮質激素能增加下丘腦ARC中NPY的表達。在本試驗中,DEX處理提高了肉仔雞下丘腦NPY的表達量,但并沒有達到顯著水平,這可能是因為試驗重復數太少,也可能是腦內的多耐藥性糖蛋白作用,Meijer等[33]發現小鼠腦內的多耐藥性糖蛋白能把進入腦內的DEX不斷清除,這樣對下丘腦食欲基因產生作用的DEX量減少,不足以引起NPY基因表達量的顯著變化。但多耐藥性糖蛋白在家禽中還未見有關報道,是否是多耐藥性糖蛋白的作用結果還需進一步的研究。
另外,NPY也能感受能量信號,通過控制機體采食量,來協調動物采食量和能量的穩態平衡[32]。已有的研究對于高脂飼糧影響下丘腦NPY表達主要集中在哺乳動物上,結果并不一致。Giraudo等[34]研究發現,高脂飼糧降低了大鼠 ARC中NPY基因的表達;而Lin等[35]發現,飼喂1周高脂飼糧對大鼠下丘腦NPY基因表達的影響不顯著。但在本試驗中,肉仔雞采食H-LE飼糧時下丘腦NPY基因表達顯著增加,可能的解釋是NPY對高脂飼糧的響應具有物種特異性,或是由于肉仔雞可根據自己的需求而自由選擇能量的采食,肉仔雞體內的能量穩態處于一個相對穩定的動態過程,NPY感應到體內的能量狀態,對HE或LE飼糧產生了食欲感。
4 結論
①DEX應激可使肉仔雞血漿中葡萄糖、尿酸和甘油三酯的濃度升高。
②DEX應激和HE飼糧均能增加脂肪在腹部沉積,提高耗料增重比和耗能增重比。
③H-LE飼糧能上調肉仔雞下丘腦NPY基因表達。
[1]孔雪.糖皮質激素調控家禽采食的機制[D].碩士學位論文.泰安:山東農業大學,2010:35-47.
[2]袁磊.應激對肉雞采食量影響及其調節機制[D].碩士學位論文.泰安:山東農業大學,2007:31-45.
[3]姜克杰.應激和飼糧能量水平對肉雞生長發育和脂肪代謝的影響[D].碩士學位論文.泰安:山東農業大學,2007:15-16.
[4]ZULKIFLIA I,SIEGE P B.Is there a positive side to stress?[J].World’s Poultry Science Journal,1995,51(1):63-76.
[5]FOUCAUD L,NIOT I,KANDA T,et al.Indirect dexamethasone down-regulation of the liver fatty acidbinding protein expression in rat liver[J].Biochimica et Biophysica Acta,1998,1391(2):204-212.
[6]QI D,PULINILKUNNIL T,AN D,et al.Singledose dexamethasone induces whole-body insulin resistance and alters both cardiac fatty acid and carbohydrate metabolism [J].Diabetes,2004,53(7):1790-1797.
[7]CAI Y,SONG Z,WANG X,et al.Dexamethasoneinduced hepatic lipogenesis is insulin dependent in chickens(Gallus gallus domesticus)[J].Stress,2011,14(3):273-281.
[8]CAI Y,SONG Z,ZHANG X,et al.Increased de novo lipogenesis in liver contributes to the augmented fat deposition in dexamethasone exposed broiler chickens(Gallus gallus domesticus)[J].Comparative Biochemistry and Physiology Part C:Toxicology&Pharmacology,2009,150(2):164-169.
[9]WANG X,LIN H,SONG Z,et al.Dexamethasone facilitates lipid accumulation and mild feed restriction improves fatty acids oxidation in skeletal muscle of broiler chicks(Gallus gallus domesticus)[J].Comparative Biochemistry and Physiology Part C:Toxicology&Pharmacology,2010,151(4):447-454.
[10]TOMASZUK A,SIMPSON C,WILLIAMS G.Neuropeptide Y,the hypothalamus and the regulation of energy homeostasis[J].Hormone Research in Paediatrics,1996,46(2):53-58.
[11]KALRA S P,HORVATH T L.Neuroendocrine interactions between galanin,opioids,and neuropeptide Y in the control of reproduction and appetite[J].Annals of the New York Academy of Sciences,1998,863(1):236-240.
[12]KOVACS K J,WESTPHAL H M,PECZELY P.Distribution of glucocorticoid receptor-like immunoreactivity in the brain,and its relation to CRF and ACTH immunoreactivity in the hypothalamus of the Japanese quail,coturnix coturnix japonica[J].Brain Research,1989,505(2):239-245.
[13]LIVAK K J,SEHMITTGEN T D.Analysis of relative gene expression data using rea1-time quantitative PCR and the 2‐ΔΔCtmethod[J].Methods,2001,25(4):402-408.
[14]HERMIER D.Lipoprotein metabolism and fattening in poultry[J].The Journal of Nutrition,1997,127(5):805S-808S.
[15]LIN H,DECUYPERE E,BUYSE J.Oxidative stress induced by corticosterone administration in broiler chickens(Gallus gallus domesticus):1.Chronic exposure[J].Comparative Biochemistry and Physiology Part B:Biochemistry and Molecular Biology,2004,139(4):737-744.
[16]KITRAKI E,SOULIS G,GEROZISSIS K.Impaired neuroendocrine response to stress following a shortterm fat-enriched diet[J]. Neuroendocrinology,2004,79(6):338-345.
[17]MENCONI M,GONNELLA P,PETKOVA V,et al.Dexamethasone and corticosterone induce similar,but not identical,muscle wasting responses in cultured L6 and C2C12 myotubes[J].Journal of Cellular Biochemistry,2008,105(2):353-364.
[18]MALHEIROS R D,MORAES V M,COLLIN A,et al.Free diet selection by broilers as influenced by dietary macronutrient ratio and corticosterone supplementation.1.Diet selection,organ weights,and plasma metabolites[J].Poultry Science,2003,82(1):123-131.
[19]BARTOV I,JENSEN L S,VELTMANN,Jr.Effect of dietary protein and fat levels on fattening of corticosterone-injected broiler chicks[J].Poultry Science,1980,59(8):1864-1872.
[20]HAYASHI K,KAYALI A G,TOMITA Y.Reduction of corticosterone-induced growth impairment by testosterone and its mechanism[J].Journal of Animal Science and Technology,1992,63(10):1001 -1008.
[21]REBUFFE-SCRIVA M,WALSH U A,MCEWEN B,et al.Effect of chronic stress and exogenous glucocorticoids on regional fat distribution and metabolism[J].Physiology & Behavior,1992,52(3):583-590.
[22]LIN H,SUI S J,JIAO H C,et al.Impaired development of broiler chickens by stress mimicked by corticosterone exposure[J].Comparative Biochemistry and Physiology-Part A:Molecular&Integrative Physiology,2006,143(3):400-405.
[23]ELWINGER K.Performance and abdominal and carcass fat in broilers as influenced by strain and diet energy concentration[C]//Proceedings 6th European Poultry Conference. Hamburg: [s. n.],1980:256-263.
[24]MANTHA L,PALACIOS E,DESHAIES Y.Modulation of triglyceride metabolism by glucocorticoids in diet-induced obesity[J].American Journal of Physiology:Regulatory,Integrative and Comparative Physiology,1999,277(2):R455-R464.
[25]COVASAA M,FORBESA J M.Selection of foods by broiler chickens following corticosterone administration[J].British Poultry Science,1995,36(3):489-501.
[26]BLIGH M E,DOUGLASS L W,CASTONGUAY T W.Corticosterone modulation of dietary selection patterns[J].Physiology & Behavior,1993,53(5):975-982.
[27]DENBOW D M.Food intake control in birds[J].Neuroscience&Biobehavioral Reviews,1985,9(2):223-232.
[28]MAYER J,THOMAS D W.Regulation of food intake and obesity[J].Science,1967,156:328 -337.
[29]EDWARDS C M,ABBOTT C R,SUNTER D,et al. Cocaine-and amphetamine-regulated transcript,glucagon-like peptide-1 and corticotrophin releasing factor inhibit feeding via agouti-related protein independent pathways in the rat[J].Brain Research,2000,866(1/2):128-134.
[30]ZAKRZEWSKA J M,CHAUDHRY Z,NURMIKKO T J,et al.Lamotrigine(lamictal)in refractory trigeminal neuralgia:results from a double-blind placebo controlled crossover trial[J].Pain,1997,73(2):223-230.
[31]MATTERI R L.Overview of central targets for appetite regulation[J].Journal of Animal Science,2001,79(E-Suppl.):E148 - E158.
[32]SHIMIZU H,ARIMA H,OZAWA Y,et al.Glucocorticoids increase NPY gene expression in the arcuate nucleus by inhibiting mTOR signaling in rat hypothalamic organotypic cultures[J].Peptides,2010,31(1):145-149.
[33]MEIJER O C,DE LANGE E C,BREIMER D D,et al.Penetration of dexamethasone into brain glucocorticoid targets is enhanced in mdr1A P-glycoprotein knockout mice[J].Endocrinology,1998,139(4):1789-1793.
[34]GIRAUDO S Q,KOTZ C M,GRACE M K,et al.Rat hypothalamic NPY mRNA and brown fat uncoupling protein mRNA after high-carbohydrate or high-fat diets[J].American Journal of Physiology:Regulatory,Integrative and Comparative Physiology,1994,266:R1578-R1583.
[35]LIN S,STORLIEN L H,HUANG X F.Leptin receptor,NPY,POMC mRNA expression in the dietinduced obese mouse brain[J].Brain Research,2000,875:89-95.
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
學苑創造·A版(2015年11期)2016-01-14 09:03:27
中國火炬(2010年8期)2010-07-25 11:34:30
