烤煙幾個重要植物學性狀的遺傳分析
2012-07-31 07:22:16張興偉王志德劉艷華牟建民
中國煙草科學 2012年5期
張興偉,王志德,任 民,劉艷華,牟建民
(農業部煙草生物學與加工重點實驗室,中國農業科學院煙草研究所,青島 266101)
煙草(Nicotiana tobacumL.)是我國重要的經濟作物之一,其中烤煙每年種植面積及產量均占世界三分之一左右。烤煙的植物學性狀會直接影響煙葉產值及化學成分,因此受到研究者的關注。國內外多數研究認為,株高、葉數、葉長、葉寬等大多數性狀的遺傳以加性效應為主,顯性效應作用較小[1-6],只有少數性狀受顯性效應影響較大[4-6],基本無上位性效應[3]。本研究以烤煙品種丸葉和Coker319雜交,獲得雜交組合的P1、P2、F1、B1、B2和F2六個世代,以數量性狀“主基因+多基因”混合遺傳模型進行分析,對重要植物學性狀的遺傳率和基因效應等參數進行估計,旨在探討烤煙重要植物學性狀的遺傳模式,為烤煙株型育種提供理論依據。
1 材料與方法
1.1 試驗材料
本試驗采用來源于國家煙草中期庫的烤煙品種丸葉(P1,引自日本)和Coker319(P2,引自美國)為親本,構建 F1、F2、B1(F1×P1)和 B2(F1×P2)群體。2009年春季將全部供試材料種植于青島即墨實驗農場,株行距為55 cm×120 cm,親本和F1每小區種植15株,共45株,B1、B2和F2每小區種植75株,共225株。
1.2 試驗方法
1.2.1 性狀指標測定 性狀指標測定參照文獻[7]。
1.2.2 試驗設計及數據分析 隨機區組設計,3次重復。采用植物數量性狀主基因+多基因混合遺傳多世代聯合分析方法,對該組合6個世代的重要植物學性狀進行分析。通過極大似然法和 Iterated expectation and conditional maximization(IECM)估計各世代、各成分的分布參數,然后通過 Akaike′s information criterion(AIC)值選擇最佳模型,同時進行一組適合性檢驗,包括均勻性U12、U22和U32檢驗,Smirnov 檢驗(nW2)和 Kolmogorov 檢驗(Dn),根據檢驗結果選擇最優遺傳模型。采用最小二乘法從最優遺傳模型的各成分分布參數估計各基因效應值,分析各遺傳效應。
在主基因+多基因的混合遺傳模型中,表現型值(p)表示為群體平均數(m)、主基因效應(g)、多基因效應(c)和環境效應(e)之和,即 p=m+g+c+e,因此,表現型方差(σ2p)可表示為主基因方差(σ2mg)、多基因方差(σ2pg)和環境方差(σ2e)之和,即 σ2p=σ2mg+σ2pg+σ2e。
通過親本和 F1同質群體提供環境誤差方差的無偏估計,可估計環境方差σ2e,成分分布方差σ2f,群體表型方差 σ2p,進而可得到主基因和多基因的遺傳方差(σ2mg、σ2pg)和遺傳率(h2mg、h2pg)。
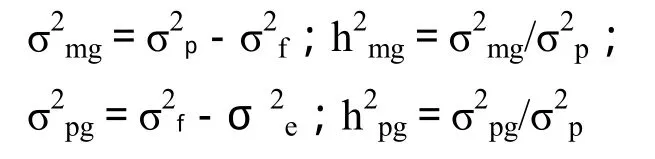
2 結 果
2.1 性狀遺傳方差分析及世代平均數分析
2個品種間幾個植物學性狀方差分析表明,重復間差異不顯著,處理間差異顯著(表1),說明2個親本重要植物學性狀在遺傳方面存在著真實穩定的差異,可以作為研究烤煙重要植物學性狀遺傳規律的材料進行分析。
從表1還可以看出,B1、B2和 F2的株高位于雙親之間,F1超過高值親本P1,說明株高的遺傳表現一定的超顯性效應,另外葉寬和節距的遺傳也表現一定的超顯性效應;葉數、莖圍遺傳表現一定的負超顯性效應,葉面積、比葉重遺傳表現一定的顯性效應,移栽至中心花開放天數、鮮葉重、葉形指數、葉長及蒴果重遺傳表現出一定的隱性效應。
2.2 性狀的遺傳模型
2.2.1 株高遺傳模型 用主基因+多基因混合遺傳模型對丸葉×Coker319組合6世代株高進行聯合分析,通過IECM算法獲得1對主基因(A)、2對主基因(B)、多基因(C)、1對主基因+多基因(D)和2對主基因+多基因(E)共5類24種遺傳模型的極大似然函數值和AIC值。根據AIC準則,E0、E1和E3的AIC值較小,作為備選遺傳模型。進一步對E0、E1和E3模型進行一組適合性[U12、U22、U32檢驗,Simirnov(nw2)檢驗和Kolmogorov檢驗(Dn)]檢驗,結果表明,在30個檢驗統計量中,E0模型沒有統計量與該模型差異達到顯著水平,E1模型有7個統計量達到顯著水平,E3模型有8個統計量達到顯著水平,綜合確定 E0為株高最優遺傳模型,即 2對加性-顯性-上位性主基因+加性-顯性-上位性多基因模型。
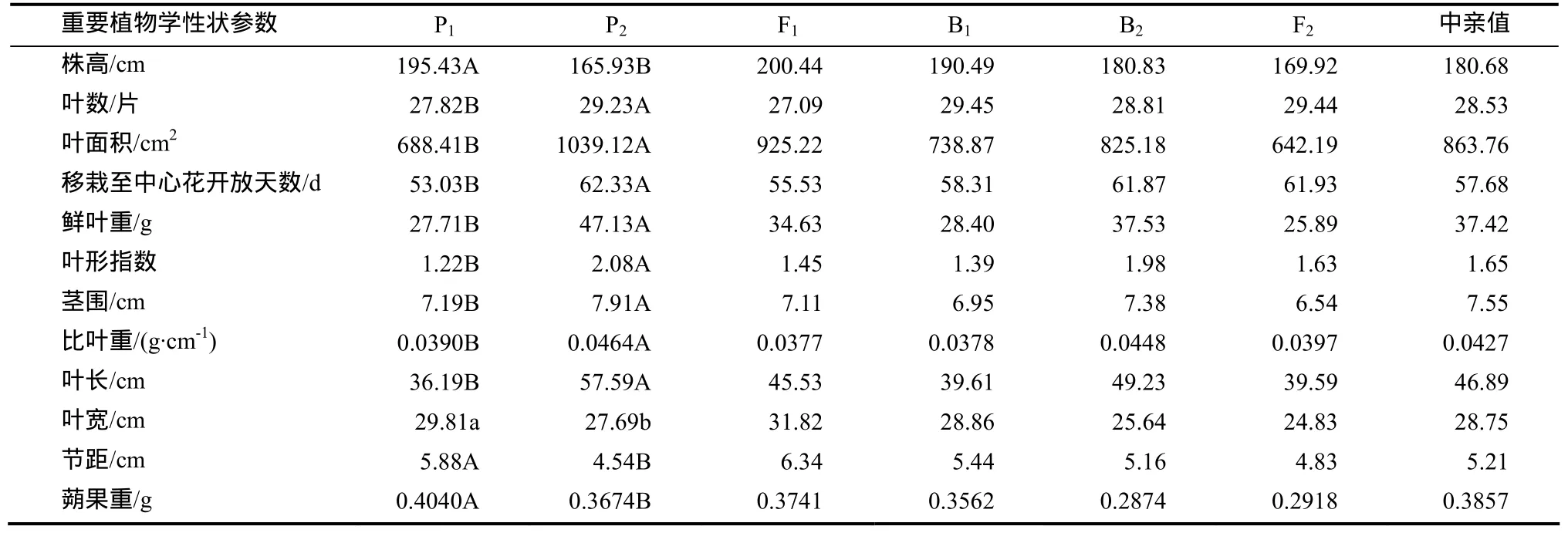
表1 各世代重要植物學性狀參數平均值Table 1 Means of the botanic trait parameters among six generations
2.2.2 葉數遺傳模型 通過IECM算法獲得5類24種遺傳模型的極大似然函數值和AIC值。根據AIC準則,B1、E0、E1和E3的AIC值較小,作為備選遺傳模型。適合性檢驗表明,在30個統計量中,E0模型沒有統計量達到顯著水平,其他幾個模型均有統計量達到顯著水平,綜合確定 E0為葉數最優遺傳模型,即 2對加性-顯性-上位性主基因+加性-顯性-上位性多基因模型。
2.2.3 葉面積遺傳模型 根據AIC準則,C0、D0、D3、E0和E1的AIC值較小,作為備選遺傳模型。適合性檢驗表明,在30個統計量中,C0、D0、D3和E0均只有2個統計量達到顯著水平,而E0的AIC值最小,所以選擇E0作為葉面積最優模型,即2對加性-顯性-上位性主基因+加性-顯性-上位性多基因模型。
2.2.4 葉長遺傳模型 根據 AIC準則,C0、D3、D4和E1的AIC值較小,作為備選遺傳模型。適合性檢驗表明,在30個統計量中,E1有11個統計量達到顯著水平,C0、D3和D4均有2個統計量達到顯著水平,但C0的AIC值最小,所以選擇C0作為葉長最優遺傳模型,即加性-顯性-上位性多基因遺傳模型。
2.2.5 葉寬遺傳模型 根據AIC準則,B1、C0和E1的AIC值較小,作為備選遺傳模型。適合性檢驗表明,在30個統計量中,C0只有1個統計量達到顯著水平,綜合確定C0為葉寬最優遺傳模型,即加性-顯性-上位性多基因模型。
2.2.6 葉形指數遺傳模型 根據 AIC準則,C0、D0、D3、E0和E1的AIC值較小,作為備選遺傳模型。適合性檢驗表明,在30個統計量中,C0、D0、E0和 E1均有 2個統計量達到顯著水平,而D0的AIC值最小,所以D0為葉形指數最優遺傳模型,即 1對加性-顯性主基因+加性-顯性-上位性多基因模型。
2.2.7 莖圍遺傳模型 根據 AIC準則,B1、D3、D4、E1和E3的AIC值較小,作為備選遺傳模型。適合性檢驗表明,在30個統計量中,D3和D4均沒有統計量達到顯著水平,而D3的AIC值最小,所以選擇D3作為莖圍最優遺傳模型,即1對完全顯性主基因+加性-顯性多基因模型。
2.2.8 節距遺傳模型 根據 AIC準則,C0、D0、E0和E1的AIC值較小,作為備選遺傳模型。適合性檢驗表明,在30個統計量中,C0、D0、E0和E1均沒有統計量達到顯著水平,選擇AIC值最小的C0為節距最優遺傳模型,即加性-顯性-上位性多基因模型。
2.2.9 移栽至中心花開放天數遺傳模型 根據AIC準則,B1、E0、E1和E3的AIC值較小,作為備選遺傳模型。適合性檢驗表明,在30個統計量中,E1只有3個統計量達到顯著水平,綜合考慮,選擇E1作為移栽至中心花開放天數最優遺傳模型,即2對加性-顯性-上位性主基因+加性-顯性多基因混合遺傳模型。
2.2.10 鮮葉重遺傳模型 根據AIC準則,D3、E0和E1的AIC值較小,作為備選遺傳模型。適合性檢驗表明,在30個統計量中,E0模型只有1個統計量達到顯著水平,綜合確定 E0為鮮葉重最優遺傳模型,即 2對加性-顯性-上位性主基因+加性-顯性-上位性多基因模型。
2.2.11 比葉重遺傳模型 根據AIC準則,C0、D0、D3、D4和E5的AIC值較小,作為備選遺傳模型。適合性檢驗表明,在30個統計量中,C0、D0、D3和D4模型均沒有統計量達到顯著水平,而D3的AIC值最小,綜合確定D3為比葉重最優遺傳模型,即1對完全顯性主基因+加性-顯性多基因模型。
2.2.12 蒴果重遺傳模型 根據AIC準則,C0、D0和E0的AIC值較小,作為備選遺傳模型。適合性檢驗表明,在30個統計量中,C0、D0和E0均只有1個統計量達到了顯著水平,而C0的AIC值最小,綜合確定C0為蒴果重最優遺傳模型,即加性-顯性-上位性多基因模型。
2.3 性狀遺傳效應預測及遺傳參數估計
2.3.1 株高遺傳效應分析 丸葉×Coker319組合的株高受2對加性-顯性-上位性主基因+加性-顯性-上位性多基因控制(表2),第1對主基因加性效應為19.65,顯性效應為5.99,第2對主基因加性效應為18.76,顯性效應為5.96,第1對主基因和第2對主基因加性×加性上位性效應為-14.44,第 1對主基因和第 2對主基因加性×顯性上位性效應為-2.10,第1對主基因和第2對主基因顯性×加性上位性效應為-1.26,第1對主基因和第2對主基因顯性×顯性上位性效應為-27.89,可見株高遺傳以加性效應及顯性×顯性上位性效應為主,但二者方向相反;B1、B2和 F2世代的主基因遺傳率分別為41.85%、51.71%和 57.53%;多基因遺傳率分別為25.50%、9.73%和16.18%。
2.3.2 葉數遺傳效應分析 葉數受 2對加性-顯性-上位性主基因+加性-顯性-上位性多基因控制(表2),第 1對主基因和第 2對主基因加性效應均為-2.65,顯性效應均為-0.95,顯性效應只有加性效應的35.85%,加性×加性上位性效應為2.03,加性×顯性上位性效應與顯性×加性上位性效應均為0.56,顯性×顯性上位性效應為 3.17,可見葉數遺傳以加性效應及顯性×顯性上位性效應為主,但二者方向相反,表現為正向中親優勢;B1、B2和 F2世代的主基因遺傳率分別為 36.91%、51.60%和42.63%;多基因遺傳率分別為 31.00%、16.84%和42.08%。
2.3.3 葉面積遺傳效應分析 葉面積受 2對加性-顯性-上位性主基因+加性-顯性-上位性多基因控制(表2),第1對主基因與第2對主基因的加性效應基本相當,均稍大于其顯性效應,加性×加性上位性效應與加性×顯性上位性效應、顯性×加性上位性效應、顯性×顯性上位性效應也基本相當,其中上位性效應>加性效應>顯性效應,上位性效應與加性效應及顯性效應方向相反,表現為正向中親優勢;B1、B2和F2世代的主基因遺傳率分別為2.13%、50.92%和30.32%;多基因遺傳率分別為19.53%、13.26%和12.18%。
2.3.4 葉長遺傳效應分析 葉長受加性-顯性-上位性多基因控制(表3),B1、B2和F2世代的多基因遺傳率分別為38.35%、71.54%和60.75%。
2.3.5 葉寬遺傳效應分析 葉寬受加性-顯性-上位性多基因控制(表3),B1、B2和F2世代的多基因遺傳率分別為46.66%、69.05%和62.14%。
2.3.6 葉形指數遺傳效應分析 葉形指數受1對加性-顯性主基因+加性-顯性-上位性多基因控制(表4),主基因加性效應與顯性效應基本相當,表現為正向中親優勢;B1、B2和 F2世代的主基因遺傳率分別為0.00%、35.43%和49.64%,多基因遺傳率分別為0.00%、46.95%和24.00%。
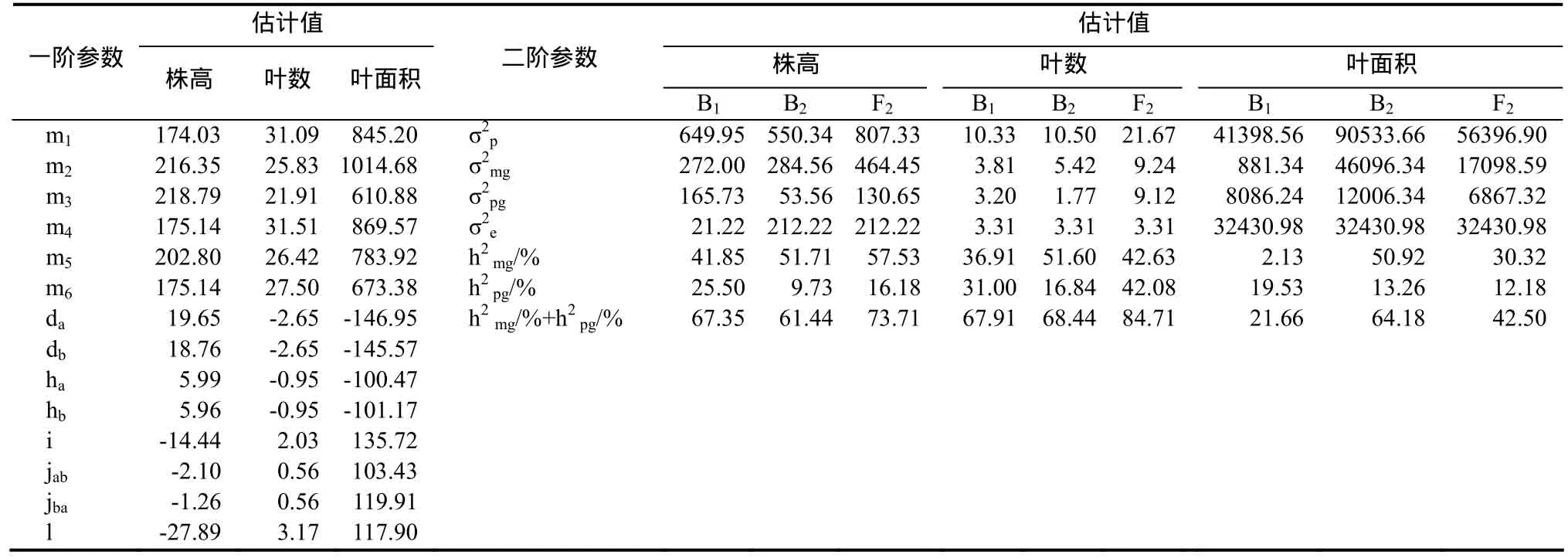
表2 株高、葉數和葉面積的E0模型遺傳參數Table 2 The estimate of plant height, leaf number and leaf area in E0 model
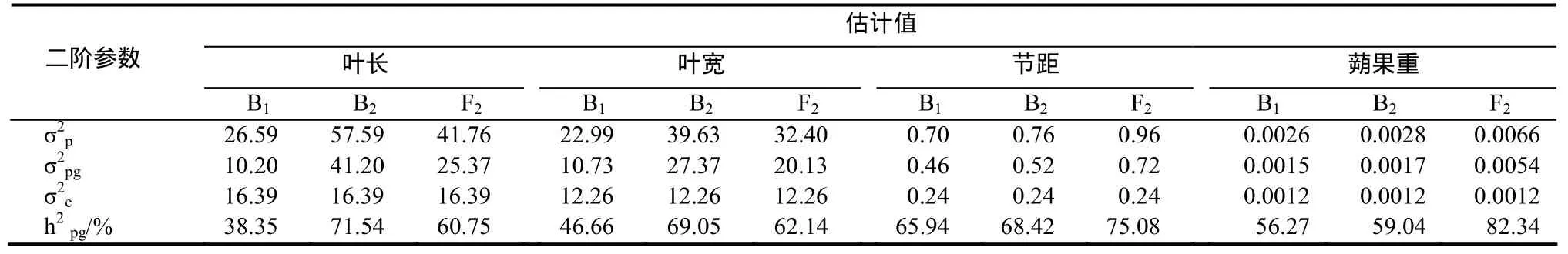
表3 葉長、葉寬、節距和蒴果重的C0模型遺傳參數Table 3 The estimate of leaf length, leaf width, intermodal distance and capsule weight in C0 model
2.3.7 莖圍遺傳效應分析 莖圍受1對完全顯性主基因+加性-顯性多基因控制(表4),主基因加性效應和顯性效應均為0.02,多基因加性效應為-0.43,多基因顯性效應為-0.58,可見莖圍遺傳以多基因為主,其多基因加性效應和顯性效應大小相當;B1、B2和F2世代的主基因遺傳率分別為8.84%、0.53%和2.48%;多基因遺傳率分別為46.48%、64.12%和53.97%。
2.3.8 節距遺傳效應分析 節距受加性-顯性-上位性多基因控制(表3),B1、B2和F2世代的主基因遺傳率均為0.00%,多基因遺傳率分別為65.94%、68.42%和75.08%。
2.3.9 移栽至中心花開放天數遺傳效應分析 移栽至中心花開放天數受 2對加性-顯性-上位性主基因+加性-顯性多基因控制(表5),第1對主基因加性效應為-7.90,顯性效應為-2.66,顯性效應只占加性效應的33.67%,第2對主基因加性效應為6.69,顯性效應為0.25,顯性效應只占加性效應的3.74%,第1對主基因與第2對主基因方向相反,加性×加性上位性效應為-8.39,加性×顯性上位性效應為0.53,顯性×加性上位性效應為-0.90,顯性×顯性上位性效應為-6.34,多基因加性效應為-2.80,多基因顯性效應為-1.44,可見移栽至中心花開放天數遺傳以加性×加性上位性效應、加性效應及顯性×顯性上位性效應為主,親本型上位性效應大于重組型上位性效應,主基因和多基因均表現為正向中親優勢;B1、B2和F2世代的主基因遺傳率分別為50.01%、59.73%和 64.79%;多基因遺傳率分別為 0.00%、7.24%和9.87%。
2.3.10 鮮葉重遺傳效應分析 鮮葉重受2對加性-顯性-上位性主基因+加性-顯性-上位性多基因控制(表5),第1對主基因和第2對主基因加性效應均為-7.90,顯性效應均為-7.79,加性×加性上位性效應為7.47,加性×顯性上位性效應為7.54,顯性×加性上位性效應為7.56,顯性×顯性上位性效應為9.51,可見鮮葉重遺傳幾種效應大致相當,其中上位性效應>加性效應>顯性效應,上位性效應與加性效應及顯性效應方向相反,表現為正向中親優勢;B1、B2和 F2世代的主基因遺傳率分別為 1.36%、55.38%和44.26%;多基因遺傳率分別為31.38%、18.45%和3.18%。
2.3.11 比葉重遺傳效應分析 比葉重受 1對完全顯性主基因+加性-顯性多基因控制(表4),主基因加性效應和顯性效應均為-0.0025,多基因加性效應為-0.0022,多基因顯性效應為-0.0019,可見比葉重遺傳主基因和多基因的加性效應和顯性效應大致相當;B1、B2和 F2世代的主基因遺傳率分別為0.60%、1.05%和 38.71%;多基因遺傳率分別為62.66%、77.66%和25.36%。
2.3.12 蒴果重遺傳效應分析 蒴果重受加性-顯性-上位性多基因控制(表3),B1、B2和F2世代的多基因遺傳率分別為56.27%、59.04%和82.34%。
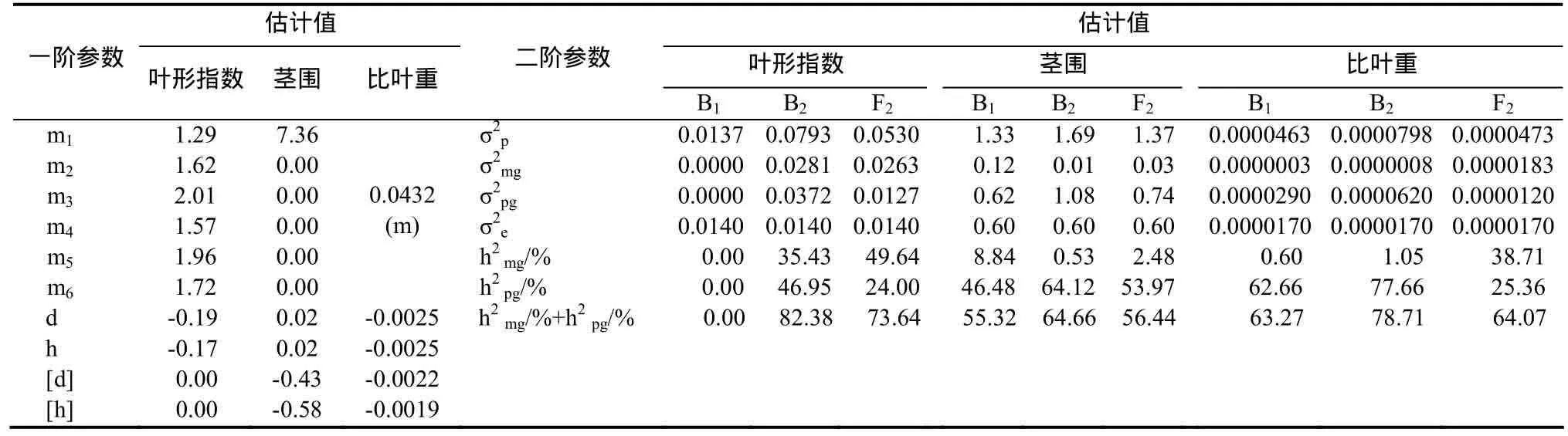
表4 葉形指數的D0模型和莖圍、比葉重的D3模型遺傳參數Table 4 The estimate of genetic parameters of leaf index in D0 model and stem girth, leaf mass per area in D3 model
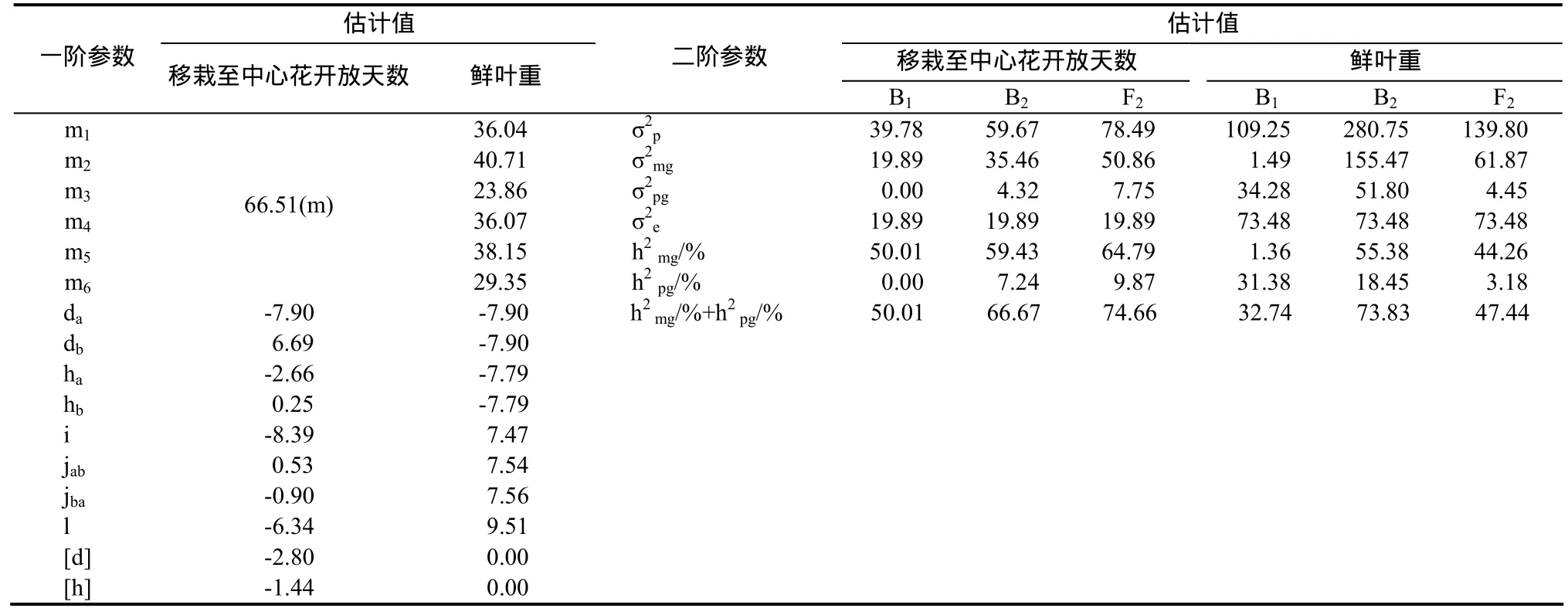
表5 移栽至中心花開放天數的E1模型和鮮葉重的E0模型遺傳參數Table 5 The estimate of genetic parameters of days of transplanting to flowering in E1 model and fresh leaf weight in E0 model
3 討 論
3.1 性狀遺傳率比較
在F2世代中,按照遺傳率大小各植物學性狀排序依次為:葉數、蒴果重、節距、移栽至中心花開放天數、株高、葉形指數、比葉重、葉寬、葉長、莖圍、鮮葉重和葉面積,幾個相關性狀的遺傳率大小、順序與牛佩蘭[8-10]和艾樹理[11]的研究結果基本一致,其中株高結果與王日新[12]的研究結果接近。按照主基因遺傳率大小各性狀排序依次為:移栽至中心花開放天數、株高、葉形指數、鮮葉重、葉數、比葉重、葉面積和莖圍,而葉長、葉寬、節距和蒴果重沒有檢測到主基因的存在。移栽至中心花開放天數、株高、葉形指數、鮮葉重、葉數、比葉重及葉面積的主基因遺傳率較高,適合在早代選擇,而莖圍、葉長、葉寬、節距和蒴果重則宜在晚代選擇。
3.2 性狀基因效應分析
株高遺傳以加性效應及顯性×顯性上位性效應為主,這與肖炳光[13]、盧秀萍[15]的研究結果一致,后者與王日新[12]及Matzinger[2]的研究結果一致;葉數遺傳以加性效應及顯性×顯性上位性效應為主,其中前者與Matzinger[2]、艾樹理[11]、巫升鑫[5]和陳順輝[6]的研究結果一致,后者與牛佩蘭[10]的研究結果不一致,為首次發現;葉面積幾種遺傳效應差不多,其中上位性效應>加性效應>顯性效應;葉形指數主基因加性效應與顯性效應基本相當;莖圍遺傳以多基因為主,其多基因加性效應和顯性效應大小相當,這與許健[14]和牛佩蘭[10]研究結果不一致;移栽至中心花開放天數遺傳以加性×加性上位性效應、加性效應及顯性×顯性上位性效應為主,其中前者與大多數研究者的結果不一致,中者與Matzinger[2]的研究結果一致,后者與Matzinger[2]和Pandeya[3]的研究結果不一致;鮮葉重幾種遺傳效應大致相當,其中上位性效應>加性效應>顯性效應;比葉重遺傳主基因和多基因的加性效應和顯性效應大致相當;而葉長、葉寬、節距和蒴果重沒有檢測到主基因的存在。
本研究還發現,上位性效應是一種非常重要的遺傳效應,如在株高、葉數、葉面積、移栽至中心花開放天數及鮮葉重的雜種優勢形成過程中起到非常重要的作用,一些作用方向相同,而另一些作用則方向相反,說明基因聚合后會產生不同的互作效應。另外在常規選育中,一般認為只有加性效應可以穩定遺傳,選擇育種中得到的可穩定遺傳的變異就是加性效應值。事實上,對于煙草來說,加性×加性上位性效應可以通過選擇穩定遺傳,如果加性×加性上位效應存在,那么在選擇育種中將會產生額外的遺傳收益。在移栽至中心花開放天數中,加性×加性上位效應占據主導地位,可以通過選擇穩定地遺傳雜種優勢。通過上位性效應分析,還可以為育種親本的選擇提供更為詳細的信息。即通過對選育親本的上位性遺傳效應預測,選擇既有較大加性效應,又有較高上位性效應的育種親本,以期獲得更理想的育種效果。
本研究結果與前人研究有相同也有不同,前人研究基本不能像孟德爾方法那樣追蹤個別基因,而只能從一組基因的總體上得到有關遺傳效應及其相對重要的信息。對植物遺傳改良來說,十分需要有關數量性狀遺傳體系以及其中個別重要基因的信息,以便通過育種手段進行遺傳操作。“主基因+多基因”混合遺傳模型的出現,有效地解決了上述問題。
4 結 論
烤煙的株高、葉數、葉面積和鮮葉重均受2對加性-顯性-上位性主基因+加性-顯性-上位性多基因(E0)控制,移栽至中心花開放天數受兩對加性-顯性-上位性主基因+加性-顯性多基因(E1)控制,莖圍和比葉重均受1對完全顯性主基因+加性-顯性多基因(D3)控制,葉形指數受 1對加性-顯性主基因+加性-顯性-上位性多基因(D0)控制,葉長、葉寬、節距和蒴果重均受加性-顯性-上位性多基因(C0)控制,移栽至中心花開放天數、株高、葉形指數、鮮葉重、葉數、比葉重及葉面積的主基因遺傳率較高,適合在早代選擇,而莖圍、葉長、葉寬、節距和蒴果重則宜在晚代選擇。
[1]Gopinath D M, Ramanarao V V, Subrahmanyam M, et al.A study of diallel crosses between varieties of Nicotiana tabacum L.for yield component[J].Euphytica, 1966,15(2): 171-178.
[2]Matzinger D F, Mann T J, Cocherham C C.Diallel crosses in Nicotiana tabacum[J].Crop Science, 1962,2(2): 383-386.
[3]Pandeya R S, Dirks V A, Poushinsky G.Quantitative genetic studies in flue-cured (Nicotiana tabacum).Ⅰ.Agronomic characters[J].Canadian Journal of Genetics and Cytology, 1983, 25(2): 336-345.
[4]Wiklinson C A, Tilson W M.Diallel analysis of crosses among Virginia fire-cured tobacco cultivars[J].Tobacco Science, 1994, 38(1): 21-24.
[5]巫升鑫,潘建菁,陳順輝,等.烤煙若干農藝性狀的雜種優勢及其遺傳分析[J].中國煙草學報,2001,7(4):17-22.
[6]陳順輝,巫升鑫,倪金應,等.烤煙主要數量性狀的配合力研究[J].中國煙草學報,2004,10(3):25-28.
[7]王志德,王元英,牟建民.煙草種質資源描述規范和數據標準[M].北京:中國農業出版社,2006.
[8]牛佩蘭,佟道儒,駱啟章.烤煙幾個主要數量性狀遺傳力的估算[J].中國煙草,1980(1):13-14.
[9]牛佩蘭,劉洪祥,劉偉.烤煙幾個主要數量性狀相關遺傳力的初步研究[J].中國煙草,1984(4):4-5.
[10]牛佩蘭,佟道儒.煙草幾個主要農藝性狀的基因效應分析[J].中國煙草,1989(1):9-12.
[11]艾樹理,王秀蓉.烤煙數量性狀配合力和遺傳力研究[J].煙草科技,1984(3):40-46.
[12]王日新,任民,張興偉,等.普通煙草栽培種內株高性狀主基因加多基因遺傳分析[J].中國煙草科學,2009,30(2):15-20.
[13]肖炳光,朱軍,盧秀萍,等.烤煙主要農藝性狀的遺傳與相關分析[J].遺傳,2006,28(3):317-323.
[14]許健,楊德,張錦偉.烤煙親本配合力的雙列雜交分析[J].煙草科技,2004(l):29-32.
[15]盧秀萍,肖炳光.烤煙株高的發育遺傳研究[J].中國煙草學報,2006,12(4):31-34.
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
核科學與工程(2021年4期)2022-01-12 06:30:26
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
中學物理·高中(2016年12期)2017-04-22 11:53:03
光學精密工程(2016年6期)2016-11-07 09:07:19
核科學與工程(2015年4期)2015-09-26 11:59:03
小櫻桃·童年閱讀(2014年11期)2014-12-01 22:21:30
