三個(gè)煙草防御素基因的克隆及序列分析
2012-07-31 07:22:18林世鋒任學(xué)良王仁剛
中國(guó)煙草科學(xué) 2012年5期
林世鋒,任學(xué)良,王 軼,王仁剛
(貴州省煙草科學(xué)研究所,貴陽(yáng) 550081)
植物防御素(Plant defensins)是一類富含半胱胺酸的低分子量短肽,廣泛分布于植物體內(nèi),參與植物的防衛(wèi)反應(yīng),是極為重要的一種抗病因子[1]。植物防御素因種類和結(jié)構(gòu)的差別具有不同作用,包括抗真菌[2-3]和抗細(xì)菌[4],以及蛋白酶抑制和抗蟲(chóng)作用[5-6],部分研究也發(fā)現(xiàn)防御素可以在環(huán)境脅迫情況下對(duì)植物體起保護(hù)作用[7]。目前,已有不同學(xué)者分別從煙草屬的普通煙草(Nicotiana tabacum)[8]、高煙草(N.excelsior)[9]、圓錐煙草(N.paniculata)、漸狹葉煙草(N.attenuata)[10]和花煙草(N.alata)[11-12]中克隆到防御素基因,其中部分基因的功能已初步明確,如從花煙草中獲得的防御素基因具有抗真菌和抗蟲(chóng)雙重作用。
電子克隆(In silico cloning)是近年來(lái)伴隨著基因組計(jì)劃和 EST計(jì)劃而發(fā)展起來(lái)的基因克隆新方法[13],它依賴于各種生物信息數(shù)據(jù)庫(kù),利用生物軟件拼接延伸EST序列,獲得目的基因的部分乃至全長(zhǎng)的cDNA序列。本研究采用電子克隆的方法獲得煙草栽培品種K326(N.tabacum)的3個(gè)防御素基因cDNA序列并進(jìn)行生物信息學(xué)分析,以期為進(jìn)一步研究該類基因的進(jìn)化關(guān)系和功能差異,比較其在煙草病蟲(chóng)害防御反應(yīng)中的作用奠定基礎(chǔ)。
1 材料與方法
1.1 煙草防御素基因的電子克隆
以辣椒防御素CaDEF1基因的全長(zhǎng)cDNA序列(GenBank登陸號(hào):AF442388)[7]為信息探針,對(duì)GenBank中 EST_others數(shù)據(jù)庫(kù)指定物種煙草(Nicotiana tabacum)進(jìn)行BLAST檢索,將檢索到的來(lái)自于煙草栽培品種K326的全部EST序列利用DNAMAN 6.0軟件進(jìn)行拼接,形成重疊群(contig)。然后用此重疊群再次進(jìn)行同源檢索、拼接,重復(fù)以上過(guò)程直至沒(méi)有更多煙草栽培品種 K326的重疊EST檢出,最終獲得煙草栽培品種 K326防御素cDNA序列片段。再以此片段在 GenBank中進(jìn)行blastx同源搜索,與其他物種的防御素同源蛋白進(jìn)行序列比較,進(jìn)而判斷拼接得到的煙草防御素基因cDNA的正確性以及其ORF序列的完整性。
1.2 煙草防御素基因的生物信息學(xué)分析
利用NCBI網(wǎng)站(http://www.ncbi.nlm.nih.gov/)提供的ORF finder軟件對(duì)開(kāi)放閱讀框進(jìn)行分析并翻譯成蛋白質(zhì),通過(guò)BlastP程序進(jìn)行序列相似性分析和氨基酸保守性預(yù)測(cè);利用瑞士生物信息研究所網(wǎng)站(http://www.expasy.org/tools/)提供的ProtParam、SignalP和NetPhos軟件進(jìn)行蛋白的基本特性、信號(hào)肽和磷酸化位點(diǎn)分析,通過(guò) SWISS-MODEL和CPHmodels在線工具對(duì)蛋白質(zhì)的三級(jí)結(jié)構(gòu)進(jìn)行預(yù)測(cè);利用 http://www.predictprotein.org/網(wǎng)站提供的Predict Protein軟件進(jìn)行蛋白質(zhì)二硫鍵的預(yù)測(cè);運(yùn)用DNAMAN 6.0、ClustalX 2.0和MEGA3.1軟件進(jìn)行氨基酸序列比對(duì)和進(jìn)化樹(shù)分析。
2 結(jié) 果
2.1 煙草防御素基因的電子克隆
利用已知的辣椒防御素基因的核苷酸序列對(duì)GenBank中煙草EST數(shù)據(jù)庫(kù)進(jìn)行BLAST檢索分析,發(fā)現(xiàn) 97條相似性高的 EST序列來(lái)自煙草品種K326。對(duì)它們進(jìn)行重疊群分析,結(jié)果顯示97條EST可拼接成3個(gè)獨(dú)立的cDNA序列,長(zhǎng)度為554 bp、496 bp和434 bp,代表煙草3個(gè)不同的防御素基因,分別命名為NtDEF1、NtDEF2和NtDEF3(Nicotiana tabacumdefensins)。經(jīng)NCBI ORF finder分析,三者分別包括一個(gè)長(zhǎng)度237 bp、234 bp和231 bp的開(kāi)放讀碼框,依次編碼由78、77和76個(gè)氨基酸組成的防御素(圖1)。
2.2 煙草防御素的結(jié)構(gòu)特征分析
將推導(dǎo)出的 3個(gè)煙草防御素氨基酸序列利用SignalP軟件進(jìn)行信號(hào)肽分析,發(fā)現(xiàn) NtDEF1和NtDEF2蛋白氨基端的前20個(gè)氨基酸,NtDEF3蛋白氨基端的前19個(gè)氨基酸為信號(hào)肽(圖1)。植物防御素的信號(hào)肽不僅具有幫助多肽有效靶向分泌的作用,同時(shí)也具有緩解蛋白毒性的作用,只在到達(dá)作用空間需要防御素發(fā)揮毒性作用的時(shí)候才將成熟多肽釋放出來(lái)[14]。
用推測(cè)的氨基酸序列進(jìn)行BLASTp比較,發(fā)現(xiàn)NtDEF1、NtDEF2和NtDEF3均具有γ-硫素蛋白家族(γ-thionin family)結(jié)構(gòu)域(圖1),該序列編碼具有殺菌功能的毒素功能結(jié)構(gòu)[14-15]。
2.3 煙草防御素的基本特性分析
利用ProtParam軟件對(duì)3個(gè)煙草防御素成熟蛋白進(jìn)行基本特性分析,結(jié)果顯示NtDEF1含有原子總數(shù)為855,分子式為C259H415N87O84S10,其分子量為6412.2 Da,理論等電點(diǎn)為8.52。在氨基酸組成上,半胱氨酸和精氨酸相同各有8個(gè),均占13.8%;其次是甘胺酸和蘇氨酸相同各有6個(gè),均占10.3%。總的帶正電荷和總的帶負(fù)電荷的氨基酸殘基數(shù)分別為9和6。預(yù)測(cè)該蛋白不穩(wěn)定指數(shù)為61.06,屬于不穩(wěn)定蛋白。
顯示NtDEF2含有原子總數(shù)為860,分子式為C268H414N90O78S10,其分子量為6 465.3 Da,理論等電點(diǎn)為8.95。在氨基酸組成上,半胱氨酸含量最高有8個(gè),占14.0%;其次是精胺酸7個(gè)占12.3%。總的帶正電荷和總的帶負(fù)電荷的氨基酸殘基數(shù)分別為11和4。預(yù)測(cè)該蛋白不穩(wěn)定指數(shù)為63.59,屬于不穩(wěn)定蛋白。

圖1 煙草防御素1(A)、2(B)和3(C)cDNA序列及推導(dǎo)的氨基酸序列Fig.1 Nucleotide and deduced amino acid sequences of the tobacco defensins
顯示NtDEF3含有原子總數(shù)為879,分子式為C271H430N90O80S8,其分子量為 6485.4 Da,理論等電點(diǎn)為9.22。在氨基酸組成上,半胱氨酸含量最高有8個(gè),占14.0%;其次是甘胺酸和精氨酸相同各有7個(gè),均占12.3%。總的帶正電荷和總的帶負(fù)電荷的氨基酸殘基數(shù)分別為11和4。預(yù)測(cè)該蛋白不穩(wěn)定指數(shù)為28.89,屬于穩(wěn)定蛋白。
總平均親水性 GRAVY(grand average of hydropathicity)分析表明,三者 GRAVY值分別為-0.579、-0.523和-0.705,這表明三者均為親水性蛋白。
2.4 煙草防御素的二硫鍵預(yù)測(cè)
NtDEF1、NtDEF2和NtDEF3都含有8個(gè)半胱氨酸,經(jīng)PredictProtein預(yù)測(cè)軟件分析,發(fā)現(xiàn)8個(gè)半胱氨酸在煙草3種防御素中形成的二硫鍵連接方式一致,均為Cys-1-Cys-8、Cys-2-Cys-5、Cys-3-Cys-6、Cys-4-Cys-7。其中,圖2為NtDEF1二硫鍵預(yù)測(cè)結(jié)果,這與典型的植物防御素二硫鍵結(jié)構(gòu)模型相同[14]。
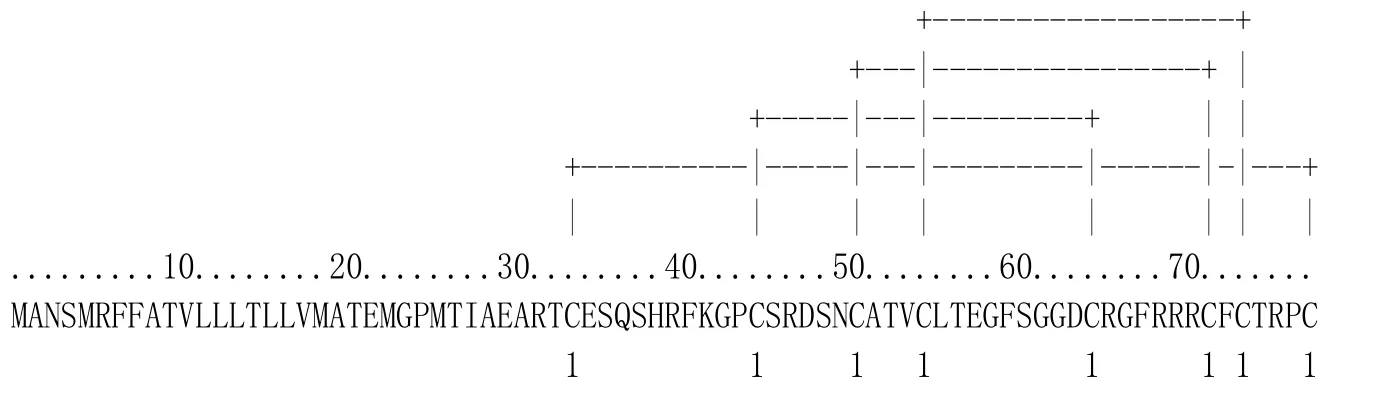
圖2 煙草防御素NtDEF1二硫鍵預(yù)測(cè)Fig.2 Predicted disulifide bridges in NtDEF1
2.5 煙草防御素三維結(jié)構(gòu)預(yù)測(cè)
分別應(yīng)用SWISS-MODEL和CPHmodels在線工具對(duì)3個(gè)煙草防御素進(jìn)行蛋白三維結(jié)構(gòu)預(yù)測(cè),以PDB數(shù)據(jù)庫(kù)中大麥(Hordeum vulgare)防御素(DEF1_HORVU)[16-17]的三維結(jié)構(gòu)(PDB id=1gpt)為模板,對(duì)NtDEF1和NtDEF2進(jìn)行三維結(jié)構(gòu)同源建模,序列一致性為53.2%;以PDB數(shù)據(jù)庫(kù)中小麥(Triticum aestivum)防御素(DEF1_WHEAT)[17-18]的三維結(jié)構(gòu)(PDB id=1gps)為模板,對(duì) NtDEF3進(jìn)行三維結(jié)構(gòu)同源建模,序列一致性為46.8%。預(yù)測(cè)結(jié)果如圖3所示。從圖中可以看出煙草三種防御素蛋白的三維結(jié)構(gòu)都含有三段反向平行的 β-折疊和一段α-螺旋,與己推導(dǎo)出的植物防御素蛋白結(jié)構(gòu)一致。
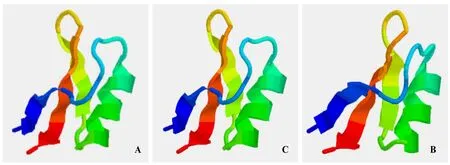
圖3 煙草防御素1(A)、2(B)和3(C)的三級(jí)結(jié)構(gòu)預(yù)測(cè)Fig.3 Tertiary structure prediction of the tobacco defensins
2.6 氨基酸序列比對(duì)和同源性分析
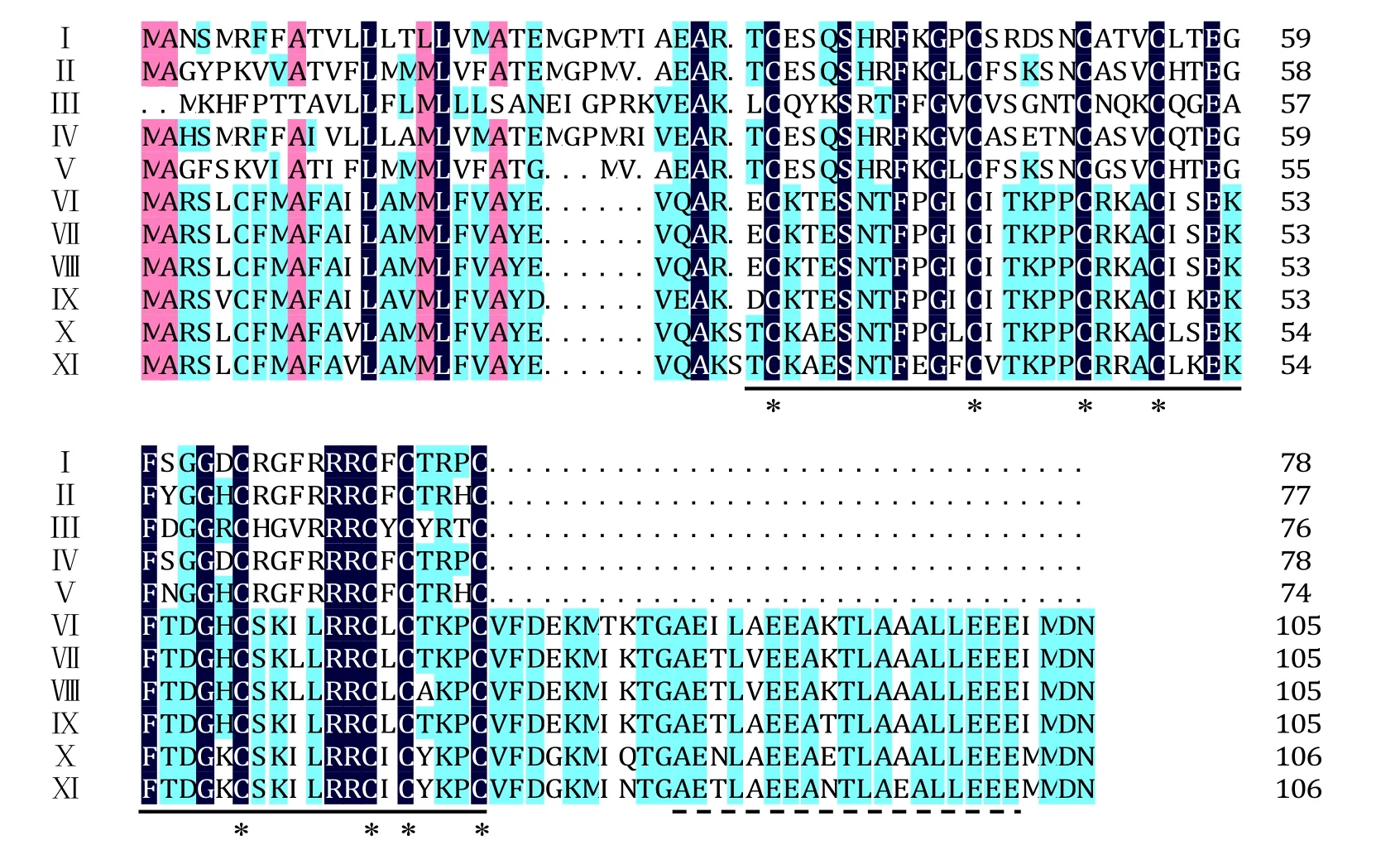
圖4 不同植物防御素基因氨基酸序列同源性比較Fig.4 Alignment of deduced amino acid sequences of defensin genes of different plants
3個(gè)煙草防御素氨基酸序列與部分其他植物進(jìn)行比對(duì),結(jié)果顯示在植物防御素中一些保守的氨基酸在煙草中同樣存在(圖4),例如:植物防御素的典型特征序列即γ-硫素蛋白結(jié)構(gòu)域在煙草中同樣保守,且具有構(gòu)成4個(gè)二硫鍵所需的8個(gè)半胱氨酸殘基。利用DNAMAN軟件對(duì)煙草3種防御素進(jìn)行同源性分析顯示,NtDEF1和NtDEF2的一致性最高為72%,NtDEF1和NtDEF3的一致性最低為40%,NtDEF2和NtDEF3的一致性為47%。與其他物種相比較,三者均與辣椒的一致性最高,在50%~87%。盡管在普通煙草、高煙草、圓錐煙草、漸狹葉煙草及花煙草中已有防御素基因的報(bào)道,但是與其一致性都較低,在 33%~40%,且氨基酸 C-端不具備與其對(duì)應(yīng)的低復(fù)雜性區(qū)域(Low Complexity Region,LCR),表明NtDEF1、NtDEF2和NtDEF3是3個(gè)新的煙草防御素基因。
2.7 系統(tǒng)進(jìn)化樹(shù)分析
從 GenBank上獲得已知的植物防御素的氨基酸序列經(jīng)ClustalX 2.0比對(duì)后,利用Mega 3.1以鄰位相接(NJ)構(gòu)建系統(tǒng)進(jìn)化樹(shù)(圖5)。結(jié)果表明,防御素基因在煙草、辣椒、番茄、擬南芥等植物中均以多種形式存在,且序列差異較大,在系統(tǒng)進(jìn)化樹(shù)中分散在遺傳距離很遠(yuǎn)的分支上。NtDEF1、NtDEF2和NtDEF3處于第Ⅰ分支的不同小分支上,而已報(bào)道的煙草屬防御素基因均處于第Ⅱ分支上,進(jìn)一步證明前者是煙草3種新的防御素基因。

圖5 煙草與其他物種防御素同源蛋白的系統(tǒng)進(jìn)化分析Fig.5 Phylogenetic analysis of amino acid sequences of defensin genes from tobacco and other species
3 討 論
基于表達(dá)序列標(biāo)簽(Expressed Sequence Tags,EST)的電子克隆策略,具有簡(jiǎn)單、快速、針對(duì)性強(qiáng)等優(yōu)點(diǎn),現(xiàn)已成為一種廣泛使用的基因克隆技術(shù)[13]。本研究以煙草與辣椒同屬茄科植物,物種間同源基因序列相對(duì)保守為切入點(diǎn),以辣椒防御素CaDEF1基因cDNA序列作為探針,對(duì)煙草EST數(shù)據(jù)庫(kù)進(jìn)行檢索拼接,成功克隆到目的物種的3個(gè)新防御素基因序列。在電子克隆檢索過(guò)程中,目標(biāo)物種鎖定煙草(Nicotiana tabacum),全部EST序列來(lái)源于同一品種 K326,避免了品種間基因多態(tài)性造成序列拼接工作的復(fù)雜性,進(jìn)而增加了電子克隆的準(zhǔn)確性。同時(shí),K326是我國(guó)主栽煙草品種之一,對(duì)其重要功能基因的研究,勢(shì)必具有重要的理論及應(yīng)用價(jià)值。
研究證實(shí)植物防御素具有抗真菌[2-3]、抗細(xì)菌[4]和蛋白酶抑制劑[5]等生物活性,而 γ-硫素類防御素由于其功能廣泛、分子量小以及穩(wěn)定等特點(diǎn)在植物基因工程中極具應(yīng)有前景[14-15]。γ-硫素蛋白超家族功能區(qū)具有嚴(yán)格保守的8個(gè)半胱氨酸殘基,形成4個(gè)分子內(nèi)二硫鍵以穩(wěn)定三段反向平行的 β-折疊和一段α-螺旋,從而形成具有復(fù)雜三維折疊結(jié)構(gòu)的抗菌短肽[14-15]。盡管本研究克隆到的3個(gè)防御素同樣具備上述保守功能區(qū),但3個(gè)防御素在氨基酸組成上存在一定的區(qū)別,如纈氨酸、賴氨酸、蘇氨酸和苯丙氨酸的數(shù)目差異較顯著。這些氨基酸的差異是否造成功能上的差異還需要進(jìn)一步深入研究。
目前,在番茄、辣椒等茄科植物中已克隆得到多個(gè)防御素cDNA序列,其中辣椒防御素CaDEF1和CaDEF2的氨基酸序列一致性為64%,它們?cè)诟⑶o、葉和果實(shí)中的表達(dá)模式各不相同[7,19]。我們利用本研究克隆的防御素基因進(jìn)行 Blastn同源檢索時(shí),在煙草的EST數(shù)據(jù)庫(kù)中得到了數(shù)目不同且高度同源的煙草品種K326的防御素EST序列,其中以NtDEF1為最高。這預(yù)示著在煙草中盡管存在結(jié)構(gòu)相對(duì)保守的多種防御素,但是其表達(dá)分布或者在同一組織中的表達(dá)量可能各不相同,提示它們可能在不同或相同的組織中發(fā)揮著特異的作用。
[1]Broekaert W F, Terras F R, Cammue B P, et al.Plant defensins: novel antimicrobial peptides as components of the host defense system [J].Plant Physiol, 1995, 108:1353-1358.
[2]Lai F M, DeLong C, Mei K, et al.Analysis of the DRR230 family of pea defensins: gene expression pattern and evidence of broad host-range antifungal activity [J].Plant Sci, 2002, 163: 855-864.
[3]Solis J, Medrano G, Ghislain M.Inhibitory effect of a defensin gene from the Andean crop maca (Lepidium meyenii) againstPhytophthora infestans[J].J Plant Physiol, 2007, 164: 1071-1082.
[4]Choi Y, Choi Y D, Lee J S.Antimicrobial activity of γ-thionin-like soybean SE60 inE.coliand tobacco plants[J].Biochem Biophys Res Commun, 2008, 375: 230-234.
[5]Wijaya R, Neumann G M, Condron R, et al.Defense proteins from seed ofCassia fistulainclude a lipid transfer protein homologue and a protease inhibitory plant defensin [J].Plant Sci, 2000, 159: 243-255.
[6]Chen K C, Lin C Y, Kuan C C, et al.A novel defensin encoded by a mungbean cDNA exhibits insecticidal activity against bruchid [J].J Agric Food Chem, 2002, 50:7258-7263.
[7]Do H M, Lee S C, Jung H W, et al.Differential expression and in situ localization of a pepper defensin(CADEF1) gene in response to pathogen infection,abiotic elicitors and environmental stresses inCapsicum annuum[J].Plant Sci, 2004, 166: 1297-1305.
[8]Gu Q, Kawata E E, Morse M J, et al.A flower-specific cDNA encoding a novel thionin in tobacco [J].Mol Gen Genet, 1992, 234: 89-96.
[9]Yamada S, Komori T, Imaseki H.cDNA cloning of γ-thionin fromNicotiana excelsior(Accession No.AB005266) (PGR97-131) [J].Plant Physiol, 1997, 115:314.
[10]Lou Y, Baldwin I T.Nitrogen supply influences herbivore-induced direct and indirect defenses and transcriptional responses inNicotiana attenuate[J].Plant Physiol, 2004, 135: 496-506.
[11]Lay F T, Brugliera F, Anderson M A.Isolation and properties of floral defensins from ornamental tobacco and petunia [J].Plant Physiol, 2003, 131: 1283-1293.
[12]Lay F T, Schirra H J, Scanlon M J, et al.The three-dimensional solution of NaD1,a new floral defensin fromNicotiana alataand its application to a homology model of the crop defense protein alfAFP [J].J Mol Biol, 2003, 325: 175-188.
[13]Gill R W, Sanseau P.Rapid in silico cloning of genes using expressed sequence tags (ESTs) [J].Biotechnol Annu Rev, 2000, 5: 25-44.
[14]Carvalho A O, Gomes V M.Plant defensins-prospects for the biological functions and biotechnological properties[J].Peptides, 2009, 30: 1007-1020.
[15]Pelegrini P B,F(xiàn)ranco O L.Plant γ-thionins: novel insights on the mechanisms of actions of a multi-functional class of defense proteins [J].Int J Biochem Cell Biol, 2005, 37:2239-2253.
[16]Mendez E, Moreno A, Colilla F, et al.Primary structure and inhibition of protein synthesis in eukaryotic cell-free system of a novel thionin, γ-hordothionin, from barley endosperm [J].Eur J Biochem, 1990, 194: 533-539.
[17]Bruix M, Jimenez M A, Santoro J, et al.Solution structure of gamma 1-H and gamma 1-P thionins from barley and wheat endosperm determined by 1H-NMR: a structural motif common to toxic arthropod proteins [J].Biochemistry, 1993, 32: 715-724.
[18]Colilla F J, Rocher A, Mendez E.γ-Purothionins: amino acid sequence of two polypeptides of a new family of thionins from wheat endosperm [J].FEBS Lett, 1990,270: 191-194.
[19]Meyer B, Houlne G, Pozueta-Romero J, et al.Fruit-specific expression of a defensin-type gene family in bell pepper.Upregulation during ripening and upon wounding [J].Plant Physiol, 1996, 112: 615-622.
猜你喜歡
奧秘(創(chuàng)新大賽)(2023年3期)2023-05-06 01:48:20
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
浙江中西醫(yī)結(jié)合雜志(2017年2期)2017-01-12 18:23:59
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代化工研究(2016年9期)2016-03-20 16:22:08
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·兒童版(2015年1期)2015-07-07 04:12:52
