微生物產(chǎn)3-羥基丙酸研究進(jìn)展
2012-08-17 11:17:41常雁紅劉曉惠北京科技大學(xué)化學(xué)與生物工程學(xué)院北京00083北京科技大學(xué)土木與環(huán)境工程學(xué)院北京00083
化學(xué)與生物工程 2012年11期
關(guān)鍵詞:途徑
程 嫚,羅 暉,常雁紅,劉曉惠(.北京科技大學(xué)化學(xué)與生物工程學(xué)院,北京00083;.北京科技大學(xué)土木與環(huán)境工程學(xué)院,北京00083)
微生物產(chǎn)3-羥基丙酸研究進(jìn)展
程 嫚1,2,羅 暉1,常雁紅2,劉曉惠1
(1.北京科技大學(xué)化學(xué)與生物工程學(xué)院,北京100083;2.北京科技大學(xué)土木與環(huán)境工程學(xué)院,北京100083)
介紹了3-羥基丙酸的性質(zhì)、用途以及研究歷程,討論了采用誘變法和基因工程法對(duì)產(chǎn)3-羥基丙酸菌種進(jìn)行改造的機(jī)理以及產(chǎn)物分離純化等方面的研究進(jìn)展。
3-羥基丙酸;基因工程法;誘變法;分離純化
3-羥基丙酸(3-Hydroxypropionic acid,3-HP),分子式C3H6O3,化學(xué)索引號(hào)503-66-2,為三碳無手性β-羥基羧酸,無色、無味,易溶于水、醇等有機(jī)溶劑[1]。3-HP和乳酸是同分異構(gòu)體,由于羥基位置的不同,3-HP的化學(xué)性質(zhì)更為活潑。3-HP是一種重要的功能性有機(jī)酸,利用已有的成熟催化技術(shù),通過氧化、脫水、酯化反應(yīng)等可以將其轉(zhuǎn)化為多種重要的化學(xué)物質(zhì),如丙烯酸、丙二酸、聚3-HP等,3-HP還可以作為食品或飼料的添加劑和防腐劑。由于3-HP應(yīng)用廣泛,近年來已成為研究熱點(diǎn),美國能源部也將其列為12種最具開發(fā)潛力的化工產(chǎn)品之一[2]。
目前,3-HP的生產(chǎn)方法有化學(xué)法和微生物法,化學(xué)法使用不可再生資源、副產(chǎn)物多、分離困難、污染嚴(yán)重,而微生物法可以有效彌補(bǔ)這些缺點(diǎn),因此,以微生物法生產(chǎn)3-HP成為大勢(shì)所趨。作者在此綜述了微生物產(chǎn)3-HP的研究進(jìn)展。
1 微生物法產(chǎn)3-HP的研究歷程
早在20世紀(jì)70年代,人們就發(fā)現(xiàn)了能夠產(chǎn)3-HP的野生菌株,但普遍產(chǎn)率低、生產(chǎn)周期長,應(yīng)用前景非常有限。隨著基因重組技術(shù)的日益成熟,近年來人們開始研究構(gòu)建基因工程菌來生產(chǎn)3-HP,微生物產(chǎn)3-HP的能力得到明顯提高,部分研究狀況見表1。
2 產(chǎn)3-HP菌種的選育與構(gòu)建
微生物法生產(chǎn)3-HP的關(guān)鍵在于選育高產(chǎn)的菌種,現(xiàn)有的菌種改造方法主要有誘變法和基因工程法。其中誘變法主要是針對(duì)假絲酵母野生菌株,基因工程法主要是構(gòu)建以葡萄糖或甘油為底物的重組菌株。
2.1 誘變法提高微生物產(chǎn)3-HP的能力
在一些自養(yǎng)微生物體內(nèi)[6],3-HP作為一種中間代謝產(chǎn)物存在于某些代謝途徑中,但一般不是微生物生長所必需的物質(zhì),所以在菌體中不能大量積累,其過度積累會(huì)破壞微生物細(xì)胞內(nèi)的代謝平衡。為了提高菌株的3-HP積累量,需要對(duì)菌種進(jìn)行改造,誘變法就是對(duì)產(chǎn)3-HP野生菌株進(jìn)行改造的一種常規(guī)方法。
早在1982年,Hasegawa等[5]分離出一株能夠產(chǎn)3-HP的皺褶假絲酵母,以亞硝基胍為誘變劑,采用以葡萄糖和丙酸分別為唯一碳源的篩選培養(yǎng)基,篩選不能同化丙酸的突變株,最終得到能夠高產(chǎn)β-羥基異丁酸和3-HP的突變體,以葡萄糖和丙酸作為碳源進(jìn)行發(fā)酵,3-HP產(chǎn)量達(dá)到40.0 g·L-1。
近年來,國內(nèi)研究者開始關(guān)注能夠產(chǎn)3-HP的野生菌株,主要以假絲酵母為主。朱建航等[13]分階段控制皺褶假絲酵母突變株發(fā)酵過程的p H值及糖含量等參數(shù),3-HP的最終產(chǎn)量達(dá)到20.0 g·L-1。范煒煒等[9]對(duì)皺褶假絲酵母進(jìn)行誘變育種,3-HP產(chǎn)量達(dá)到8.88 g·L-1。諸葛斌等[19]對(duì)假絲酵母進(jìn)行復(fù)合誘變,在優(yōu)化的培養(yǎng)條件下,3-HP產(chǎn)量達(dá)到30.0 g· L-1。
Hasegawa等[5]提出了皺褶假絲酵母體內(nèi)可能存在的產(chǎn)3-HP的途徑,見圖1。

表1 微生物產(chǎn)3-HP的部分研究狀況Tab.1 Some research of 3-HP production by microorganisms

圖1 皺褶假絲酵母體內(nèi)產(chǎn)3-HP途徑Fig.1 The pathway of producing 3-HP in Candida rugosa
2011年,Zhou等[21]根據(jù)酰基Co A脫氫酶保守序列將皺褶假絲酵母中的丙酰Co A脫氫酶克隆出來,并在Pichia pastoris GS115中表達(dá)出重組蛋白,分析了蛋白的生化性質(zhì),部分證實(shí)了在皺褶假絲酵母中存在如圖1所示的3-HP合成途徑。
在此途徑中,為了提高3-HP的積累量,需要關(guān)注菌株的3-HP降解酶。一般認(rèn)為,降解3-HP的酶為3-羥基丙酸脫氫酶。Hasegawa等[5]所篩選出來的高產(chǎn)3-HP的突變株,其3-羥基丙酸脫氫酶的酶活幾乎為0。Yao等[22]從蠟狀芽孢桿菌中獲得β-羥基異丁酸脫氫酶基因,并在大腸桿菌中表達(dá),發(fā)現(xiàn)該酶能夠使3-HP脫氫,但是,對(duì)于皺褶假絲酵母中存在的β-羥基異丁酸脫氫酶是否也能使3-HP脫氫還需要進(jìn)一步研究證實(shí)。
2.2 構(gòu)建基因工程菌產(chǎn)3-HP
2.2.1 以葡萄糖為底物
葡萄糖是構(gòu)成生物質(zhì)的重要單元,開發(fā)以葡萄糖為底物的產(chǎn)3-HP菌株是以生物質(zhì)為原料基礎(chǔ)的現(xiàn)代生物煉制技術(shù)的重要發(fā)展方向。美國卡吉爾公司在研究由葡萄糖生產(chǎn)3-HP方面做了大量工作,在2002年申請(qǐng)的專利中提出了以葡萄糖為底物產(chǎn)3-HP的途徑,并從不同微生物體內(nèi)獲得途徑所需的酶基因,構(gòu)建了基因工程菌,3-HP產(chǎn)量最高達(dá)到20.0 g·L-1[8]。
到目前為止,根據(jù)微生物體內(nèi)的代謝途徑,學(xué)者共提出了7條由葡萄糖到3-HP的途徑[23](如圖2所示),并分析了每條途徑的能量代謝、輔酶再生等情況。由于產(chǎn)物的合成和輸出細(xì)胞需要ATP,并且需要達(dá)到一定的平衡,而圖2所示的各條途徑生成和消耗的ATP的量不同,因此3-HP的生成量差別很大。其中乳酸途徑的理論轉(zhuǎn)化率很高。該途徑涉及3個(gè)酶:丙酰Co A轉(zhuǎn)移酶(PCT)、乳酰Co A脫水酶(LCD)和3-羥基丙酰Co A脫水酶(OS19)。在此途徑中,由于LCD嚴(yán)格厭氧[24],所以構(gòu)建的基因工程菌需要厭氧發(fā)酵。乳酸作為代謝中間產(chǎn)物和3-HP同時(shí)存在于發(fā)酵液中,由于它們性質(zhì)相似而較難分離,因此,3-HP的 分離純化是此途徑需要解決的一個(gè)難題。
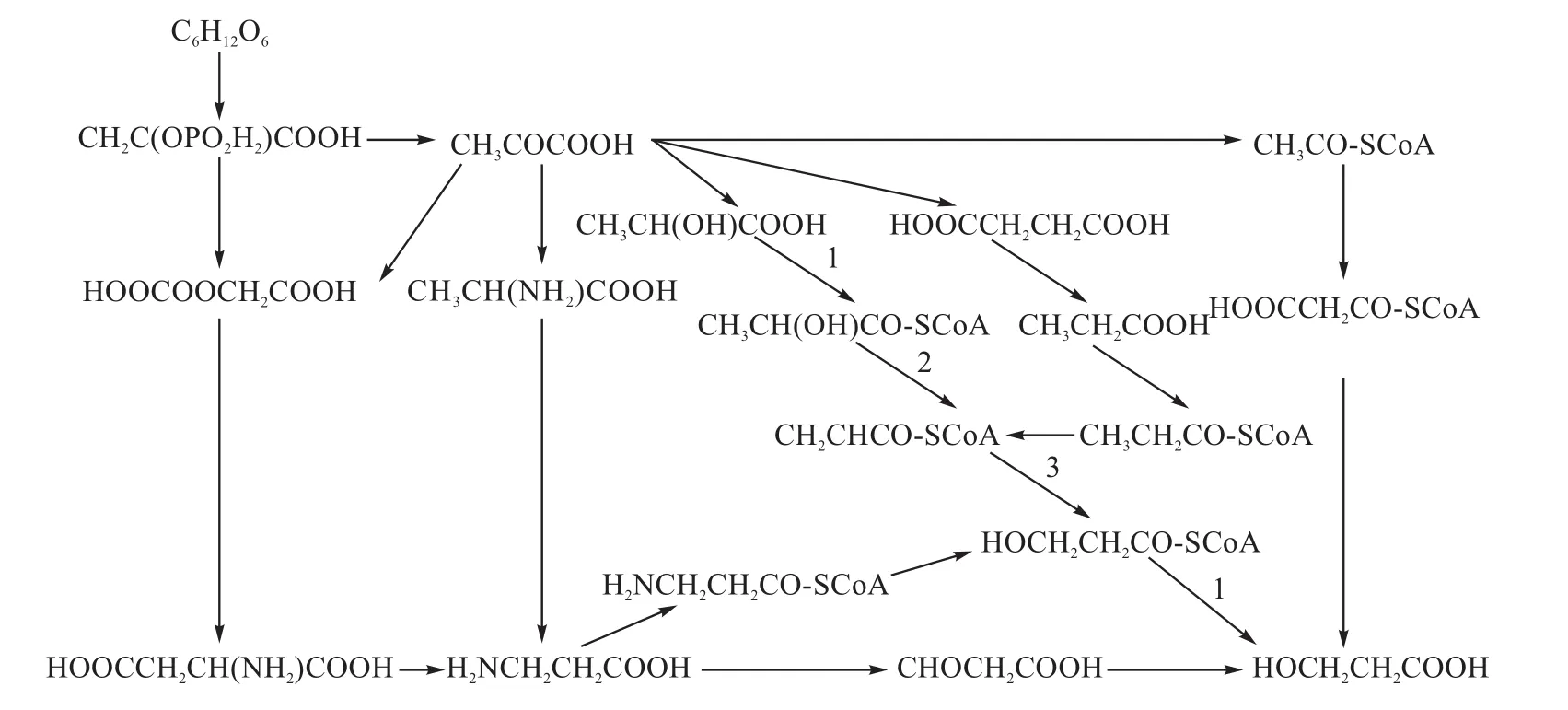
圖2 以葡萄糖為底物產(chǎn)3-HP的代謝途徑Fig.2 The metabolic pathways of producing 3-HP with glucose as substrate
2.2.2 以甘油為底物
21世紀(jì)初,由于生物柴油的大量生產(chǎn),甘油作為一種廉價(jià)的副產(chǎn)物需要加以利用,因此近年來利用甘油生產(chǎn)3-HP的研究得到相當(dāng)?shù)闹匾暋H藗円话阃ㄟ^構(gòu)建基因工程菌來代謝甘油產(chǎn)生3-HP,其主要機(jī)理是:甘油在微生物體內(nèi)發(fā)生氧化還原反應(yīng),氧化反應(yīng)為還原反應(yīng)提供ATP和NAD+[25~27],如圖3所示。

圖3 由甘油產(chǎn)3-HP機(jī)理Fig.3 The mechanism of producing 3-HP from glycerol
由圖3可知,甘油產(chǎn)3-HP需要兩種酶:甘油脫水酶和醛脫氫酶。甘油脫水酶需要以VB12為輔酶催化甘油生成3-羥基丙醛,醛脫氫酶催化3-羥基丙醛(3-HPA)生成3-HP和NADH。甘油脫水酶在催化過程中很不穩(wěn)定,是該途徑的限速酶,甘油會(huì)抑制該酶的活性,從而降低3-HP的產(chǎn)量,解除該抑制作用,就可以有效提高3-HP產(chǎn)量。權(quán)國燕等[18]構(gòu)建了重組大腸桿菌,以甘油脫水酶再激活因子對(duì)甘油脫水酶進(jìn)行激活,3-HP產(chǎn)量提高了6.4倍。王鳳寰等[28]構(gòu)建了重組肺炎克雷伯桿菌轉(zhuǎn)化甘油進(jìn)行3-HP和1,3-丙二醇的共發(fā)酵,不但實(shí)現(xiàn)了NADH的循環(huán)再生,還有效地減少了有毒中間產(chǎn)物3-HPA的積累,3-HP最終產(chǎn)量為5 g·L-1,同時(shí)產(chǎn)生74.5 g·L-1的1,3-丙二醇。Rathnasingh等[14]構(gòu)建了以甘油為底物的重組菌,其3-HP的產(chǎn)量非常可觀,達(dá)到38.7 g·L-1,但是由于酶活丟失及氧化還原反應(yīng)不平衡,轉(zhuǎn)化率未能達(dá)到期望值。因此,還需要進(jìn)一步研究NAD+再生系統(tǒng)以及提高酶活的方法。
由于構(gòu)建基因工程菌涉及到多種基因,并且這些基因在宿主細(xì)胞中的穩(wěn)定性、酶的活性表達(dá)以及細(xì)胞代謝流量調(diào)控等在實(shí)際生產(chǎn)中面臨著巨大挑戰(zhàn),因此目前該技術(shù)尚未實(shí)現(xiàn)工業(yè)化生產(chǎn),仍有許多技術(shù)問題有待解決。
3 3-HP的分離與純化
由于發(fā)酵液成分復(fù)雜,因此3-HP的分離純化是微生物法生產(chǎn)3-HP的一個(gè)難題,但迄今為止,其研究重點(diǎn)仍集中在菌株的構(gòu)建與改造,很少有針對(duì)發(fā)酵液中3-HP分離的文獻(xiàn)報(bào)道。Meng等[29]研究了3-HP和丙烯酸等有機(jī)酸的混合物的分離并申請(qǐng)了專利,以乙酸乙酯或其它有機(jī)物為萃取劑,以逆流萃取的方式分離純化丙烯酸及3-HP,將萃取后的有機(jī)相蒸餾或者用水反萃取,實(shí)現(xiàn)了萃取劑的循環(huán)利用。Jump[30]采用電滲析方法對(duì)3-HP進(jìn)行分離回收,在分離過程中將溶液中的3-羥基丙酸鈉鹽進(jìn)行濃縮并轉(zhuǎn)化為3-HP,該技術(shù)還可以有效地分離溶液中的3-HP和葡萄糖,顯示了良好的應(yīng)用前景。
4 結(jié)語
微生物法生產(chǎn)3-HP以其污染小、副產(chǎn)物少等特點(diǎn)優(yōu)于化學(xué)法,而且構(gòu)建的重組菌利用的是可再生資源,是未來的發(fā)展方向。但迄今為止,微生物發(fā)酵產(chǎn)3-HP的產(chǎn)量最高只有30~40 g·L-1,為了進(jìn)一步提高產(chǎn)量,實(shí)現(xiàn)該方法的技術(shù)和經(jīng)濟(jì)可行性,必須解決微生物法存在的一些問題,如高產(chǎn)物濃度耐受菌株的選育[31]、基因工程菌代謝流的調(diào)控、菌株底物轉(zhuǎn)化率的提高以及產(chǎn)物的高效低成本分離技術(shù)等,這也是科研人員下一步將要重點(diǎn)解決的問題。
[1] 張鴻達(dá),劉成,高衛(wèi)華,等.微生物發(fā)酵法生產(chǎn)3-羥基丙酸的研究進(jìn)展[J].化工進(jìn)展,2007,26(1):33-36.
[2] Werpy T,Petersen G.Top value added chemicals from biomass [C].Washington D.C.:Energy Efficiency and Renewable Energy,2004.
[3] Harada T,Hirabayashi T.Utilization of alcohols by Hansenula miso[J].Agricultural and Biological Chemistry,1968,32(9): 1175-1180.
[4] Miyoshi T,Harada T.Utilization of 2-butyne-1,4-diol by a strain of Fusarium merismoides[J].Journal of Fermentation Technology,1974,52(6):196-199.
[5] Hasegawa J,Ogura M,Kanema H,et al.Production ofβ-hydroxypropionic acid from propionic acid by a Candida rugosa mutant unable to assimilate propionic acid[J].Journal of Fermentation Technology, 1982,60(6):591-594.
[6] Holo H,Sirevag R.Autotrophic growth and CO2 fixation of Chloroflexus aurantiacus[J].Archives of Microbiology,1986,145(2): 173-180.
[7] Suthers P F,Cameron D C.Production of 3-hydroxypropionic acid in recombinant organisms[P].USP 6 852 517 B1,2001-01-19.
[8] Gokarn R R.3-Hydroxypropionic acid and other organic compounds [P].WO 02/42 418 A2,2002-05-30.
[9] 范煒煒,袁其朋.亞硝基胍、紫外誘變篩選高產(chǎn)3-羥基丙酸菌株[J].現(xiàn)代化工,2008,28(Z2):241-246.
[10] Raj S M,Rathnasingh C,Jo J E,et al.Production of 3-hydroxypropionic acid from glycerol by a novel recombinant Escherichia coli BL21 strain[J].Process Biochemistry,2008,43(12):1440-1446.
[11] 黃瑞.利用甘油發(fā)酵生產(chǎn)3-羥基丙酸基因工程菌的構(gòu)建[D].無錫:江南大學(xué),2008.
[12] Lee S H,Park S J,Park O J,et al.Production of 3-hydroxypropionic acid from acrylic acid by newly isolated Rhodococcus erythropolis LG12[J].Journal of Microbiology and Biotechnology, 2009,19(5):474-481.
[13] 朱建航,辜旭輝,劉寅.一種發(fā)酵生產(chǎn)3-羥基丙酸的方法[P].CN 101 457 239,2009-06-17.
[14] Rathnasingh C,Raj S M,Jo J E,et al.Development and evaluation of efficient recombinant Escherichia coli strains for the production of 3-hydroxypropionic acid from glycerol[J].Biotechnology and Bioengineering,2009,104(4):729-739.
[15] Zhu J G,Ji X J,Huang H,et al.Production of 3-hydroxypropionic acid by recombinant Klebsiella pneumoniae based on aeration and ORP controlled strategy[J].Korean Journal Chemical Engineering,2009,26(6):1679-1685.
[16] 李冰,裴疆森.3-羥基丙酸產(chǎn)生菌株的篩選及鑒定[J].食品與發(fā)酵工業(yè),2010,36(4):28-31.
[17] Luo L H,Seo J W,Baek J O,et al.Identification and characterization of the propanediol utilization protein PduP of Lactobacillus reuteri for 3-hydroxypropionic acid production from glycerol [J].Appl Microbiol Biotechnol,2011,89(3):697-703.
[18] 權(quán)國燕,方慧英,諸葛斌,等.甘油脫水酶再激活因子提高重組大腸桿菌3-羥基丙酸合成能力[J].中國生物工程雜志,2011,31 (6):75-80.
[19] 諸葛斌,范俊英,方慧英,等.一種3-羥基丙酸高產(chǎn)菌株及其應(yīng)用[P].CN 102 382 778A,2012-03-21.
[20] 胡南,繆鑫昕,談曙明,等.重組大腸桿菌生物轉(zhuǎn)化甘油生產(chǎn)3-羥基丙酸[J].生物技術(shù),2012,22(1):72-75.
[21] Zhou F L,Zhang Y G,Zhang R B,et al.Expression and characterization of a novel propionyl-Co A dehydrogenase gene from Candida rugosa in Pichia pastoris[J].Appl Biochem Biotechnol,2011,165(7-8):1770-1778.
[22] Yao T R,Xu L,Ying H J,et al.The catalytic property of 3-hydroxyisobutyrate dehydrogenase from Bacillus cereus on 3-hydroxypropionate[J].Applied Biochemistry and Biotechnology, 2010,160(3):694-703.
[23] Jiang X L,Meng X,Xian M.Biosythetic pathways for 3-hydroxypropionic a cid production[J].Microbiol Biotechnol,2009,82(6):995-1003.
[24] Kuchta R D,Hanson G R,Holmquist B,et al.Fe-S Centers in lactyl-Co A dehydratase[J].Biochemistry,1986,25(23):7301-7307.
[25] Cecil W F.Production of 1,3-propanediol from glycerol by Clostridium[J].Applied Environmental Microbiology,1987,53(4): 639-643.
[26] Himmi E H,Bories A,Boussaid A,et al.Propionic acid fermentation of glycerol and glucose by Propionibacterium[J].Applied Microbiology Biotechnology,2000,53(4):435-440.
[27] Biebl H.Fermentation of glycerol to 1,3-propanediol and 2,3-butanediol[J].Applied Microbiol Biotechnology,1998,50(1):24-29.
[28] 王鳳寰,楊建國,王曉楠,等.利用甘油發(fā)酵耦聯(lián)生產(chǎn)3-羥基丙酸及1,3-丙二醇重組菌的構(gòu)建及篩選[J].微生物學(xué)通報(bào),2011,38 (11):1611-1617.
[29] Meng X S,Tsobanakis P,Malsam J,et al.Process for separating and recovering 3-hydroxypropionic acid and acrylic acid[P].USP 7 279 598,2006-09-13.
[30] Jump J.Process for separating and recovering 3-hydroxypropionic acid[P].WO 2011 002 892 A1,2011-01-06.
[31] Warnecke T E,Lynch M D,Lipscomb M L,et al.Identification of a 21 amino acid peptide conferring 3-hydroxypropionic acid stress-tolerance to Escherichia coli[J].Biotechnology and Bioengineering,2012,109(5):1347-1352.
Research Progress of Producing 3-Hydroxypropionic Acid by Microorganisms
CHENG Man1,2,LUO Hui1,CHANG Yan-hong2,LIU Xiao-hui1
(1.School of Chemistry and Biological Engineering,University of Science and Technology Beijing, Beijing 100083,China;2.School of Civil and Environmental Engineering,University of Science and Technology Beijing,Beijing 100083,China)
As a kind of important chemical intermediate,3-hydroxypropionic acid has gained more and more attention by researchers.In this paper,the properties,applications and developing process of 3-hydroxypropionic acid were introduced.The research progress of the mechanisms of the strains producing 3-hydroxypropionic acid by means of genetic engineering method and traditional mutagenesis method,and the product separation and purification were mainly reviewed.
3-hydroxypropionic acid;genetic engineering method;mutagenesis method;separation and purification
TQ 921.7 Q 819
A
1672-5425(2012)11-0009-004
10.3969/j.issn.1672-5425.2012.11.003
2012-07-15
程嫚(1986-),女,河南蘭考人,碩士研究生,研究方向:生物化工,E-mail:chengman-123@163.com;通訊作者:羅暉,副教授,E-mail:luohui@ustb.edu.cn。
猜你喜歡
語數(shù)外學(xué)習(xí)·高中版中旬(2023年2期)2023-05-10 13:26:53
語數(shù)外學(xué)習(xí)·高中版中旬(2022年5期)2022-07-13 20:47:51
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年10期)2019-11-25 07:33:58
中學(xué)生數(shù)理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中學(xué)生百科·大語文(2017年10期)2017-11-04 06:56:38
中國衛(wèi)生(2016年3期)2016-11-12 13:23:26
公民與法治(2016年22期)2016-05-17 04:20:13
中國衛(wèi)生(2014年12期)2014-11-12 13:12:52
癌變·畸變·突變(2014年6期)2014-02-27 06:15:03
