外源油菜素內(nèi)酯對番茄銅脅迫的緩解效應(yīng)
2012-08-31 02:15:20王秀峰姜春輝李曉云崔秀敏
植物營養(yǎng)與肥料學(xué)報 2012年1期
關(guān)鍵詞:植物
尹 博,王秀峰,姜春輝,李曉云,崔秀敏*
(1山東農(nóng)業(yè)大學(xué)資源與環(huán)境學(xué)院,山東泰安271018;2山東農(nóng)業(yè)大學(xué)園藝科學(xué)與工程學(xué)院,山東泰安271018)
銅是植物正常生命活動所必需的微量礦質(zhì)元素,廣泛參與各種生命活動。但植物對銅的需求很低,稍過量就會對植物造成傷害,大部分作物以土壤有效銅含量低于5 mg/kg為缺乏臨界值。銅過量會導(dǎo)致一系列生物代謝過程的紊亂,并最終抑制植物生長而致作物減產(chǎn)[1]。目前銅礦的開采和冶煉廠三廢的排放、含銅農(nóng)業(yè)化學(xué)物質(zhì)(殺真菌劑、殺蟲劑和化肥)和有機肥(高銅豬糞、雞糞和廄肥)的施用可使農(nóng)田土壤,特別是溫室土壤銅含量達到原始土壤的幾倍乃至幾十倍[2-3],溫室土壤重金屬污染變得日益嚴(yán)峻[4-5]。因此,研究重金屬毒害的緩解機理已成為農(nóng)業(yè)可持續(xù)發(fā)展中亟待解決的問題。
表油菜素內(nèi)酯(Epibrassinolide,簡稱 EBR),是一類具有高活性的甾醇內(nèi)酯類結(jié)晶物,是植物正常生長和發(fā)育所必需的,被公認(rèn)為第六大類植物激素,具有促進作物生長,提高含糖量,增加產(chǎn)量和增強對寒、旱、澇、鹽等多種逆境抵抗能力的作用。近年來,一些研究表明,油菜素內(nèi)酯能夠提高植物對鋁、鎳等重金屬的抗性[6-7],但對重金屬銅脅迫的效應(yīng)還未見報道。
本文主要以番茄為試驗材料,研究外源EBR對銅脅迫下番茄的緩解效應(yīng),為利用常規(guī)作物進行生物修復(fù)治理重金屬污染提供一定的科學(xué)依據(jù)。
1 材料和方法
1.1 供試材料
供試番茄為“改良毛粉802F1”。Hoagland營養(yǎng)液組 成:Ca(NO3)2·4H2O、KNO3、NH4NO3、KH2PO3、MgSO4、鐵鹽、微量元素,以上試劑均為分析純,用蒸餾水配制適宜濃度待用。0.1 mg/L EBR:準(zhǔn)確稱取一定量的EBR,用少量無水乙醇溶解,用蒸餾水定容到一定體積,4℃保存待用。
1.2 試驗設(shè)計
種子經(jīng)55℃溫湯浸種消毒15 min,然后在潤濕的吸水紙上26℃催芽。待種子露白后,播于洗凈的蛭石中,萌發(fā)后用1/4 Hoagland營養(yǎng)液澆灌。當(dāng)幼苗長出3~4片真葉時,挑選生長一致的植株洗凈根系,移栽于4L塑料桶中,用厚度為3 cm的泡沫塑料板做成圓形蓋子,覆蓋在塑料桶頂部,每盆栽4株,先用1/2 Hoagland營養(yǎng)液培,1周后換成完全營養(yǎng)液,此后每3d更換一次營養(yǎng)液。營養(yǎng)液栽培期間用電動氣泵24 h連續(xù)通氣,待番茄植株長至5~6片真葉時進行銅脅迫處理,處理期間,對相應(yīng)處理每天均勻細(xì)霧噴施0.1 mg/L的EBR溶液(以葉面滴水為止),對照噴等量清水。試驗用水全部為蒸餾水。
試驗設(shè)4個處理:1)完全營養(yǎng)液(CK);2)50 μmol/L CuCl2,銅脅迫處理 (CK+Cu);3)50 μmol/L CuCl2+0.1 mg/L EBR,油菜素內(nèi)酯緩解處理(CK+Cu+EBR);4)0.1 mg/L EBR,油菜素內(nèi)酯對照(CK+EBR)。每個處理三次重復(fù),溫室內(nèi)隨機排列。用低濃度的KOH或者HCl調(diào)節(jié)pH至5.5±0.2,處理8 d后,分別收獲各處理植株,分成不同部位,液氮速凍,置于-80℃左右貯存?zhèn)錅y。
1.3 測定項目與方法
葉綠素含量的測定用80%丙酮提取法;根系活力用 TTC 法測定[8]。
抗氧化酶系比活力測定的樣品制備,取0.5g葉片加3 mL 50 mmol/L PBS緩沖液(含2%PVP,0.2 mmol/L EDTA,提取液中加入5 mmol/L ASA),于冰浴中研磨提取,勻漿液于12000 r/min下低溫離心20 min。上清液用于酶比活力和丙二醛含量測定。
植物葉片和根系中 Cu、Fe、Mn、Zn含量的測定采用HNO3-HClO4消解—原子吸收分光光度法。
1.4 數(shù)據(jù)處理
采用Microsoft Excel軟件對數(shù)據(jù)進行處理及繪圖,采用DPS統(tǒng)計軟件對平均數(shù)進行多重比較。
2 結(jié)果與分析
2.1 外源EBR對銅脅迫下抗氧化酶系統(tǒng)的影響
2.1.1 外源EBR對銅脅迫下SOD比活力的影響
超氧化物歧化酶(SOD)是植物體內(nèi)以氧自由基為底物的酶,在活性氧代謝中處于重要地位,可淬滅超氧負(fù)離子的毒性,終止由超氧負(fù)離子啟動的一系列自由基連鎖反應(yīng)所造成的生物膜損傷。圖1表明,單獨銅脅迫處理使番茄葉片和根系SOD比活力分別比對照降低了68.1%和86.6%,差異達極顯著水平;與CK+Cu處理相比,CK+Cu+EBR處理的番茄葉片和根系SOD比活力顯著升高,根系達到銅脅迫處理的2.8倍,而葉片提高了1倍。這表明外源EBR能夠顯著緩解銅脅迫對SOD比活力的抑制。正常營養(yǎng)液培養(yǎng)條件下,外施EBR降低了葉片SOD的比活力,對根系SOD比活力影響不顯著。
2.1.2 外源EBR對銅脅迫下番茄POD比活力的影響 過氧化物酶(POD)廣泛存在于植物體不同組織中,能夠反映植物生長發(fā)育的特點、體內(nèi)代謝狀況以及對外界環(huán)境的適應(yīng)性。POD活性的升高是植物應(yīng)對環(huán)境脅迫的一種適應(yīng)性反應(yīng)。一定濃度的重金屬脅迫能夠誘導(dǎo)植物組織中POD活性升高,這是植物對所有非生物脅迫的共同響應(yīng)[14]。POD活性的增加可能是由于重金屬脅迫誘導(dǎo)產(chǎn)生了一些對植物體有害的過氧化物,POD利用H2O2催化對自身有害的過氧化物(POD底物)的氧化和分解,POD活性隨著重金屬毒害的增加而逐漸增加[15-16]。圖1還看出,番茄葉片與根系POD比活力對銅脅迫的響應(yīng)趨勢不盡一致。與對照相比,CK+Cu處理使番茄葉片POD比活力降低了10%,而使根系POD比活力提高了81.5%,達顯著水平;而CK+Cu+EBR與CK+Cu處理相比,葉片中POD比活力提高了27.6%,根系POD比活力降低了14.6%。對正常供應(yīng)營養(yǎng)液的番茄而言,外源EBR顯著提高了葉片POD的比活力,對根系POD的比活力影響不顯著。
2.1.3 外源EBR對銅脅迫下番茄CAT比活力的影響 過氧化氫酶(CAT)主要存在于植物過氧化物酶體與乙醛酸循環(huán)體中,是清除H2O2的主要酶類之一。過氧化氫酶可催化如下反應(yīng):2H2O2→O2+2H2O,但對H2O2的親和力較弱。酶的亞細(xì)胞分布表明,過氧化氫酶主要清除光呼吸中產(chǎn)生的H2O2,但因為H2O2可以穿過膜擴散,而非嚴(yán)格的區(qū)域化。研究表明,CAT是C3植物中H2O2清除的關(guān)鍵酶,而且是C3植物耐受脅迫所必需的。圖1顯示,與對照相比,番茄葉片和根系CAT比活力在單獨銅脅迫處理下分別降低了53.3%、36.4%;而CK+Cu+EBR處理的番茄葉片和根系CAT比活力比CK+Cu處理有顯著升高,葉片提高 39.5%,根系提高41.6%,均達到顯著水平。正常營養(yǎng)液培養(yǎng)的番茄噴施外源EBR后對葉片和根系CAT比活力沒有顯著影響。
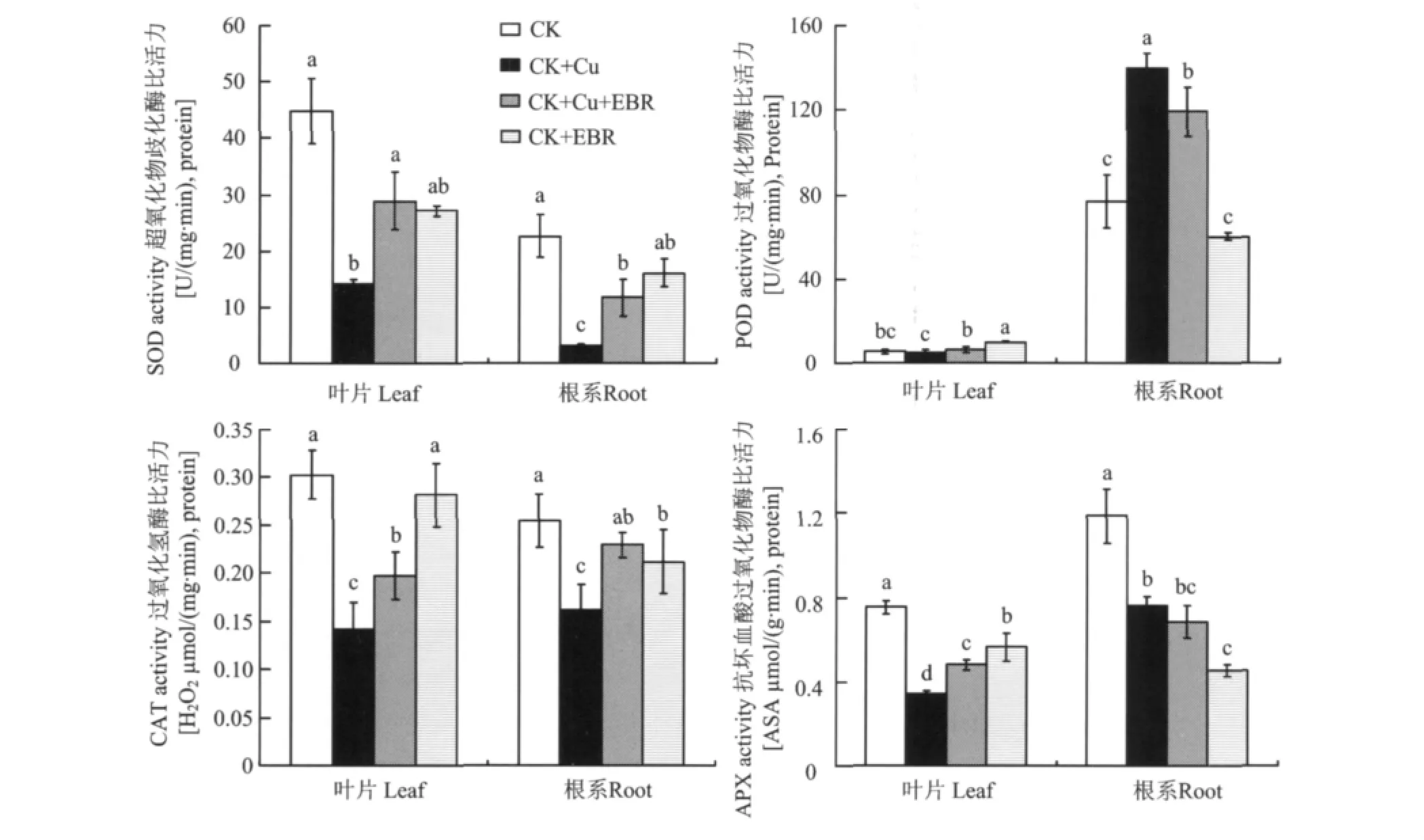
圖1 外源EBR對銅脅迫下番茄葉片和根系SOD、POD、CAT和APX比活力的影響Fig.1 Effects of the exogenous EBR on SOD,POD,CAT and APX activities in tomato leaves and roots under the copper stress
2.1.4 外源EBR對銅脅迫下番茄APX比活力的影響 抗壞血酸過氧化物(APX)酶位于細(xì)胞質(zhì)中或與細(xì)胞壁結(jié)合,與其它氧化還原反應(yīng)相偶聯(lián)起到末端氧化酶的作用,能催化抗壞血酸的氧化,具有抗衰老等作用,其對H2O2的親和力遠(yuǎn)遠(yuǎn)大于CAT,在植物體的物質(zhì)代謝中具有重要的作用。與對照相比,單獨銅脅迫處理分別使番茄葉片APX比活力降低54.9%,根系A(chǔ)PX比活力降低36%;而噴施EBR后,與CK+Cu處理相比,番茄葉片APX比活力提高41.4%,根系A(chǔ)PX比活力降低10%。正常營養(yǎng)液培養(yǎng)的番茄幼苗噴施外源EBR對葉片和根系A(chǔ)PX比活力有一定的抑制作用(圖1)。
第二,實現(xiàn)企業(yè)管理會計與全面預(yù)算管理整合,能夠在企業(yè)內(nèi)部進行動態(tài)預(yù)算管理,及時準(zhǔn)確的掌握預(yù)算有關(guān)的動態(tài)信息,并結(jié)合企業(yè)經(jīng)營管理的實際情況,對預(yù)算進行調(diào)整完善,以進一步強化對企業(yè)整體管控的效果。
2.2 外源EBR對銅脅迫下番茄MDA含量的影響
丙二醛(MDA)是植物受到逆境脅迫時的指示指標(biāo),MDA含量越高說明植物受到的脅迫程度越高。圖2表明,與對照相比,單獨銅脅迫分別提高了番茄葉片和根系的MDA含量,提高幅度為17.4%和34.1%;與CK+Cu處理相比,CK+Cu+EBR處理的植株葉片和根系MDA含量分別降低了12.5%和36.6%,幾乎恢復(fù)到CK處量的水平。正常營養(yǎng)液培養(yǎng)的番茄噴施外源EBR對葉片和根系MDA含量沒有明顯影響。
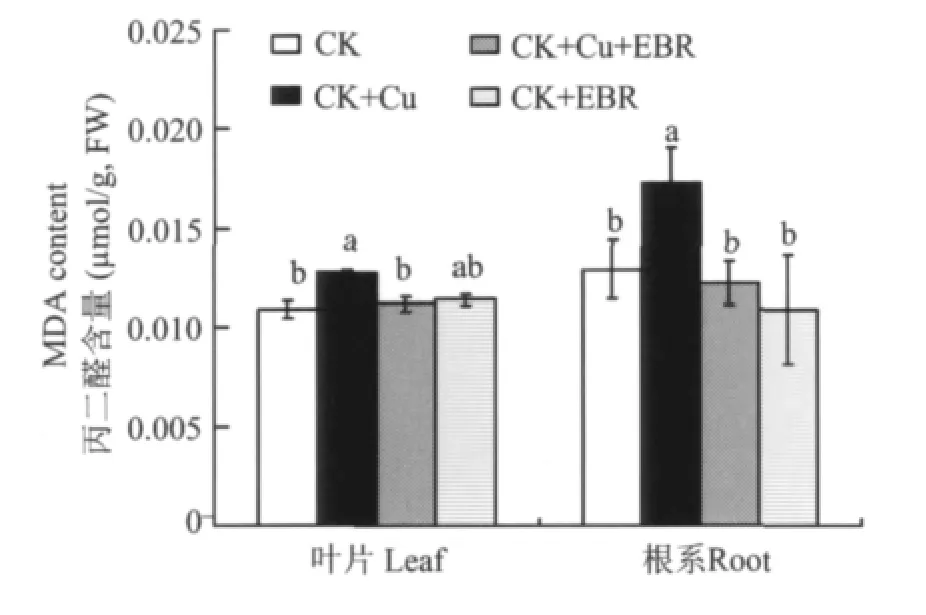
圖2 外源EBR對銅脅迫下番茄葉片和根系MDA含量的影響Fig.2 Effects of the exogenous EBR on MDA contents of tomato leaves and roots under the copper stress
2.3 外源EBR對銅脅迫下番茄銅、鐵、鋅、錳吸收的影響
銅是植物正常生長所必需的,廣泛參與植物的各種生理代謝活動,是細(xì)胞色素氧化酶、多酚氧化酶、抗壞血酸氧化酶、多胺氧化酶、Cu-Zn-SOD等蛋白質(zhì)的重要輔因子,主要參與呼吸代謝中的氧化還原反應(yīng)。然而過量銅能抑制光合作用暗反應(yīng)中的幾個關(guān)鍵酶Rubsic和PEPC活性,損傷光合細(xì)胞器、破壞類囊體結(jié)構(gòu)、抑制光合鏈電子傳遞,還導(dǎo)致植株葉片鐵元素含量的大幅度下降。
圖3表明,銅脅迫下番茄葉片和根系銅的吸收顯著增加,與對照相比,CK+Cu處理番茄葉片和根系銅含量分別提高41.5%和3.4倍。而CK+Cu+EBR比CK+Cu處理明顯降低番茄對銅的吸收與轉(zhuǎn)運,番茄葉片銅含量降低15.4%,根系降低36.3%,差異顯著;而單獨噴施EBR對植株銅含量的影響不顯著。
銅脅迫顯著影響番茄對帶有同等電荷的鐵、鋅、錳的吸收與轉(zhuǎn)運。與對照相比,CK+Cu處理的番茄植株葉片鐵、鋅、錳元素的含量分別降低了12%、21.2%、13.2%;根系鐵、鋅含量分別升高97.3%、25.3%,而錳含量降低48.2%,差異顯著;與CK+Cu處理相比,CK+Cu+EBR處理番茄葉片鐵、鋅、錳含量分別提高了10%、23.3%、15.9%;根系鐵、鋅、錳含量則分別降低了22.6%、31.5%、40%,均達到顯著水平。
2.4 外源EBR對銅脅迫下番茄葉綠素和根系活力的影響
葉綠素作為植物進行光合作用的主要色素,其含量高低可以直接反映光合作用能力的強弱。過量銅會使植株銅元素含量上升而鐵元素含量的大幅下降,植株葉片鐵缺乏引起細(xì)胞代謝紊亂,從而使葉綠素的合成受到抑制。
圖4表明,與對照相比,銅脅迫處理番茄葉片葉綠素含量降低了41.4%,而外源EBR能在一定程度上緩解此傷害。與CK+Cu處理相比,CK+Cu+EBR處理的植株葉綠素含量提高32.2%,單獨施EBR對葉綠素含量有所提高,但差異不顯著。就根系活力來說,與對照相比,單獨銅脅迫處理使番茄根系活力降低了56.4%,而噴施外源EBR后,CK+Cu+EBR處理的番茄植株根系活力比CK+Cu處理提高了92.7%。
2.5 外源EBR對生物量的影響
圖5可以看出,銅脅迫對番茄幼苗生物量有顯著影響,與對照相比,單獨銅脅迫處理單株生物量下降了32%;而CK+Cu+EBR處理則顯著緩解了這種抑制,番茄幼苗的單株重量提高了20.9%,達顯著水平,但沒有恢復(fù)到對照水平。單獨噴施EBR反而抑制了番茄的生長。
3 討論與結(jié)論
銅脅迫能夠誘發(fā)膜脂過氧化、膜透性上升、離子大量外滲,且MDA大量積累,根系活力受到抑制。正常情況下,植物自身能通過抗氧化酶系統(tǒng)清除自身產(chǎn)生的活性氧,緩解其對植物自身的傷害;其中SOD作為植物抗氧化系統(tǒng)的第一道防線,清除細(xì)胞中多余的超氧陰離子,POD、CAT、APX也是清除過氧化物和H2O2的主要酶類。本研究銅脅迫下SOD、CAT、APX等的比活力明顯受到抑制,而外加EBR后其比活力顯著升高,并且MDA含量顯著下降,說明銅脅迫下EBR一定程度上可以通過調(diào)控抗氧化酶的活性來緩解銅脅迫。而銅脅迫下根系POD的比活力明顯提高,施入EBR后POD比活力下降,但仍比對照高,原因可能是EBR作為生物活性分子激活了POD的活性。這與王友保和劉登義報道的銅脅迫在小麥上的效應(yīng)一致[17],可能是由于銅脅迫通過一系列生理生化反應(yīng),使POD底物濃度增加、POD比活力被刺激所致;銅脅迫后SOD、POD、CAT、APX的變化趨勢并非完全一致,這些現(xiàn)象可能與不同氧化酶類對銅脅迫處理下銅的不同適應(yīng)時間和濃度有關(guān)系。
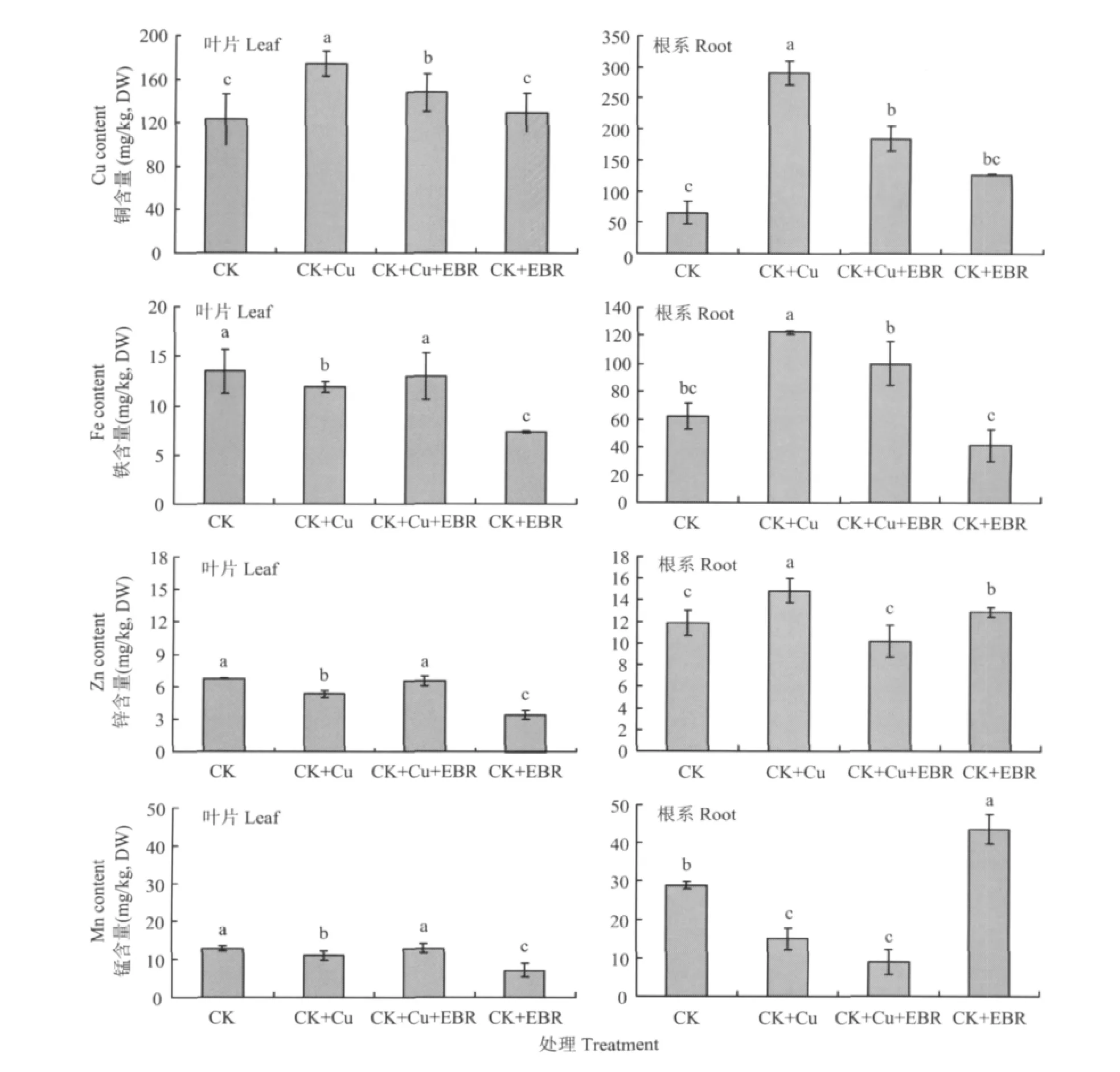
圖3 外源EBR對銅脅迫下番茄葉片和根系銅、鐵、鋅、錳含量的影響Fig.3 Effects of the exogenous EBR on Cu,F(xiàn)e,Zn and Mn contents of tomato leaves and roots under the copper stress
葉綠素作為主要的光合色素,參與光能的吸收、傳遞和轉(zhuǎn)化。銅脅迫下番茄葉綠素含量顯著下降。一方面,銅進入植物體內(nèi)可使葉綠素酶活性比例失調(diào),致使葉片葉綠素分解加快;另一方面,由于銅局部積累過多,與葉綠素蛋白質(zhì)的-SH結(jié)合或取代其中鐵、鋅、錳等,使得葉綠素蛋白中心離子組成發(fā)生變化,破壞了葉綠素的結(jié)構(gòu)和功能。此外,高濃度銅脅迫通過抑制根系生長,使根系的養(yǎng)分吸收能力下降,造成鐵缺乏,也會導(dǎo)致葉綠素合成降低。而外加EBR后,葉綠素的含量有明顯提高,原因一方面可能是EBR激活了在銅脅迫下的葉綠素合成過程中的某些酶的活性;另一方面也可能是EBR阻止了銅與葉綠素的接觸;外源EBR提高了番茄銅脅迫下的根系活力,增加了葉片鐵、鋅等元素的含量,在一定程度上也減緩了對葉綠素的破壞。

圖4 外源EBR對銅脅迫下番茄葉綠素含量和根系活力的影響Fig.4 Effects of the exogenous EBR on the chlorophyll contents and root activities of tomato under the copper stress

圖5 外源EBR對銅脅迫下番茄生物量的影響Fig.5 Effects of the exogenous EBR on the plant weight of tomato under the copper stress
適應(yīng)植物體正常生長的銅含量范圍很窄,稍微過量的銅便會干擾細(xì)胞代謝和離子平衡對植物產(chǎn)生毒害作用。Patsikka等認(rèn)為[18],鈾脅迫提高光系統(tǒng)Ⅱ(PSⅡ)對光的敏感性,原因是過量銅處理使鐵含量大幅度下降,植株葉片鐵缺乏引起細(xì)胞代謝紊亂,葉綠素合成受到抑制。由圖3可以看出,50 μmol/L銅處理一周后番茄銅含量顯著升高,鐵含量顯著降低,而外施EBR能有效地降低番茄銅含量,葉片中鐵含量幾乎恢復(fù)到正常水平。外源EBR同樣對銅脅迫下番茄葉片鋅和錳的含量有調(diào)節(jié)作用,鋅、錳含量幾乎恢復(fù)到了正常水平;而銅脅迫下根系中鐵、鋅的吸收與葉片趨勢相反,與銅脅迫處理相比,施加外源EBR顯著降低了根系鐵、鋅含量,對錳的影響不顯著,這與以往研究結(jié)果不太一致[19],有待進一步探索。
從抗氧化酶活性、礦質(zhì)元素、根系活性、生物量等指標(biāo)看,同正常營養(yǎng)液處理相比,外源EBR在銅脅迫逆境條件下更能發(fā)揮作用,說明外施EBR在銅脅迫的緩解方面有顯著的作用。
[1]Wojcik M,Tukiendorf A.Response of wild type ofArabidopsis thalianato copper stress[J].Biol.Plantarum,2003,46(1):79-84.
[2]Brun L A,Maillet J,Hinsinger Pet al.Evaluation of copper availability to plants in copper-contaminated vineyard soils[J].Environ.Pollut.,2001,111:293 -302.
[3]Kaplan M.Accumulation of Cu in soils and leaves of tomato plants in greenhouses in Turkey[J].Plant Nutr.,1999,22:237 -244.
[4]黃治平,徐斌,張克強,楊秀春.連續(xù)四年施用規(guī)模化豬場豬糞溫室土壤重金屬積累研究[J].農(nóng)業(yè)工程學(xué)報,2007,23(11):239-243.
Huang Z P,Xu B,Zhang K Q,Yang X C.Accumulation of heavy metals in the four years’continual swine manure-applied greenhouse soils [J].Trans.Chin.Soc.Agric.Eng.,2007,23(11):239-243.
[5]曾希柏,李蓮芳,白玲玉,等.山東壽光農(nóng)業(yè)利用方式對土壤砷累積的影響[J].應(yīng)用生態(tài)學(xué)報,2007,18(2):310-316.
Zeng X B,Li L F,Bai L Y.Arsenic accumulation in different agricultural soils in Shouguang of Shandong Province[J].Chin.J.Appl.Ecol.,2007,18(2):310 -316.
[6]Abdullahi B A,Gu X G,Gan Q L,Yang Y H.Brassinolide amelioration of aluminium toxicity in mung bean seedling growth [J].Plant Nutr.,2003,26:1725 -1734.
[7]Alam M M,Hayat S,Ali B,Ahmad A.Effect of 28-h(huán)omobrassinolidetreatment on nickel toxicity inBrassica juncea[J].Photosynthetica,2007,45:139-142.
[8]Arnon D I.Copper enzymes in isolated chloroplast:Polyphenoloxidase in Beta vulgaris[J].Plant Physiol.,1949,24:1 -15.
[9]Gong H J,Zhu X Y,Chen K Met al.Silicon alleviates oxidative damage of wheat plants in pots under drought[J].Plant Sci.,2005,169:313–321.
[10]Cakmak I,Marschner H.Magnesium deficiency and highlight intensity enhance activities of superoxide dismutase,ascro bate peroxidase,and glutathione reductase in bean leaves[J].Plant Physiol.,1992,98(4):1222 -1227.
[11]Giannopotitis C N,Ries S K.Superoxide dismutase:I.Occurrence in higher plants[J].Plant Physiol,1977,59:309 -314.
[12]Nakano Y,Asada K.Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloropasts[J].Plant Cell Physiol.,1981,(22):867 -880.
[13]趙世杰,許長成,鄒琦.植物組織中丙二醛測定方法的改進[J].植物生理學(xué)通訊,1994,30(3):207-210.
Zhao S J,Xu C C,Zou Q.Improvements of method for measurement of malondialdehvde in plant tissues[J].Plant Physiol.Comm.,1994,30(3):207-210.
[14]Mishra S,Srivastava S,Tripathi R Det al.Lead detox-ification by coontail(Ceratophyllum demersumL.)involves induction of phytochelatins and antioxidant systemin response to its accumulation[J].Chemosphere,2006,65:1027-1039.
[15]Piechalak A,Tomaszewska B,Baralkiewicz Det al.Accumulation and detoxification of lead ions in legumes[J].Phytochemistry,2002,60:153-162.
[16]Ruley A T,Nilesh S C,Sahi S V.Antioxidant defense in a lead accumulating plant,Sesbania Drummond [J].Plant Physiol.Biochem.,2004,42:899-906.
[17]王友保,劉登義.Cu,As及其復(fù)合污染對小麥生理生態(tài)指標(biāo)的影響[J].應(yīng)用生態(tài)學(xué)報,2001,12(5):773-776.
Wang Y B,Liu D Y.Effect of Cu,As and their combination pollution on ecophysio logical index of wheat[J].Trans.Chin.Soc.Agric.Eng.,2001,12(5):773 -776.
[18]Patsikka E,Kairavuo M,Season Fet al.Excess copper predisposes photosystemⅡto photoinhibitionin vivoby out competing iron and causing decrease in leaf chlorophyll[J].Plant Physiol.,2002,129:1359 -1367
[19]張義凱,韓小嬌,崔秀敏,等.外源NO對銅脅迫下番茄光合、生物發(fā)光特性及礦質(zhì)元素吸收的影響[J].植物營養(yǎng)與肥料學(xué)報,2010,16(1):174-180.
Zhang Y K,Han X J,Cui X Met al.Effects of exogenous nitric oxide on photosynthetic,bioluminescent characteristics and mineral element contents in tomato under copper stress[J].Plant Nutr.Fert.Sci.,2010,16(1):174 -180.
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
