施氮對海濱錦葵鹽脅迫下生物量分配特征及其關聯的影響1)
2012-09-18 11:11:06任燕燕連俊方閆道良
東北林業大學學報 2012年6期
任燕燕 連俊方 閆道良
(浙江農林大學,臨安,311300)
鹽脅迫所引起的水分脅迫和離子毒害等,對植物的生長、植被分布和農業生產均造成了較大的影響。為開發利用我國后備土地資源—鹽堿地,引種耐鹽經濟植物是最有效的途徑。海濱錦葵是一種多年生宿根植物,具較強的耐鹽性,且其宿根的更新周期長,適合在沿海灘涂大面積種植,改造鹽堿地。引種試驗表明,海濱錦葵具有較高的綜合開發利用前景[1-2]。
氮營養缺乏可能是鹽堿地區植物生長的主要限制因素之一。研究表明,施氮在一定程度上提高了逆境下(干旱)導致的水分脅迫對植物生長的影響[3]。對于鹽生植物而言,施氮對其在逆境下的生長的影響研究較少。如鹽生植物囊果堿蓬(Suaeda physophora Pall),硝態氮增加其高鹽脅迫下地上部有機氮,改善了植株的營養狀況和滲透調節,提高了囊果堿蓬的耐鹽能力。本文以鹽生植物海濱錦葵為材料,研究了施氮對鹽脅迫下海濱錦葵的生物量特征及其施氮前后生物量關聯的影響,為在鹽堿地科學栽培經濟海濱錦葵,提高其產量提供參考。
1 材料與方法
選取完好飽滿的海濱錦葵種子經表面消毒后于溫開水中浸泡2 h,播種于口徑為12 cm,高為11 cm的塑料缽內,苗缽放在規格一致的塑料碗內,缽內基質為河沙和珍珠巖按體積比3∶1配比。出苗后每盆保留健壯一致小苗6棵,待小苗有3~4片真葉完全展開后,分別用 50、150、250 mmol/L NaCl(分別記為 SC1,SC2,SC3)和 50、150、250 mmol/L NaCl+15 mmol/L NH4NO3(分別記為 SC-A1,SC-A2,SC-A3)處理,每塑料碗中加入處理液100 mL,并同時添加不含NH4NO3的改良Hoagland營養液10 mL,此時苗缽底部大約1.5 cm高度處于處理液中,每隔3~4 d更換1次處理液及添加相應營養液。每處理5個重復,共30缽。待小苗處理35 d后收獲測量。
把收獲的海濱錦葵植株于水中小心洗凈根部基質,測定株高、主根直徑(根頸下1 cm長度內)、主根長度之后110℃烘箱中殺青20 min,轉至80℃烘干至恒質量,測定全株干質量以及葉、莖、主根和須根干質量。計算葉重比為葉干質量/全株干質量(Leaf weight ratio,RLW)、莖重比為莖干質量/全株干質量(Stem weight ratio,RSW)、根重比為根干質量/全株干質量(Root weight ratio,RRW)和根冠比為根干質量/地上部分干質量(R/S)。
利用SPSS13.0對數據進行統計分析,處理間差異顯著性采用Duncan氏多重比較。圖表繪制借助Sigmaplot 10.0軟件。
2 結果與分析
2.1 施氮對鹽脅迫下海濱錦葵株高和根系的影響
當NaCl鹽濃度小于250 mmol/L時,施氮明顯增加了植物的高度,差異顯著(P<0.05)。隨著NaCl鹽濃度的提高,達250 mmol/L時,即使施以氮肥,植株的高度與不施氮的相比差異不明顯,說明氮肥施加并不能消除高鹽下對植株生長的抑制作用。無論施氮與否,隨著鹽梯度的增加,植株的高度變化不顯著,這也說明海濱錦葵對鹽變化不敏感,海濱錦葵具有較強的抗鹽特性,見表1所示。

表1 鹽脅迫下施氮處理對海濱錦葵株高和根系的影響
在3個NaCl處理水平條件下,主根長變化不明顯,不同的鹽濃度對主根長影響不大。施氮后,主根長也沒有發生明顯的變化。而主根直徑隨著鹽濃度的提升,逐漸減小。施氮后,主根直徑與不施氮相比有所增大。在低鹽(50 mmol/L)脅迫下時,施氮效果較為明顯。在鹽濃度小于150 mmol/L處理下,施氮可以緩解鹽脅迫對海濱錦葵主根增粗的抑制。在250 mmol/L NaCl處理條件下,施氮后的主根直徑(6.79 mm)比不施氮主根直徑(5.56 mm)雖有所增粗,但兩者差異并不顯著。說明高濃度的NaCl對主根增粗生長的抑制作用并不為施以氮肥而有所緩解。
2.2 施氮對鹽脅迫下海濱錦葵生物量及分配的影響
在NaCl鹽濃度小于150 mmol/L處理下,施氮顯著提高了全株的生物量。在150 mmol/L處理下施氮,植株生物量是不施氮的1.68倍,而在低鹽50 mmol/L處理下施氮則是不施氮的2倍,差異顯著(p<0.05)。當鹽濃度為250 mmol/L時施氮,同樣增加了植株的生物量,是不施氮的1.53倍,差異顯著(p<0.05)。在高鹽(250 mmol/L)脅迫下施氮植株生物量與小于150 mmol/L NaCl鹽處理下不施氮植株生物量差異不明顯(表2)。說明施氮大大減緩了由于鹽害造成的生物量積累的脅迫效應,有利于植物光合碳物質合成。
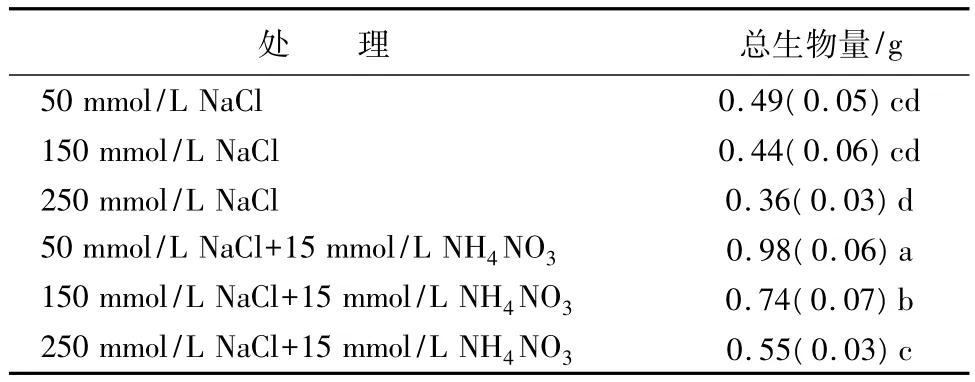
表2 鹽脅迫下施氮對海濱錦葵植株總生物量的影響
葉重比、莖重比和根重比反映了生物量在葉、莖和根3種器官之間分配的比例。從表3可以看出,施氮顯著提高了葉重比,在250 mmol/L NaCl處理下,施氮下的葉重比是不施氮的1.79倍,而在50 mmol/L NaCl處理下,施氮下的葉重比是不施氮的2.15倍,表明鹽脅迫下施氮有利于海濱錦葵葉片碳物質的積累。施氮對莖重比沒有明顯影響。根重比不論在低鹽和高鹽處理下,施氮比不施氮顯著降低海濱錦葵的根重比,在250 mmol/L NaCl處理下施氮的根重比約是不施氮的80%,表明鹽脅迫下施氮并沒有促進海濱錦葵在根系上的碳投資,反而對根系的生長有抑制作用。但施氮后主須根比有顯著的增加,尤其在鹽濃度小于150 mmol/L NaCl處理下,施氮效果更為明顯,如在150 mmol/L NaCl處理下,施氮下的主須根比是不施氮的2.18倍,這表明施氮有利于海濱錦葵主根的生長。
根冠比反映了生物量在地上和地下部分之間分配的關系(表4)。鹽脅迫下海濱錦葵根冠比在施氮條件下顯著下降。其中250 mmol/L NaCl處理下施氮的根冠比是非施氮的60%。結合根重比,可以看出施氮肥對海濱錦葵根的生長不利,分配到根部的生物量下降。
2.3 施氮處理下海濱錦葵葉、根和地上干質量與相關性狀的關聯性
植物的葉片和根作為植物體地上和地下部分的重要的營養器官,很多功能性狀在二者之間存在著一定的關聯性。本研究結果顯示(表4),施氮后鹽脅迫下的海濱錦葵葉干質量與其他性狀相關性得以加強,而未施氮肥鹽脅迫下的葉干質量僅與地上干質量表現相關。作為吸收養分的根和制造養料的葉在施氮后生物量顯著相關,線性回歸分析其兩者關系為 y(根干質量)=1.74x(葉干質量)+0.083(R2=0.703,F=16.592,p=0.005),而在施氮前并沒明顯的關聯。這說明施氮后更密切了葉和根功能器官的協調關系,有利于提高植株對逆境的抵抗。

表3 鹽脅迫下施氮處理對海濱錦葵生物量分配的影響

表4 鹽脅迫下施氮處理對海濱錦葵葉、根和地上干質量相關性的影響
3 結論與討論
植物生長環境中鹽分過多危害植物的正常生長和發育,生長抑制是植物對高鹽漬響應最敏感的生理現象。本實驗觀察到當鹽濃度處理小于150 mmol/L NaCl時,施氮增加了植株的高度。結合施氮后的葉重比來看,植株高度的增加更有利于在鹽逆境下通過承載莖上擴張的葉面積或增加葉數量來制造更多的碳水化合物。這與逆境(干旱)條件下,較大的冠層葉面積對作物產量的貢獻要大于較大根系的貢獻研究結果類似[3]。在本研究中,施氮顯著提高了海濱錦葵植株的總生物量,增加了鹽脅迫下海濱錦葵葉比重,而這種施氮的影響效果無論在低鹽(50 mmol/L NaCl)還是高鹽(250 mmol/L NaCl)環境下,沒有顯著差異。
植物最先感受逆境脅迫的器官是根系,逆境脅迫下根系形態上的變化是最為直觀的[4]。海濱錦葵在鹽脅迫下施氮并沒有顯著增加主根的長度,在小于150 mmol/L NaCl處理下施氮,主根直徑卻有顯著增加,主根的增粗相應增加海濱錦葵主根的干重,但是并沒有增加植株的根比重,相反,卻降低了根比重。這與施氮后降低海濱錦葵的根冠比有一致之處。根作為異養器官,適宜的根系似乎更有利于植株把逆境下“來之不易”的氮轉向于葉以此來合成積累更多的有機物質。這對于鹽生環境植物來說,是對變化的環境作出的適宜碳投資權衡。
目前,由于環境污染而導致的氮沉降有全球增加的趨勢[5],氮沉降在一定程度上對地上部分的生長有促進作用,對根系生長則不利,表現在氮沉降增加會使根部生物量生產減少[6]。這些結論在本研究中得以進一步證實。施氮后海濱錦葵根比重顯著下降,而葉比重顯著增加。從主須根比可以看出,施氮后增加了主根的生長,對須根的生長則有抑制作用,這與氮沉降抑制細根的生長有一致之處[6]。
我們的研究還發現,氮處理鹽脅迫下的海濱錦葵根與葉生物量關聯性顯著提高。這種更為密切的關系,或許是植物功能器官在更有利于生長的環境下,更好地協調生長過程中對資源的利用和分配[7]。
鹽影響鹽生植物的生長(促進或抑制),必然直接或間接影響植物的光合作用。施氮后,關于海濱錦葵的光合指標的變化需要進一步研究。另有研究表明,施氮增加了鹽生植物葉片脯氨酸的含量[8-9],降低耐鹽作物地上部Na+、Cl-的含量,增加K+的向上運輸和葉氮的含量[10-13]。對于海濱錦葵而言,植株的Cl-、Na+和K+在施氮前后的變化特征和施氮后對生殖生長的影響需要繼續研究。
[1]徐國萬,欽佩,謝民,等.海濱錦葵的引種生態學研究[J].南京大學學報,1996,32(2):268-274.
[2]趙可夫,馮立田.中國鹽生植物資源[M].北京:科學出版社,2001.
[3]張歲岐,徐炳成.根系與植物高效用水[M].北京:科學出版社,2010.
[4]原俊鳳,田長彥,馮固,等.硝態氮對鹽脅迫下囊果堿蓬幼苗根系生長和耐鹽性的影響[J].植物營養與肥料學報,2009,15(4):953-959.
[5]Matson P A,Mcdowell W H,Townsen A R,et al.The globalization of N deposition:ecosystem consequences in tropical environments[J].Biogeochemistry,1999,46(2):67-83.
[6]Persson H,Ahlstr?m K,Clemensson Lindell A.Nitrogen addition and removal at G?rdsj?n-effects on fine-root growth and fine-root chemistry[J].Forest Ecology and Management,1998,101(1/3):199-205.
[7]徐冰,程雨曦,甘慧潔,等.內蒙古錫林河流域典型草原植物葉片與細根性狀在種間及種內水平上的關聯[J].植物生態學報,2010,34(1):29-38.
[8]段德玉,劉小京,李存楨,喬海龍.N素營養對NaCl脅迫下鹽地堿蓬幼苗生長及滲透調節物質變化的影響[J].草業學報,2005,14(1):63-681.
[9]Naidoo G,Naidoo Y.Effects of salinity and nitrogen on growth,ion relations and proline accumulation in Triglochin bulbosa[J].Wetlands Ecol.Manage,2001,9(6):491-497.
[10]Gimeno V,Syvertsen J P,Nieves M.et al.Additional nitrogenfertilization affects salt tolerance of lemon trees on different rootstocks[J].Scientia Agricola,2011,68(3):298-305.
[11]Levy Y,Syvertsen J P.Irrigation water quality and salinity effects in citrus trees[J].Hort Agricola Rev,2004,30(4):37-82.
[12]Garcia-Sanchez F,Perez-Perez J G,Botia P,et al.The response of young mandarin trees grown under saline conditions depends on the rootstock[J].Eur J Agron,2006,24(1/3):129-139.
[13]沈振國,沈其榮,管紅英,等.NaCl脅迫下氮素營養與大麥幼苗生長和離子平衡的關系[J].南京農業大學學報,1994,17(1):22-26.
猜你喜歡
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年2期)2019-08-23 08:12:08
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
汽車觀察(2016年3期)2016-02-28 13:16:26
