鱸魚視黃酸受體α和視黃酸受體γ cDNA克隆和基因表達(dá)分析
2012-09-20 00:32:12錢云霞童彩環(huán)鄭偉賢
關(guān)鍵詞:小鼠
錢云霞 韓 柳 童彩環(huán) 鄭偉賢
(寧波大學(xué)生命科學(xué)與生物工程學(xué)院,應(yīng)用海洋生物技術(shù)教育部重點(diǎn)實(shí)驗(yàn)室,寧波 315211)
維生素A(視黃醇)是機(jī)體必需的一種營(yíng)養(yǎng)素,在哺乳動(dòng)物的生長(zhǎng)、發(fā)育、免疫和繁殖等生命過程中起著非常重要的作用[1]。與哺乳動(dòng)物一樣,維生素A也是魚類必不可少的營(yíng)養(yǎng)素,維生素A不足會(huì)導(dǎo)致魚類生長(zhǎng)減慢,出現(xiàn)眼、鰭和表皮出血等癥狀,嚴(yán)重時(shí)會(huì)引起死亡[2-4]。維生素A在體內(nèi)的代謝產(chǎn)物主要是視黃酸,即由視黃醇類氧化成視黃醛,視黃醛再氧化成視黃酸[5]。維生素A的多種生物學(xué)功能都是通過視黃酸信號(hào)途徑實(shí)現(xiàn)的,視黃酸作為配體只有運(yùn)輸?shù)郊?xì)胞核內(nèi)與視黃酸受體(retinoic acid receptor,RAR)和視黃醇類X受體(retinoid X receptor,RXR)形成的異源二聚體結(jié)合,才能識(shí)別并結(jié)合到目的基因啟動(dòng)子上的視黃酸作用元件(retinoic acid response element,RARE),進(jìn)而實(shí)現(xiàn)對(duì)目的基因的轉(zhuǎn)錄調(diào)節(jié)[6-7]。RARE的結(jié)構(gòu)特點(diǎn)是5'-AGGTCA-3'單元的直接重復(fù)序列(direct repeat,DR),DR間存在1~5個(gè)堿基的插入,分別稱為 DR1、DR2、DR3、DR4 和 DR5,而RAR二聚體主要識(shí)別 DR5,也識(shí)別 DR1和DR2[8]。RAR在哺乳動(dòng)物中有3種亞型,即RARα、RARβ 和 RARγ,分別由不同的基因編碼[9]。作為核受體超家族的成員,RAR分子具有典型的核受體結(jié)構(gòu),由非保守的N端轉(zhuǎn)錄激活域(transactivation domain,A/B 區(qū))、保守的DNA 結(jié)合區(qū)(DNA-binding domain,DBD,C 區(qū))、鉸鏈區(qū)(linker region,D 區(qū))、配體結(jié)合區(qū)(ligand-binding do-main,LBD,E 區(qū))和 F 區(qū)組成[10]。所有的全反式視黃酸和9-順式視黃酸都是RAR的配體,與其他配體依賴的轉(zhuǎn)錄激活受體不同,當(dāng)配體存在時(shí),RAR二聚體與RARE結(jié)合并進(jìn)一步激活轉(zhuǎn)錄;而配體不存在時(shí),RAR二聚體也可以與RARE結(jié)合,但同時(shí)與維甲酸和甲狀腺激素受體沉默調(diào)節(jié)子(silencing mediator of retinoic acid receptor and thyroid hormone receptor,SMRT)和核受體輔阻遏物(nuclear receptor corepressor,N-CoR)等抑制子結(jié)合,從而抑制目的基因的轉(zhuǎn)錄[11]。由于許多基因都受RAR途徑的調(diào)控,因而RAR參與機(jī)體的許多生命過程[12-13],但關(guān)于魚類RAR的研究只有很少的報(bào)道,RAR基因僅在斑馬魚(Danio rerio)、紅鰭東方鲀(Takifugu rubripes)、佛羅里達(dá)雀鱔(Lepisosteus platyrhincus)和褐牙鲆(Paralichthys olivaceus)等少數(shù)幾種魚類中被分離和鑒定[14-15]。
鱸魚(Lateolabrax japonicus),屬鱸形目鮨科鱸屬,屬于廣鹽性肉食性兇猛魚類,是我國(guó)沿海主要的海水養(yǎng)殖魚類之一。本研究克隆了鱸魚的RARα和RARγ基因cDNA序列,并分析了它們的結(jié)構(gòu)及在各個(gè)組織中的表達(dá),期望能為研究RAR基因在鱸魚視黃酸信號(hào)中的作用提供基礎(chǔ)資料。
1 材料與方法
1.1 試驗(yàn)材料
試驗(yàn)用2齡鱸魚購買于寧波水產(chǎn)品大世界。
總RNA提取試劑(Trizol)購自美國(guó)Invitrogen公司;擴(kuò)增試劑盒(SMARTTMRACE cDNA Amplification Kit)購自美國(guó)Clontech公司;逆轉(zhuǎn)錄試劑(PrimeScriptTM1stStrand cDNA Synthesis Kit)、Taq酶和pMD-18T載體均購自日本TaKaRa公司;間氨基苯甲酸乙酯甲磺酸鹽(MS-222)為美國(guó)Sigma公司產(chǎn)品;其他試劑為國(guó)產(chǎn)分析純?cè)噭T囼?yàn)所用引物由上海捷瑞生物工程有限公司合成。大腸桿菌(Escherichia coli)DH5α由本實(shí)驗(yàn)室保存。
1.2 總RNA的提取和cDNA第1條鏈的合成
鱸魚經(jīng)MS-222麻醉后取肝臟組織,參照Trizol試劑操作說明提取總RNA,經(jīng)紫外分光光度計(jì)(NanoDrop 1000 Spectrophotometer)檢測(cè)總RNA的濃度和純度,用1.2%的瓊脂糖凝膠電泳檢測(cè)總RNA的完整性。取總RNA 1 μg參照逆轉(zhuǎn)錄試劑盒操作說明書合成cDNA第1條鏈。
1.3 RAR全長(zhǎng)cDNA擴(kuò)增
根據(jù)人(Homo sapiens,BC071733)、牛(Bos Taurus,NM_001014942)、雞 (Gallusgallus,X78335)、大西洋鮭(Salmo salar,AF290610)、佛羅里達(dá)雀鱔(Lepisosteus platyrhincus,AB292640)、斑馬魚(Danio rerio,NM_131399)和紅鰭東方鲀(Takifugu rubripes,NM_001032753)的 RARα 基因cDNA序列設(shè)計(jì)RACE兼并引物(表1)。
同理根據(jù)人(BC098421)、小鼠(Mus musculus,NM_001042727)、雞 (X73973)、牛 (NM_001130756)、青鳉魚(Oryzias latipes,EF546455)、佛羅里達(dá)雀鱔(AB292642)、斑馬魚(BC098519)和虹鱒(Oncorhynchus mykiss,NM_001246348)的RARγ基因cDNA序列設(shè)計(jì)RACE兼并引物(表1)。
以上述肝臟cDNA為模板,用兼并引物分別擴(kuò)增 RARα和 RARγ基因核心片段。其中 RARα基因的擴(kuò)增條件為:預(yù)變性94℃ 3 min;94℃30 s,56~52 ℃ 30 s,72 ℃ 1 min,共進(jìn)行 36 個(gè)循環(huán)(前4個(gè)循環(huán)退火溫度每循環(huán)下降1℃);最后72℃延伸5 min。RARγ基因的梯度退火溫度為60~55℃,其他條件均與RARα基因相同。PCR產(chǎn)物經(jīng)1.2%的瓊脂糖凝膠電泳檢測(cè),目標(biāo)片段割膠純化后與pMD18-T載體連接,連接產(chǎn)物轉(zhuǎn)化大腸桿菌DH5α感受態(tài)細(xì)胞。PCR檢測(cè)后將陽性克隆送上海英駿生物工程公司進(jìn)行序列測(cè)定。
參照SMARTTMRACE cDNA Amplification Kit說明書,將鱸魚肝臟總RNA分別合成5'端和3'端RACE cDNA片段(5'-and 3'-RACE-ready cDNA)。在測(cè)序得到的核心序列的基礎(chǔ)上,分別設(shè)計(jì)RARα和RARγ基因5'端和3'端 RACE特異性引物(表1)。PCR產(chǎn)物純化、克隆和測(cè)序方法同上。最后對(duì)所獲得的序列進(jìn)行拼接,獲得RAR全長(zhǎng)cDNA序列。
1.4 序列分析
應(yīng)用BioEdit軟件進(jìn)行氨基酸序列推導(dǎo)及同源性分析。經(jīng) ScanProsite軟件(http://www.expasy.ch/prosite/)對(duì)鋅指結(jié)構(gòu)進(jìn)行預(yù)測(cè)。使用MEGA 4.0軟件,通過鄰位相連(neihbor joining,NJ)法對(duì)鱸魚RAR氨基酸序列和GenBank中已收錄的其他已知?jiǎng)游镞M(jìn)行分子進(jìn)化樹聚類分析。
1.5 RAR基因在各組織中表達(dá)分析
根據(jù)得到的RAR基因序列設(shè)計(jì)逆轉(zhuǎn)錄PCR特異性引物,序列見表1。提取鱸魚肝臟、肌肉、心臟、眼、腸、腎臟、脂肪、脾臟、鰓和大腦10個(gè)組織的總RNA,分別用無RNA酶的DNA酶(RNase free DNase)消化,各取1 μg參照 PrimeScriptTM1stStrand cDNA Synthesis Kit說明書逆轉(zhuǎn)錄合成cDNA。以18S RNA為內(nèi)參基因(序列見表1),檢測(cè)RAR基因在不同組織中的相對(duì)表達(dá)量。18S RNA的PCR反應(yīng)條件如下:94℃預(yù)變性3 min;94 ℃ 50 s,54 ℃ 40 s,72 ℃ 40 s,26 個(gè)循環(huán)(18S RNA的表達(dá)量較目標(biāo)基因高);72℃延伸2 min。RARα與RARγ基因的退火溫度分別56和54℃,循環(huán)數(shù)均為28,其他條件與18S RNA相同。
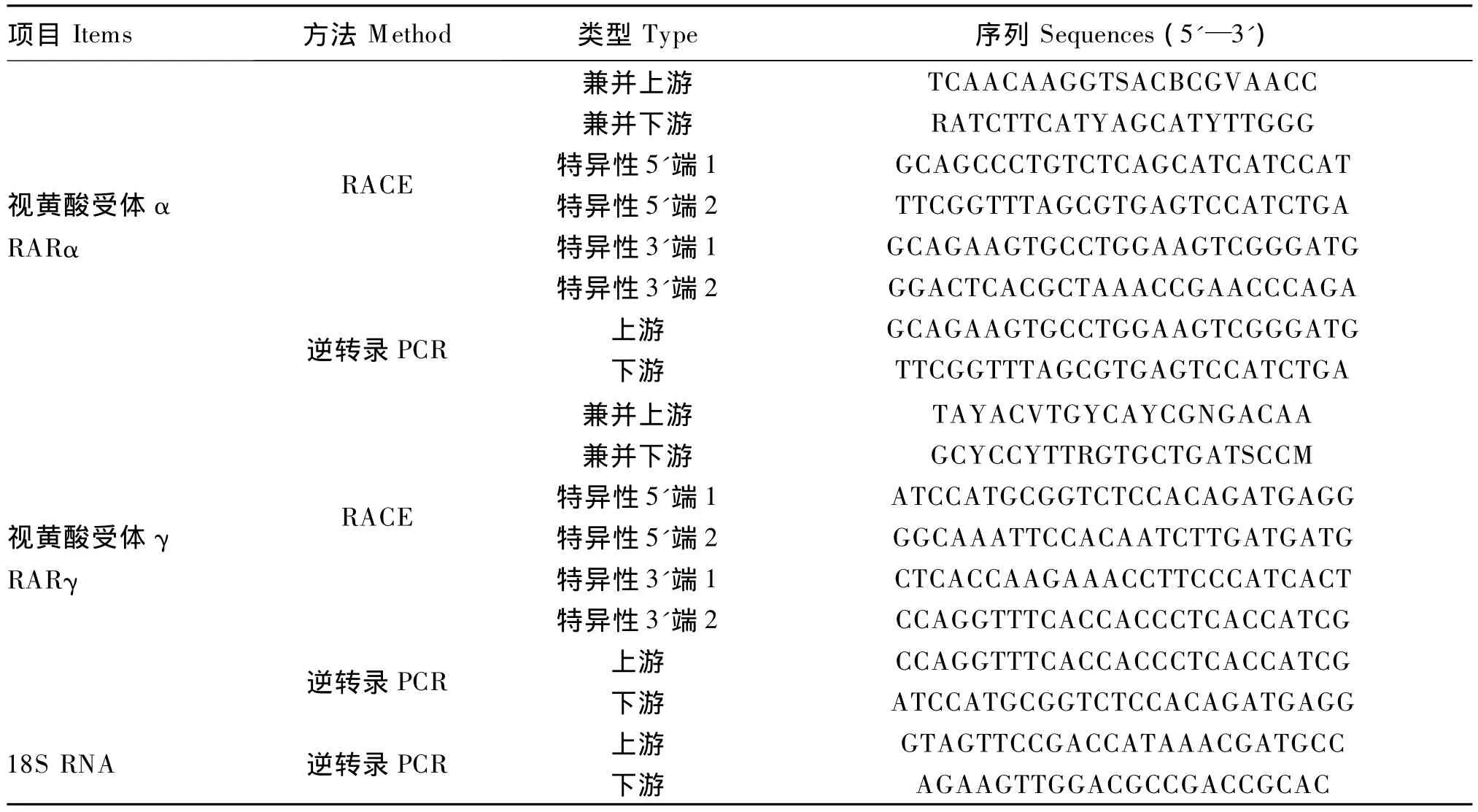
表1 PCR引物序列Table1 Primer sequences for PCRs
2 結(jié)果
2.1 RAR cDNA的克隆及序列分析
首先,利用RARα的兼并引物從鱸魚肝臟cDNA中PCR得到749 bp的核心片段,再由RACE特異性引物5'端1和5'端2進(jìn)行2輪RACE,得到5'RARα產(chǎn)物1 001 bp,與核心序列有489 bp的重疊區(qū)域。由RACE特異性引物3'端1和3'端2得到3'RACE產(chǎn)物1 112 bp,與核心序列有279 bp的重疊區(qū)域。拼接結(jié)果表明,RARα cDNA全長(zhǎng)序列為2 094 bp,其中5'非翻譯區(qū)(5'-untranslated region,5'-UTR)為 124 bp,3'非翻譯區(qū)(3'-untranslated region,3'-UTR)為 608 bp,開放閱讀框(open readingframe,ORF)為 1 362 bp,編碼 453 個(gè)氨基酸(圖1),分子質(zhì)量為50.64 ku,理論等電點(diǎn)為8.20,GenBank登陸號(hào)為JN226572。氨基酸序列同源性分析發(fā)現(xiàn),鱸魚RARα與尼羅羅非魚(Oreochromis niloticus)、紅鰭東方鲀、斑馬魚和人的RARα的同源性分別為97.6%、96.9%、91.0%和83.5%。
同樣,利用RARγ兼并引物克隆得到815 bp的RARγ核心片段,在得到的核心序列的基礎(chǔ)上,設(shè)計(jì)5'端RACE和3'端RACE特異性引物分別進(jìn)行2輪RACE,得到5'RACE產(chǎn)物和3'RACE產(chǎn)物分別為758和927 bp,與核心序列分別有369和437 bp的重疊。最后拼接獲得RARγ cDNA全長(zhǎng)序列共 1 671 bp,其中 5'-UTR 36 bp,3'-UTR 129 bp,ORF 1 506 bp,共編碼 501 個(gè)氨基酸(圖2),理論等電點(diǎn)為4.96,分子質(zhì)量為56.20 ku,GenBank登陸號(hào)為 JU226571。分析表明,鱸魚RARγ氨基酸序列與紅鰭東方鲀、斑馬魚、大西洋鮭和人的同源性分別為95.2%、90.2%、89.9%和74.1%。

圖1 鱸魚RARα基因的全長(zhǎng)cDNA序列及推導(dǎo)的氨基酸序列Fig.1 The full length cDNA and the deduced amino acid sequences of the RARα gene of Lateolabrax japonicus
2.2 RAR的結(jié)構(gòu)與系統(tǒng)進(jìn)化分析
用BioEdit軟件將鱸魚RARα和RARγ氨基酸序列和斑馬魚 RARα(NP_571481)、RARγ(CAM16092)以及人 RARα(NP_001019980)、RARγ(NP_001036193)的氨基酸序列進(jìn)行比對(duì)分析。結(jié)果顯示,鱸魚RARα和RARγ的N端為保守性很低的A/B區(qū),A/B區(qū)末端為非配體依賴的激活功能(ligand-independent transactivation function,AF-1)區(qū)結(jié)構(gòu)。其次,鱸魚RARα和RARγ均具有保守的DBD和LBD,DBD包括參與識(shí)別目標(biāo)基因作用元件的P盒子(P box)結(jié)構(gòu)和參與二聚體形成的D 盒子(D box)結(jié)構(gòu)[16],經(jīng) ScanPros-ite預(yù)測(cè),DBD含有2個(gè)鋅指結(jié)構(gòu)(圖3)。鱸魚RARα和RARγ的LBD均含有11個(gè)α螺旋(H1和H3~H12)和2個(gè)β轉(zhuǎn)角結(jié)構(gòu),其H12的C端均含有配體依賴的激活功能(ligand-dependent transactivation function,AF-2)區(qū),其序列分別為L(zhǎng)IQETLEN和LIREMLEN。圖4為用MEGA 4.0軟件的NJ法構(gòu)建的鱸魚RARα、RARγ和其他物種的RAR氨基酸序列的系統(tǒng)樹。結(jié)果顯示,RARα、RARβ和RARγ 3種亞型分別成簇,鱸魚的RARα和RARγ分別與紅鰭東方鲀的 RARα和RARγ聚為一支,再與斑馬魚等其他魚類的RARα和RARγ聚合,然后與哺乳類和兩棲動(dòng)物聚合。

圖2 鱸魚RARγ的全長(zhǎng)cDNA序列及其推導(dǎo)的氨基酸序列Fig.2 The full length cDNA and the deduced amino acid sequences of the RARγ gene of Lateolabrax japonicus
2.3 RAR基因在各組織中的表達(dá)
從圖5可見,RARα基因除在心臟和大腦中表達(dá)量較少外,其他組織中的表達(dá)量均較高,以眼、鰓和腸中的表達(dá)量最高,其次是肝臟、脾臟、脂肪和腎臟。RARγ基因僅在鰓和脾臟中有較高表達(dá),其他組織檢測(cè)到的表達(dá)量都較低。

圖3 鱸魚RARα和RARγ的氨基酸序列與斑馬魚和人的多重比對(duì)結(jié)果Fig.3 Multiple alignment of amino acid sequences of RARα and RARγ among Lateolabrax japonicus,Danio rerio and Homo sapiens
3 討論
維生素A的多種生物學(xué)功能都是通過RAR和RXR 2種核受體起作用的[7]。人和小鼠RARα、RARβ和 RARγ分別由3種不同基因編碼,由于不同的拼接方式或使用不同的啟動(dòng)子使得每個(gè)亞型又會(huì)產(chǎn)生多種不同的類群,同一亞型的不同類群在DBD和LBD的序列是完全相同的,差異主要在A/B區(qū)[17]。本研究分離到了鱸魚RARα和RARγ 2種視黃酸受體基因,二者氨基酸序列同源性高達(dá)63.1%,但沒有克隆到RARβ基因亞型,Hale等[15]對(duì)斑馬魚的研究中也沒克隆到該亞型。
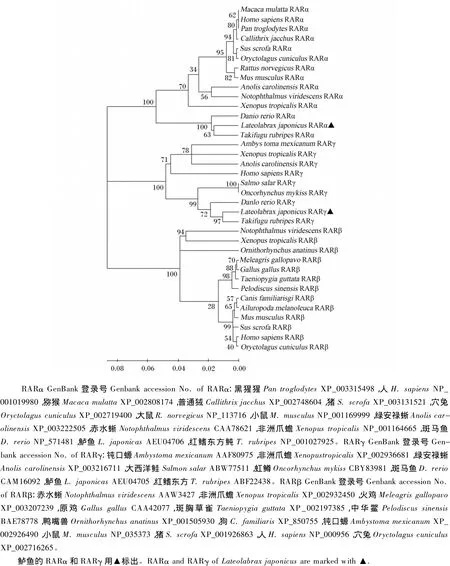
圖4 鱸魚和其他動(dòng)物RAR的氨基酸序列構(gòu)建的系統(tǒng)進(jìn)化樹Fig.4 Phylogenetic analysis of RARs based on amino acid sequence of Lateolabrax japonicus and other species
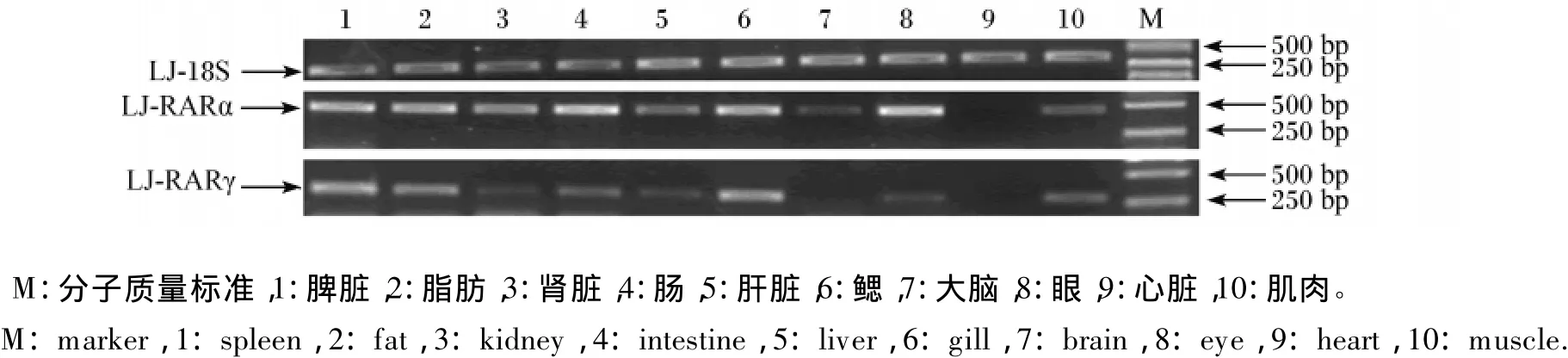
圖5 鱸魚RARα和RARγ基因在各組織中的表達(dá)Fig.5 Expressions of RARα and RARγ genes of Lateolabrax japonicus in various tissues
RAR的結(jié)構(gòu)分析發(fā)現(xiàn),鱸魚的 RARα和RARγ包括保守性低的A/B區(qū)、非常保守的DBD和LBD、D區(qū)和C端的F區(qū)6個(gè)功能區(qū)。DBD非常保守,鱸魚RARα的DBD共76個(gè)氨基酸,與斑馬魚、小鼠和人RARα的DBD的同源性高達(dá)97.3%、97.3%和94.7%。鱸魚RARγ與斑馬魚、人和小鼠在DBD的同源性分別為100.0%,96.0%和93.7%。另外,鱸魚RARα和RARγ的在DBD的同源性也高達(dá)92.1%。可見DBD作為最保守的功能區(qū)不僅在不同物種的同一亞型間同源性很高,且在不同亞型間也很保守。序列分析發(fā)現(xiàn),鱸魚DBD含有2個(gè)鋅指結(jié)構(gòu)(每個(gè)鋅指結(jié)構(gòu)中有4個(gè)不變的半胱氨酸)及P盒子和D盒子結(jié)構(gòu)。多重比對(duì)分析發(fā)現(xiàn),鱸魚、斑馬魚和人的RARα和RARγ的P盒子序列完全相同,均為EGCKG,但鱸魚和斑馬魚的D盒子在RARα和RARγ中分別是CHREKN和CHRDKN,而人的卻都是CHRDKN。相比之下,LBD的保守性沒有 DBD高,鱸魚RARα的 LBD與斑馬魚、小鼠和人的 RARα的LBD的同源性分別為96.7%、90.2%和89.7%,鱸魚RARγ與斑馬魚、小鼠和人的LBD的同源性分別為97.5%、97.1%和92.6%。鱸魚RARα與RARγ在LBD同源性為82.0%。核受體超家族成員的LBD大多有12個(gè)α螺旋,而鱸魚的RARα和RARγ與人的RARγ相似,其LBD都缺少H2結(jié)構(gòu)[18]。另外,鱸魚和其他動(dòng)物一樣,配體激活域AF-1和AF-2在RARα和RARγ中均只有1個(gè)氨基酸的差異,即AF-1的第3個(gè)氨基酸在RARα為P,而在 RARγ為 S,AF-2的第 3個(gè)氨基酸在RARα為Q,而在RARγ為R。作者還發(fā)現(xiàn),與其他動(dòng)物的AF-2第5個(gè)氨基酸都為M不同,鱸魚的RARα為T,但這種差異對(duì)鱸魚RARα的轉(zhuǎn)錄激活功能是否有影響還有待研究。與RARα和人的RARγ不同,鱸魚和斑馬魚的RARγ在C末端延伸區(qū)(F區(qū))都有不同數(shù)目的氨基酸序列多出,但是其功能尚不清楚。
鱸魚2種RAR在肝臟、肌肉、心臟、眼、腸、腎臟、脂肪、脾臟、鰓和大腦等10個(gè)組織中均有表達(dá),這種廣泛的組織分布表明這些組織都可能存在RAR的目標(biāo)基因。其中鱸魚RARα基因在眼、腸、肝、脾、脂肪和腎等檢測(cè)組織中都有明顯的表達(dá),以眼、鰓和腸中表達(dá)量最高,這點(diǎn)與哺乳動(dòng)物的RARα廣泛地分布于的大腦、皮膚、肌肉、心臟和腎臟等組織中的結(jié)果一致[19-20]。相比之下,RARγ基因在所檢測(cè)組織中表達(dá)量均較低,僅在鰓和脾臟中有較高表達(dá)。鱸魚RARα和RARγ基因在組織中這種表達(dá)特點(diǎn)與小鼠類似,在小鼠胚胎發(fā)育過程中RARα基因在大多數(shù)組織中都有表達(dá),而RARγ基因的表達(dá)有一定的選擇性[21]。雖然單個(gè)亞型的RAR缺陷小鼠都沒有視力缺陷,但胎兒或出生后個(gè)體會(huì)出現(xiàn)維生素A缺乏癥狀,且RARα無效突變體小鼠出現(xiàn)生精上皮退化,最終導(dǎo)致雄性不育,而RARγ失活會(huì)導(dǎo)致個(gè)體骨骼和上皮缺陷,說明RARα和RARγ在不同的組織中可能有著不同的功能[22-23],所以RARα和RARγ在鱸魚的各個(gè)組織中的功能差異及其在鱸魚維生素A的信號(hào)傳導(dǎo)過程中所起的作用還有待研究。
4 結(jié)論
①本試驗(yàn)成功克隆了RARα和RARγ的全長(zhǎng)cDNA。
②鱸魚RARα和RARγ有著典型的核受體結(jié)構(gòu),在各組織中廣泛分布。
[1]CLAGETT-DAME M,DELUCA H F.The role of vitamin A inmammalian reproduction and embryonic development[J].Annual Review of Nutrition,2002,22:347-381.
[2]MOREN M,OPSTAD I,HAMRE K,et al.An optimum level of vitamin A supplements for Atlantic halibut(Hippoglossus hippoglossus L.)juveniles[J].Aquaculture,2004,235:587-599.
[3]HEMRE G I,DENG D F,WILSON R P,et al.Vitamin A metabolism and early biological responses in juvenile sunshine bass(Morone chrysops×M.saxatilis)fed graded levels of vitamin A[J].Aquaculture,2004,235:645-658.
[4]YANG Q H,ZHOU X Q,JIANG J,et al.Effect of dietary vitamin A deficiency on growth performance,feed utilization and immune responses of juvenile Jian carp(Cyprinus carpio var.Jian)[J].Aquaculture Research,2008,39:902-906.
[5]DUESTER G.Retinoic acid synthesis and signaling during early organogenesis[J].Cell,2008,134(6):921-931.
[6]LIU R Z,SHARMA M K,SUN Q,et al.Retention of the duplicated cellular retinoic acid-binding protein 1 genes(crabp1a and crabp1b)in the zebrafish genome by subfunctionalization of tissue-specific expression[J].Federation of European Biochemical Societies,2005,272(14):3561-3571.
[7]ROCHETTE-EGLY C,GERMAIN P.Dynamic and combinatorial control of gene expression by nuclear retinoic acid receptors(RARs)[J].Nuclear Receptor Signaling,2009,7:e005.
[8]DELACROIX L,MOUTIER E,ALTOBELLI G,et al.Cell-specific interaction of retinoic acid receptors with target genes in mouse embryonic fibroblasts and embryonic stem cells[J].Molecular Cell Biology,2010,30:231-244.
[9]BASTIEN J,ROCHETTE-EGLY C.Nuclear retinoid receptors and the transcription of retinoid-target genes[J].Gene,2004,328:1-16.
[10]GERMAIN P,STAELS B,DACQUET C,et al.Overview of no menclature of nuclear receptors[J].Pharmacological Reviews,2006,58:685-704.
[11]KOIDE T,DOWNES M,CHANDRARATNA R A,et al.Active repression of RAR signaling is required for head formation[J].Genes & Development,2001,15:2111-2121.
[12]NAGASHIMA M,SAKURAI H,MAWATARI K,et al.Involvement of retinoic acid signaling in goldfish optic nerve regeneration[J].Neurochemistry International,2009,54:229-236.
[13]LINVILLE A,RADTKE K,WAXMAN J S,et al.Combinatorial roles for zebrafish retinoic acid receptors in the hindbrain,limbs and pharyngeal arches[J].Developmental Biology,2009,325:60-70.
[14]WENTWORTH J M,SCHOENFELD V,MEEK S,et al.Isolation and characterisation of the retinoic acid receptor-alpha gene in the Japanese pufferfish,F(xiàn).rubripes[J].Gene,1999,236(2):315-323.
[15]HALE A,TALLAFUSS A,YAN Y L,et al.Characterization of the retinoic acid receptor genes raraa,rarab and rarg during zebrafish development[J].Gene Expression Patterns,2006,6(5):546-555.
[16]PERLMANN T,RANGARAJAN P N,UMESONO K,et al.Determinants for selective RAR and TR recognition of direct repeat HREs[J].Genes & Development,1993,7:1411-1422.
[17]CHAMBON P.A decade of molecular biology of retinoic acid receptors[J].Federation of American Societies for Experimental Biology,1996,10:940-954.
[18]RENAUD J P,ROCHEL N,RUFF M,et al.Crystal structure of the RAR-γ ligand-binding domain bound to all-trans retinoic acid[J].Nature,1995,378:681-689.
[19]MENG Q Y,CHEN X N,ZHAO J,et al.Distribution of retinoic acid receptor-α immunoreactivity in the human hypothalamus[J].Neuroscience,2011,174:132-142.
[20]DAY R M,LEE Y H,PARK A M,et al.Retinoic acid inhibits airway smooth muscle cell migration[J].A-merican Journal of Respiratory Cell and Molecular Biology,2006,34:695-703.
[21]DOLLE P,RUBERTE E,LEROY P,et al.Retinoic acid receptors and cellular retinoid binding proteins.Ⅰ.A systematic study of their differential pattern of transcription during mouse organogenesis[J].Development,1990,110:1133-1151.
[22]LUFKIN T,LOHNES D,MARK M,et al.High postnatal lethality and testis degeneration in retinoic acid receptor α mutant mice[J].Proceedings of the National Academy of Sciences,1993,90:7225-7229.
[23]CHAPELLIER B,MARK M,MESSADDEQ N,et al.Physiological and retinoid-induced proliferations of epidermis basal keratinocytes are differently controlled[J].The EMBO Journal,2002,21:3402-3413.
猜你喜歡
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學(xué)大眾(2021年6期)2021-07-20 07:42:44
科學(xué)(2020年3期)2020-11-26 08:18:30
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級(jí))(2017年10期)2017-10-10 01:00:05
中國(guó)男科學(xué)雜志(2016年4期)2016-12-01 05:20:09
湖北師范大學(xué)學(xué)報(bào)(自然科學(xué)版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
