一株底棲硅藻——雙眉藻的優化培養
2012-10-13 08:14:00周一春胡超群彭鵬飛張呂平
海洋科學 2012年10期
周一春, 胡超群, 彭鵬飛, 羅 鵬, 張呂平
(1. 中國科學院 南海海洋研究所, 廣東 廣州 510301; 2. 中國科學院 研究生院, 北京100049)
海洋底棲硅藻是鮑、海參和海膽等名貴水產動物苗種生產中的重要餌料, 是影響幼苗成活率和質量的關鍵因素之一[1-2]。與鮑的人工育苗相似, 在海參人工育苗中, 底棲硅藻影響海參幼蟲的附著與變態, 并且海參育苗成功與否、產量的高低與底棲硅藻的數量與質量有著直接的關系。目前, 北方仿刺參(Apostichopusjaponicus)的繁育及養殖技術已經成熟,但熱帶海參人工繁殖和養殖才則剛剛起步, 熱帶海參的營養需要及繁育中適合的餌料種類研究均處于空白狀態, 制約了熱帶海參養殖業的發展[3-4]。
在硅藻的大規模培養中, 氮、磷、硅營養元素質量濃度和配比以及鹽度、光照、溫度等物理因子被認為是影響硅藻生長的重要因子, 不同的培養條件對底棲硅藻的種群結構及其生物量影響顯著[5-6]。同時有研究表明營養元素比例的微小變動會導致原環境代表種類相對豐度的顯著改變[7-8]。此外, 底棲硅藻種類豐富多樣, 分布廣泛, 生存環境差異甚大, 一種固定的標準化培養液(如傳統的“f/2”培養液)并不能滿足眾多單種底棲硅藻的快速生長的營養需求[9]。因此研究底棲硅藻的最適培養條件對底棲硅藻的規模培養, 獲得優質餌料具有重要的指導意義和應用價值。
作者從西沙永興島附近海域分離到一株底棲硅藻—雙眉藻(Amphorasp.), 經海參養殖實踐證實該種硅藻是糙刺參(Stichopus horrens)喜食的優良餌料。本文首先探討溫度、鹽度、光照和不同質量濃度的氮、磷、硅鹽對Amphorasp.生長的影響, 并進一步研究氮、磷、硅三種營養鹽多因子組合對其生長的影響, 以期確定Amphorasp.最佳培養條件, 為熱帶海參的底棲硅藻餌料的進一步研究及應用提供了理論依據。
1 材料與方法
1.1 藻種來源
實驗所用藻種采自西沙永興島附近海域, 通過富集培養, 分離并純化得到。借助光鏡和電鏡技術,研究了該藻的形態學特征, 并通過18S rDNA和5.8S rDNA-ITS序列, 對這株藻進行了多相分類鑒定, 判定該藻屬于橋彎藻科(Cymbellaceae)、雙眉藻屬(Amphora), 定名為Amphorasp.。
1.2 藻種培養
藻種的初期維持培養采用添加有 Na2SiO3的 f/2培養基, 培養溫度為(30±0.5)℃, 光照強度為 85~93μmol/(m2·s), 光照周期為 12L:12D, 每天定時進行搖瓶。實驗培養液為天然海水配制, 鹽度為 35, 所用藥品為分析純。實驗器皿為1L玻璃缸、50~1 000mL三角錐瓶和 10 mL離心管, 所有實驗器皿均先經過121℃、20 min高壓滅菌, 冷卻后使用。
1.3 藻生長的測定方法
1.3.1 光密度法
在上述培養條件下, 玻璃缸內傾斜放置有若干面積相等的附著基, 每天定時隨機取出3片, 刮取附著基上的藻細胞, 稀釋到適當的質量濃度, 充分混勻后用于生長的測定。光密度法參照文獻[10-11]進行測定, 每次測樣前搖勻, 每次取樣測 8~10個值,從中去除最大值和最小值, 然后求平均代表該樣值。
比生長速率計算公式為:K=(lgAt2-lgAt1)/0.301(t2-t1), 其中,K為相對生長常數;t1為指數生長期開始時的天數;t2為指數生長期結束時的天數;At1為指數生長期開始的藻液光密度值;At2為指數生長期結束的藻液光密度值。
1.3.2 細胞計數法
細胞計數采用血球計數板, 每次取樣重復計數3次。
1.4 單因子試驗設計
1.4.1 營養鹽單因子試驗
本組實驗的物理因子為: 鹽度 35, 溫度 30℃,光照強度(85±7)μmol/(m2·s), 光照周期為 12L:12D,培養時間為8 d。
N、P、Si的質量濃度梯度設置及實驗方案如下:
以添有Na2SiO3的f/2培養液的N、P、Si為基礎, 固定 N、Si質量濃度, 改變 P質量濃度進行試驗; 同理, 固定 P、Si質量濃度, 改變 N質量濃度和固定N、P質量濃度改變Si進行試驗, 分別設置6 個梯度: 0、1/100、1/10、1、10、100(表 1)。每組設置 3個平行, 選取指數期的藻種加入各個質量濃度的培養液中, 并使每種單因子的初始培養液的A680nm值基本相同。

表1 實驗N、P、Si的質量濃度梯度設置Tab. 1 The gradient concentrations of N, P, Si
1.4.2 環境因子實驗
本組實驗培養液采用添加Na2SiO3的f/2培養液,光照周期為12L:12D, 培養時間為8d。每個物理因子梯度設置如表2, 每組設置3個平行, 選取指數期的藻種加入各個質量濃度梯度的培養液中, 并使每種單因子的初始培養液的A680nm值基本相同。
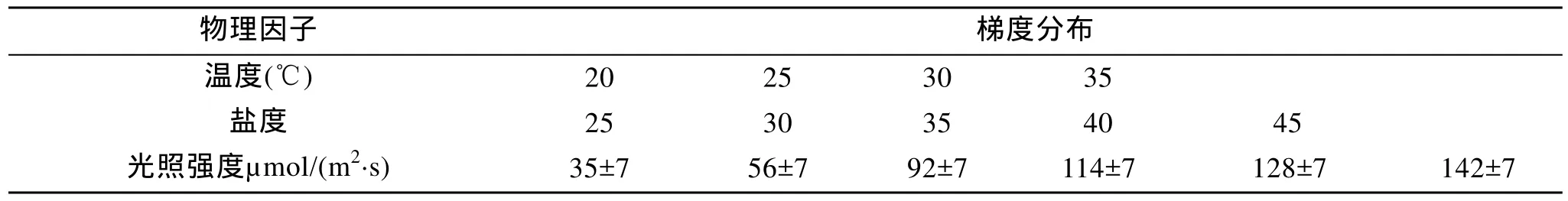
表2 物理因子梯度設置Tab. 2 The gradients of Physical factors
1.5 數據的處理
實驗數據采用 SPSS11.0軟件進行統計學分析,依據不同的實驗需要做相關性分析、方差分析和LSD多重比較, 顯著性水平為P<0.05。
2 結果
2.1 光密度法和細胞計數法檢測 Amphora sp.生長的相關性
由圖1可知, 不同的接種密度培養Amphorasp.,光密度法和細胞計數法顯示很好的相關性, 兩種方法反映該藻生長趨勢基本一致, 通過Pearson相關性檢測, A圖和B圖的相關系數為0.997和0.996, 顯著性相關(P=0.01)。
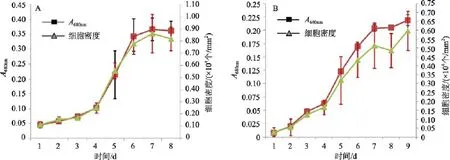
圖1 Amphora sp.在不同接種密度下的光密度和細胞計數法的相關性Fig. 1 Correlation between the values of OD680 and their cell counts per area of Amphora sp. under different inoculations
2.2 光照強度對Amphora sp.生長的影響
不同光照強度對底棲硅藻Amphorasp.生長的影響如圖2所示。單因素方差分析表明, 該藻的相對生長常數差異顯著(P=0.000), 說明光照強度對Amphorasp.的生長影響顯著, 多重比較得知,Amphorasp.最適生長光照強度為 56~99 μmol/(m2·s)。
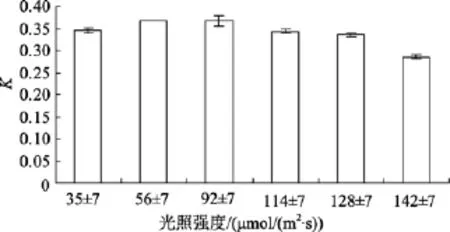
圖2 不同光照強度對Amphora sp.生長的影響Fig. 2 Effect of light intensity on growth of Amphora sp.
2.3 溫度對Amphora sp.生長的影響
不同的溫度對Amphorasp.生長的影響如圖 3所示。單因素方差分析表明, 該藻的相對生長常數差異顯著(P=0.000), 說明不同溫度對Amphorasp.的生長影響顯著。當溫度很低時, 該藻只能極緩慢地生長, 在20~30℃時, 隨著溫度的升高, 比增長速率K增大, 通過LSD多重比較得知, 該藻的最適溫度在25~30℃。
2.4 鹽度對Amphora sp.生長的影響
不同的鹽度對Amphorasp.生長的影響如圖4所示, 單因素方差分析表明, 該藻的相對生長常數差異顯著(P=0.003), 說明不同鹽度對Amphorasp.的生長影響顯著, LSD多重比較得知, 該藻的最適鹽度30~35, 鹽度過高或過低都會抑制該藻的生長, 且高鹽比低鹽抑制明顯。
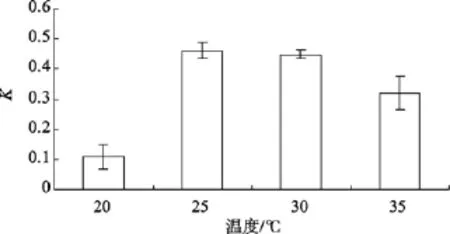
圖3 不同溫度對Amphora sp.生長的影響Fig. 3 Effect of temperature on growth of Amphora sp.
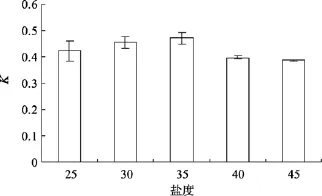
圖4 不同鹽度對Amphora sp.生長的影響Fig. 4 Effect of salinity on growth of Amphora sp.
2.5 氮質量濃度對Amphora sp.生長的影響
不同氮的質量濃度對Amphorasp.生長的影響如圖5所示, 單因素方差分析表明, 該藻的相對生長常數差異顯著(P=0.000), 說明不同氮質量濃度對Amphorasp.的生長影響顯著, LSD多重比較得知, 在氮質量濃度0~750 mg/L范圍內, 隨著氮質量濃度的增加, 該藻的相對生長速率也逐漸增大, 且表明該藻在質量濃度為750 mg/L和7 500 mg/L時, 該藻的相對生長速率K沒有差別(P>0.05)。
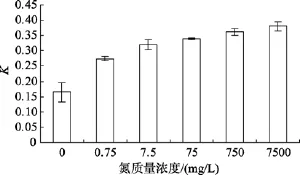
圖5 不同氮質量濃度對Amphora sp.生長的影響Fig. 5 Effect of NaNO3-N concentration on growth of Amphora sp.
2.6 磷質量濃度對Amphora sp.生長的影響
不同磷質量濃度對Amphorasp.生長的影響如圖6所示, 在一定范圍內, 隨著磷質量濃度的增加, 該藻的生長也加快, 單因素方差分析表明, 該藻的相對生長常數差異顯著(P=0.000), 說明不同磷質量濃度對Amphorasp.的生長影響顯著。多重比較得知, 44 mg/L時相對生長常數達到最大。
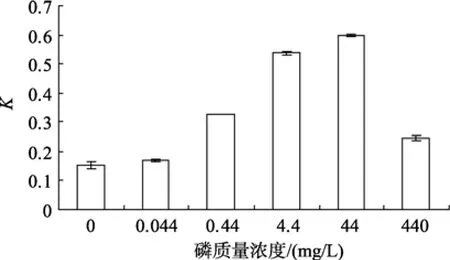
圖6 不同磷質量濃度對Amphora sp.生長的影響Fig. 6 Effect of NaH2PO4-P concentration on growth of Amphora sp.
2.7 硅質量濃度對Amphora sp.生長的影響
不同硅質量濃度對Amphorasp.生長的影響如圖7所示, 該種底棲硅藻的生長速率在硅鹽質量濃度為零時非常低, 隨著硅鹽質量濃度的升高而迅速增加。單因素方差分析, 表明該藻的比生長速率差異極顯著(P=0.000), 說明不同硅質量濃度對Amphorasp.的生長影響顯著, 多重比較得知,該藻的最適硅質量濃度為150 mg/L。
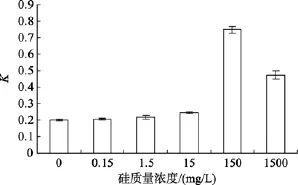
圖7 不同硅質量濃度對Amphora sp.生長的影響Fig. 7 Effect of Na2SiO3-Si concentration on growth of Amphora sp.
2.8 氮、磷、硅的正交實驗
2.8.1 正交試驗設計
選擇NaNO3-N、Na2SiO3-Si、NaH2PO4-P三因子對Amphorasp.進行了生長速率的影響試驗, 均取三水平(表 3), 各因子水平的確定主要根據本實驗中單因子試驗結果。正交試驗表4選用L9(34)。
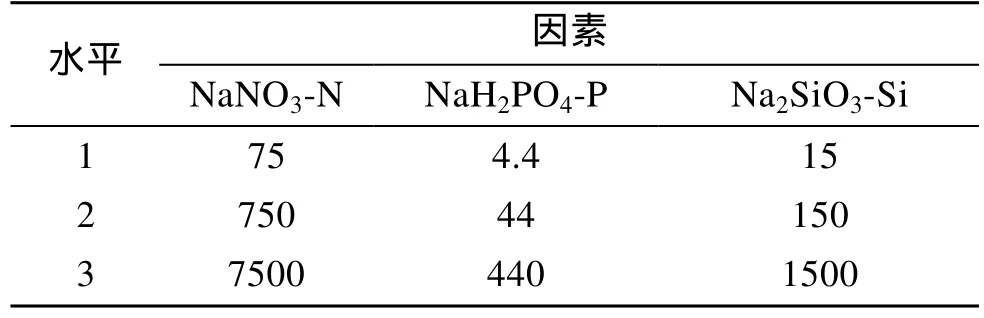
表3 正交試驗因素及水平Tab. 3 Factors and levels of orthogonal tests mg/L
表4是Amphorasp.營養鹽L9(34)正交試驗結果,由極差數值R可見, 三種營養鹽的質量濃度對該藻的影響作用由強到弱依次是: NaH2PO4-P>NaNO3-N>Na2SiO3-Si。正交實驗方差分析(表 5)表明 3種營養鹽對Amphorasp.的生長都有顯著影響, 其中磷酸二氫鈉、硝酸鈉的作用極其顯著, 綜合分析得出培養Amphorasp.的氮、磷、硅的最適質量濃度配比是N:P:Si=2.4:1:3.4, 即本實驗的2號實驗。
3 討論
由圖1可知, 不同的接種密度培養Amphorasp.,兩種方法反映該藻生長趨勢基本一致。Pearson相關性檢測A圖和B圖的相關系數為0.997和0.996, 呈顯著性相關(P=0.01), 光密度法和細胞計數法顯示很好的相關性。在生長對數期末期到穩定期, 可能由于該藻分泌胞外分泌物, 較之細胞計數,A680nm值會略高, 但兩種方法在反映藻細胞生長狀態上顯示較好的相關性, 這與之前的研究一致[10], 隨著培養時間的延長, 細胞生長經穩定期進入衰亡期, 細胞數會減少, 但細胞的胞外分泌物并不會立即停止, 二者數據的顯示就會出現很大的差異, 相關性較低[12]。光密度法檢測該藻生長率選用A680nm, 研究表明在該波長是, 分光光度計的靈敏度最高, 結果較為可靠。作者認為, 在進行A值檢測操作時盡量保證數據的穩定性, 進行多次測量, 避免數據的偶然性會得出準確的結果。該方法在檢測各項指標的優劣性上是可行的, 尤其在檢測的指標較多時, 該方法較為便捷,也能在一定程度避免細胞計數的主觀誤差。
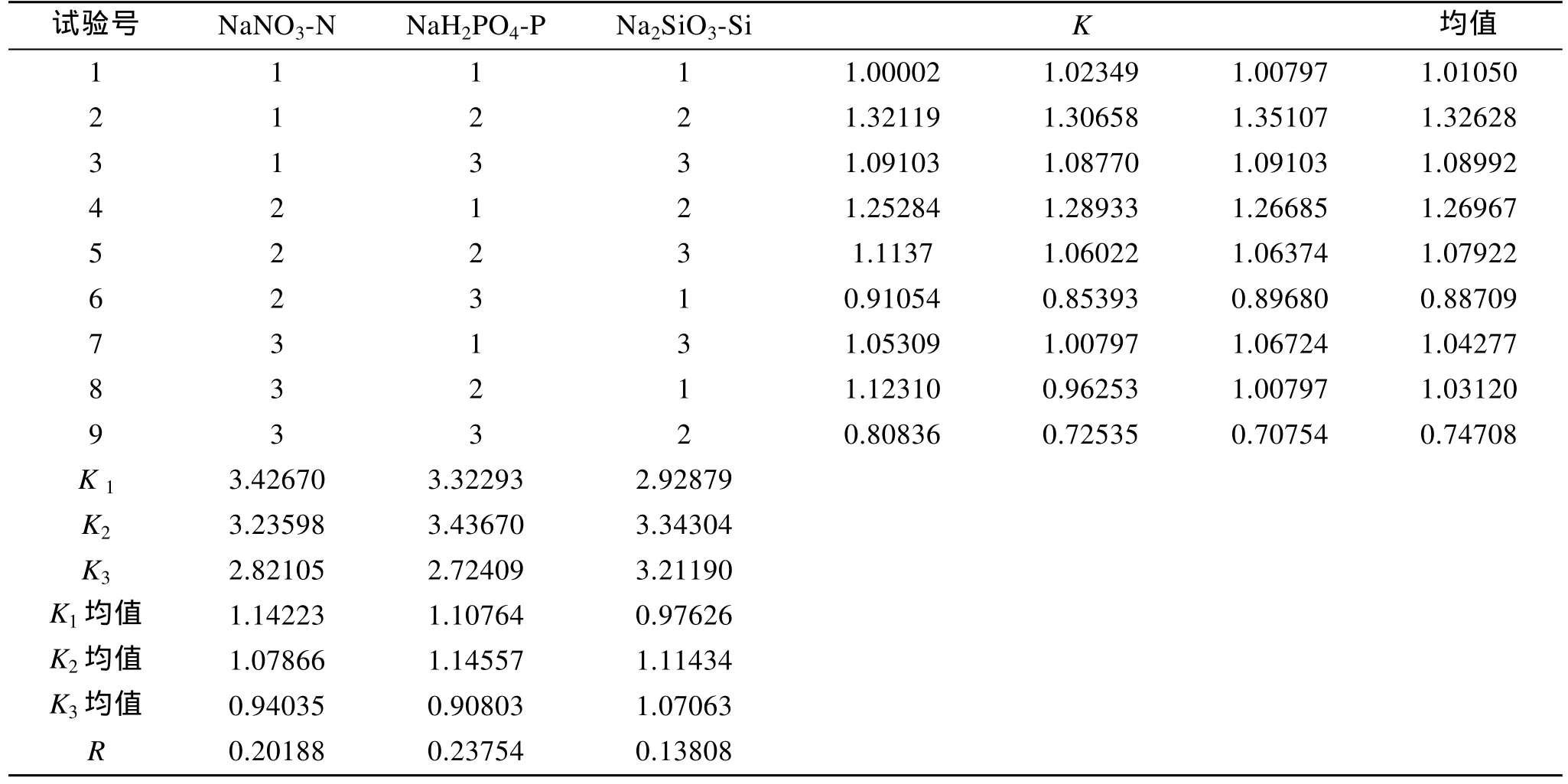
表4 正交試驗結果Tab. 4 Results of orthogonal test L9(34)design
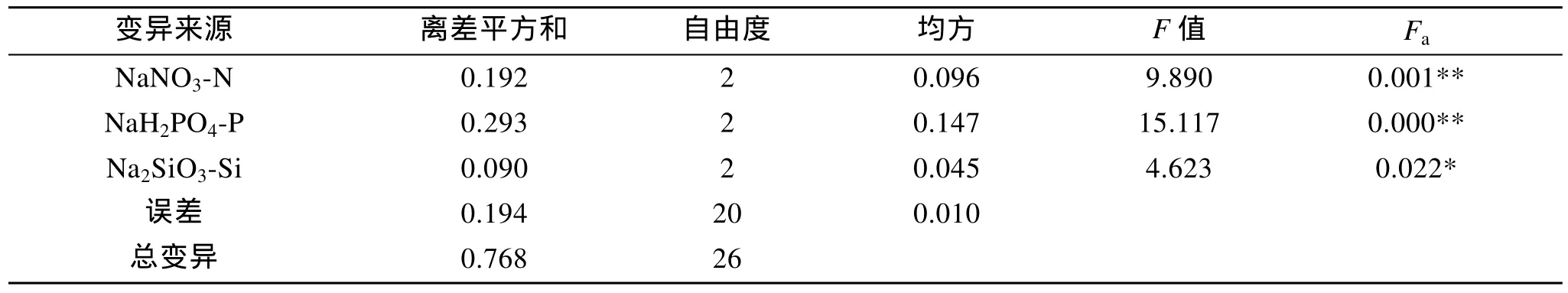
表5 正交結果的方差分析Tab. 5 Variance analysis of orthogonal test results
底棲硅藻營自養生活, 必須依靠光合作用來合成有機物質。光強和光質都會影響底棲硅藻的光合作用以及藻的群落、種群動態和適應模式[13]。郭峰等報道了底棲硅藻適宜生長的光強一般為21.4~78.6μmol/(m2·s)[14], 最適光強因種類不同而有所區別, 如亞歷山大菱形藻(Nitzschia alexandrina)適于光照較強的環境, 在 78.6 μmol/(m2·s)迅速生長, 而卵形(Cocconeissp.)則在 47.1μmol/(m2·s)生長最好[14]。光過強會發生光飽和現象, 甚至發生光氧化。本實驗數據表明Amphorasp.適宜生長的光強為 56~99μmol/(m2·s)。與上述藻類最適光照強度不同原因可能是Amphorasp.分離自西沙熱帶海域的潮間帶, 水體的透明度非常高。Amphorasp.適宜生長光照值反映了棲息的環境特征。
溫度是影響藻類細胞生理變化的主要因素[15]。對光自養生物, 溫度通過控制酶動力學來調節其生長[16]。硅藻多在溫度較低的冬春季生長繁殖[17], 各種硅藻各有其適溫范圍, 高溫和低溫都會危害底棲硅藻的生長。Amphorasp.適宜溫度為25~30℃, 與丁蕾等[18]報道的擬菱形藻(Pseudo-nitzschiasp.)高密度出現在 25~30℃相似, 但比一般的底棲硅藻的適宜溫度 10~20℃高, 與該藻生長的熱帶西沙海域偏高溫的環境相一致。
鹽度在不僅影響硅藻的分布, 而且是使某些硅藻形成水華的重要原因。在最適范圍內生長、繁殖速度最快。超出適鹽范圍, 過高和過低的鹽度都會傷害藻類細胞都會產生[15]。馬美榮等[19]培養的咖啡雙眉藻(Amphora coffeaeformis)在鹽度在8~60時生長速率逐漸減慢, 郭峰等[14]培養的矮小卵形藻(Cocconeis diminuta)和亞歷山大菱形藻(Nitzschia alexandrina)的最適鹽度分別為30和35,在20~40 鹽度范圍都能良好的生長, 與本實驗結果基本一致。同時 Clavero等[20]在34株底棲硅藻的鹽度試驗中發現, 大部分培養的種屬表現出極度的廣鹽性,最適鹽度在30。說明不同藻種最適質量濃度不同, 本實驗Amphorasp.采自西沙永興島海域, 最適生長鹽度為30~35, 也是長期適應的結果。
氮是構成蛋白質的重要元素, 是影響底棲硅藻細胞生長與分裂的重要元素[18], 是底棲硅藻生長與繁殖的一個限制性因子。實驗結果表明, 在 0~750 mg/L的質量濃度范圍內, 隨著氮質量濃度的增加,藻細胞比生長速率也相應增加, LSD多重比較得知,該藻在質量濃度為750 mg/L和7500 mg/L時, 該藻的相對生長速率K沒有差別(P>0.05)。董金利等[21]研究了氮、磷營養鹽對底棲硅藻縊縮菱形藻(Nitzschiaconstricta)的生長及生化組成影響中表明,該藻在最大氮質量濃度900 mg/L時, 該藻的比生長速率達到最大。一般認為, 不同的藻種有不同的最適氮質量濃度。在藻類的規模化培養中, 營養鹽的投入多少直接影響成本的高低, 選擇合適的氮質量濃度既要保證藻類的較快速生長, 又要講究社會效益。
由于硅藻的硅質外殼, 硅是硅藻的必需營養元素.除了作為細胞壁結構成分外, 還參與光合色素的合成、蛋白質的合成、DNA的合成和細胞分裂等多種代謝和生長過程[22]。 盡管如此, Shifrin[23]曾報道,一種小環藻在硅饑餓后大約一代時間內, 細胞分裂就完全停止, 但馬志珍[24]在實驗室用自然海水培養單種底棲舟形藻(Neviculasp.)時也發現, 即使不加硅鹽, 硅藻仍能生長良好。大貝政治[25]等采用營養去除法的實驗表明, 用完全不含硅的人工海水培養液培養一種卵形藻Cocconeissp.時, 對其生長的抑制作用很小。由圖6可知, 當硅鹽質量濃度為零時, 雙眉藻仍能較緩慢生長, 但在0~15 mg/L時, 該藻的相對生長常數沒有太大變化, 當硅的質量濃度增加到150 mg/L時,Amphorasp.呈快速增長,進一步證明傳統的 f/2海水培養基不能滿足該藻快速生長的營養需要。
磷是細胞膜的重要成分, 在細胞的能量傳遞中起著重要的作用, 而培養液中的磷對培養液的酸堿度起緩沖作用[18]。在藻類培養中, 磷的用量一般都比氮小, 但由于磷的化合物的溶解性和移動性較差,磷對水體初級生產力的限制作用更顯著[26], 本文的單因子試驗中這一結論同樣得到證實。有研究表明,微藻對營養鹽的攝取不是單獨進行的, 而是按一定比例吸收, 營養鹽之間的比例, 尤其是氮磷的質量濃度配比對細胞的生長發育有重要作用[27-28]。各種單細胞藻所要求的N、P比例不同。王淵源等[29]實驗表明, 小型舟形藻(Navicula parva)最佳 N: P=65:1,在此條件下細胞數量增加最為明顯。單因子試驗沒有考慮營養鹽之間的配比, 存在一定的缺陷。藻類生長狀態是多因子同時作用的結果, 所以多因子組合實驗更能真實反映它們的營養需求, 更接近藻類培養的最優條件。本實驗在N:P:Si=2.4:1:3.4時該雙眉藻的生長最好, 而與李雅娟等[9]培養的咖啡雙眉藻(Amphora coffeaeformis)的最佳配比為 2.5:1:2接近,兩者都與馬志珍[24]報道的舟形藻(Naviculasp.)最適質量濃度都不同, 說明不一樣的藻類適宜的營養鹽比例有異。同是雙眉藻, 分布在不同的海域, 環境條件不同, 也會表現出不同的營養需求。
[1]陳明耀. 生物餌料培養 [M]. 北京: 中國農業出版社,1995.
[2]陳世杰, 陳木, 盧豪魁, 等. 鮑苗的餌料-底棲硅藻培養試驗初報 [J]. 動物學報, 1977, 23(1): 47-52.
[3]袁成玉. 海參飼料研究的現狀與發展方向 [J]. 水產科學, 2005, 24(012): 54-56.
[4]張春云, 王印庚, 榮小軍, 等. 國內外海參自然資源、養殖狀況及存在問題[J]. 海洋水產研究, 2004,25(3): 89-97.
[5]Rivkin R B, Putt M. Photosynthesis and cell division by antarctic microalgae:comparion of benthic, planktonic and ice algae [J]. Journal of phycology, 1987, 23: 223-229.
[6]Snyder E B, Robinson C T, Minshall G W. et al. Regional patterns in periphyton accrual and diatom assemblage structure in a heterogeneous nutrient landscape [J]. Canadian Journal of Fisheries and Aquatic Sciences, 2002, 59(3): 564-577.
[7]Agatz M, Asmus R, Deventer B. Structural changes in the benthic diatom community along a eutrophication gradient on a tidal flat [J]. Helgoland Marine Research,1999, 53(2): 92-101.
[8]Armitage A R, Frankovich T A, Fourqurean J W. Variable responses within epiphytic and benthic microalgal communities to nutrient enrichment [J]. Hydrobiologia,2006, 569(1): 423-435.
[9]李雅娟, 王起華. 氮, 磷, 鐵, 硅營養鹽對底棲硅藻生長速率的影響 [J]. 大連水產學院學報, 1998,13(4): 7-14.
[10]Wang Q H, Wang S H, Ding M J, et al. Studies on culture conditions of benthic diatoms for feeding abalone[J]. Chinese Journal of Oceanology and Limnology,1998, 16(1): 78-83.
[11]周銀環. 氮, 磷, 鐵, 硅對簡單雙眉藻生長的影響[J]. 廣東海洋大學學報, 2008, 28(1): 35-39.
[12]Hoagland K D, Rosowski J R, Gretz M R, et al. Diatom extracellular polymeric substances: function, fine structure, chemistry and physiology[J]. Journal of phycology, 1993, 29:537-566.
[13]莊樹宏. 光強和光質對底棲藻類群落影響Ⅱ. 群落和種群的動態和適應模式 [J]. 生態學報, 2001, 21(12):2057-2066.
[14]郭峰, 朱凌俊, 柯才煥, 等. 兩種海洋底棲硅藻的培養條件研究 [J]. 廈門大學學報(自然科學版), 2005,44(6):831-835.
[15]華汝成. 單細胞藻類的培養與利用 [M]. 北京: 農業出版社, 1986.
[16]Davison I R. Environmental effects on algal photosynthesis: temperature [J]. Journal of phycology, 1991,27(1): 2-8.
[17]陳峰, 姜悅. 微藻生物技術 [M]. 北京: 中國輕工業出版社, 1999.
[18]丁蕾, 支崇遠. 環境對硅藻的影響及硅藻對環境的監測 [J]. 貴州師范大學學報: 自然科學版 2006, 24(3):13-16.
[19]馬美榮, 李朋富, 陳麗, 等. 鹽度和營養限制對鹽田底棲硅藻咖啡雙眉藻生長及胞外多糖產率的影響[J]. 鹽業與化工, 2008, (5): 30-34.
[20]Clavero E, Hernández-Mariné M, Grimalt J O, et al.. Salinity tolerance of diatoms from thalassic hypersaline environments [J]. Journal of Phycology, 2000, 36(6):1021-1034.
[21]董金利, 莊惠如, 占美憐,等. 氮, 磷營養鹽對底棲硅藻的生長及生化組成影響[J]. 生物技術,2010,20:64-67.
[22]Werner D. The biology of diatoms [M]. California:Univ of California Press, 1977.
[23]Shifrin N S, Chisholm S W. Phytoplankton lipids:interspecific differences and light-dark cycles[J].Journal of phycology, 1981, 17(4): 374-384.
[24]馬志珍, 季梅芳, 陳匯遠. 一種可做鮑和海參餌料的底棲舟形藻的培養條件的研究[J]. 海洋通報, 1985,4(4): 36-39.
[25]大貝政治, 松井敏夫, 高木博之. 附著硅藻Cocconeis sp.の增殖に及 ぽ環境諸要因の影響[J].水產增殖, 1992, 40(2): 240-246.
[26]吳春雪. 兩種海洋底棲硅藻生長條件及生化成分的研究[D]. 大連: 大連理工大學, 2008.
[27]Smith R E H, Geider R J. Kinetics of intracellular carbon allocation in a marine diatom[J]. Journal of Experimental Marine Biology And Ecology, 1985, 93(3):191-210.
[28]Thomas W H, Dodson A N. Effects of phosphate concentration on cell division rates and yield of a tropical oceanic diatom [J]. The Biological Bulletin, 1968,134(1): 199-208.
[29]王淵源, 姜慶國, 江航宇. 培養小型舟形藻的氮、磷肥料量 [J]. 海洋科學, 1986, 10(5): 35-37.
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
