RhoB抑制慢性粒細胞白血病細胞的生長
2012-12-25 01:43:48姚紅朱曉蘇徐康萍周海俠趙昀
東南大學學報(醫學版) 2012年5期
姚紅,朱曉蘇,徐康萍,周海俠,趙昀
(1.蘇州大學唐仲英血液學研究中心,江蘇蘇州 215123;2.蘇州大學附屬第一醫院血液科,江蘇蘇州 215006)
人類慢性粒細胞白血病(chronic myeloid leukemia,CML)是源自造血干細胞的惡性血液腫瘤。格列衛能緩解慢性期CML患者的癥狀,但單一藥物尚不能治愈。尋找新的靶點、設計新藥物已成為CML研究領域的熱點[1]。哺乳動物Rho家族屬于小GTP結合蛋白,包括22個成員[2]。其中RhoA,Rac1和Cdc42已被廣泛研究。大量研究發現Rho蛋白調控癌細胞的增殖、存活、浸潤與血管新生[3]。例如,RhoA、RhoC 以及Rac1、Cdc42 促進腫瘤形成、癌細胞侵襲和轉移[4]。RhoB是Rho家族中較為特殊的一員,在實體腫瘤中則作為抑制子或負性調控子。脂類修飾是調控RhoB功能的重要方式之一,RhoB蛋白能進行法尼基(F)和牻牛兒基牻牛兒基(GG)兩種修飾,這與多數其它Rho蛋白僅能進行一種類型的脂類修飾不同[5]。這些研究為實體腫瘤的治療提供了新思路。但是,目前RhoB在人CML中的作用及其機制尚不明確。本研究使用慢病毒在人CML細胞中過表達RhoB,研究其對CML細胞增殖能力的影響;使用幾種與RhoB脂類修飾相關的突變體研究RhoB是否需要脂類修飾行使其生物學功能及何種類型的脂類修飾是RhoB生物學功能所必需的;探討RhoB及各種脂類修飾變體對CML細胞周期進程的影響,初步揭示RhoB對調控CML細胞生長的作用機制,為CML的治療提供實驗基礎及理論依據。
1 材料和方法
1.1 材料
1.1.1 細胞 人成巨核細胞白血病MEG-01細胞系(BCR-ABL陽性)購于上海中國科學院細胞庫。CML患者骨髓樣本來源于蘇州大學附屬第一醫院。
1.1.2 主要試劑 Ex Taq DNA聚合酶、PMD19-T載體、T4DNA連接酶、EcoRⅠ、XhoⅠ、BglⅡ限制性內切酶均購于TaKaRa公司,碘化丙啶、RNase A和鼠抗人單克隆Actin抗體購于Sigma公司。PCR引物合成及基因序列檢測均由上海生物工程公司完成。兔抗人RhoB多克隆抗體購于Santa Cruz Biotechnology公司。CD34細胞純化試劑盒、纖粘連蛋白、甲基纖維素均購于StemCell Technologies公司。
1.1.3 主要儀器 流式細胞分析及分選儀(美國BD,Beckman Coulter),紫外凝膠成像儀(美國 Bio-Rad),垂直電泳儀和轉印電泳儀(上海天能),PCR儀(德國Biometra),倒置顯微鏡(日本Olympus)。
1.2 實驗方法
1.2.1 RhoB基因及其3個脂類修飾變體的克隆 參考NCBI網站公布的RhoB cDNA序列,設計如表1所示PCR引物[5]。PCR擴增出目的片段后使用T4DNA連接酶與PMD19-T載體連接,用BglⅡ將測序正確的目的基因切割純化,再與線性化的Venus載體連接。用EcoRⅠ和XhoⅠ酶切鑒定連接正反向。

表1 RhoB基因及其3個脂類修飾變體PCR引物設計Tab 1 List of primer sequences
1.2.2 慢病毒的制備 將慢病毒表達載體通過磷酸鈣沉淀的方式將其與其他3種包裝質粒△R,VSV-G和Rev共轉染293T細胞,收集轉染后48和72 h的培養上清,0.22 μm濾膜過濾后進行分裝,-80℃保存備用。將病毒上清進行1次超離心濃縮,獲得超過100倍的濃縮病毒。
1.2.3 CML患者CD34+細胞的純化與病毒侵染 將CML患者骨髓樣本經密度梯度離心分離出骨髓單核細胞,利用免疫磁珠正向富集的方法純化出CD34+細胞。細胞培養2 d后進行病毒侵染。先將濃縮病毒加到包被有纖粘連蛋白的96孔板中,4℃放置1~2 h后加入細胞,37℃共孵育12 h,PBS洗滌細胞3次后繼續培養,72 h后進行流式分選,YFP+細胞用于實驗。
1.2.4 細胞增殖和集落生成實驗 細胞增殖實驗:在24孔板中每孔種植5×104個YFP+細胞,培養的第3天和第6天分別進行細胞計數。集落生成(colonyforming cell,CFC)實驗:在甲基纖維素中加入雙抗、IMDM培養基和細胞,混勻后取1.1 ml接種到直徑為3.5 cm小皿中,37℃培養,2周后進行集落計數。
1.2.5 細胞周期分析 取1×106YFP+的細胞,70%乙醇固定過夜,加入 RNase A(100 μg·ml-1),37 ℃孵育30 min,再加入碘化丙啶(40 μg·ml-1),流式細胞儀檢測[6]。
1.2.6 蛋白質印跡法檢測過表達的蛋白 收集2×106YFP+細胞,PBS清洗2次,加入蛋白裂解液,冰上裂解15 min,4℃ 14 000×g離心20 min,收集上清,進行蛋白定量。蛋白樣品加入5×SDS加樣緩沖液,100℃煮沸5 min,進行12%SDS-PAGE,轉移至PVDF膜,5%脫脂奶粉室溫封閉1 h,以合適的稀釋比加入一抗4℃搖床過夜,TBST洗滌4次,加入辣根過氧化物酶標記的二抗,室溫孵育1 h,最后用 ECL試劑顯影[7]。
1.3 統計學分析
所有實驗均重復3次以上,數據采用GraphPad Prism 5.0處理。結果以±s表示,組間比較采用t檢驗。P<0.05表示差異有統計學意義。
2 結 果
2.1 基因克隆與載體構建
如圖1A所示,基因測序結果證實我們成功獲得野生型RhoB、脂類修飾缺失的變體RhoBC193G、只進行F修飾的變體RhoB-F(C-端殘基CKVL突變為CAIM)及只進行GG修飾的變體RhoB-GG(C-端殘基CKVL突變為CLLL)。并成功構建了表達RhoB及其變體的各類慢病毒載體,目的基因由SFFV(Spleen Focus Forming Virus)啟動子驅動,YFP報告基因可示蹤病毒轉導的細胞(圖1B)。蛋白質印跡實驗顯示RhoB及其變體均獲得了成功表達(圖1C)。
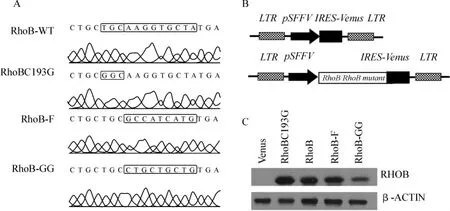
圖1 基因克隆與載體構建A.Gene sequence data;B.Schematic structure of lentiviral vector;C.Western blotting to detect the protein expression Fig 1 Gene cloning and vector construction
2.2 RhoB抑制MEG-01細胞的增殖
如圖2A所示,過表達RhoB后,培養第3天Venus組和RhoB組的細胞增殖并沒有顯著差異,而第6天RhoB組的細胞生長明顯受到抑制(P<0.01)。另外,在集落生成實驗中RhoB組的細胞集落生成能力與Venus組相比受到顯著抑制(P<0.01),抑制率約為45%(圖2B)。
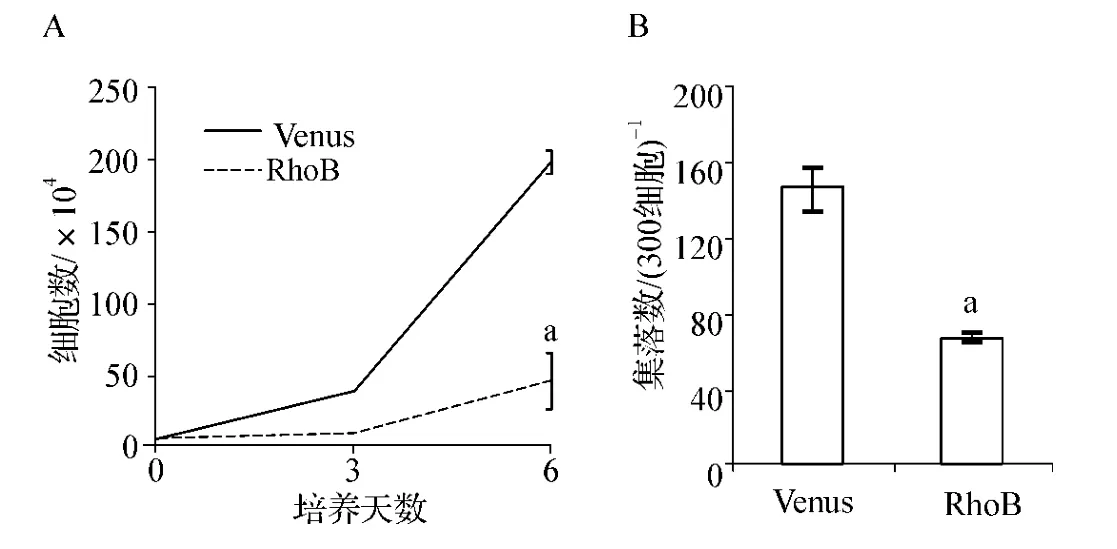
圖2 RhoB對MEG-01細胞增殖能力的影響A.Cell liquid culture;B.Colony-forming cell assayFig 2 The effect of RhoB on MEG-01 cell proliferation
2.3 RhoB抑制CML患者CD34+細胞的增殖
為了檢測RhoB能否抑制CML病患CD34+細胞的增殖,隨機選取3個CML初診患者的骨髓樣本并純化出CD34+細胞進行集落生成實驗。結果如圖3所示,過表達RhoB后3個樣本的集落生成能力與Venus組相比均明顯受到抑制,平均抑制率約為90%。
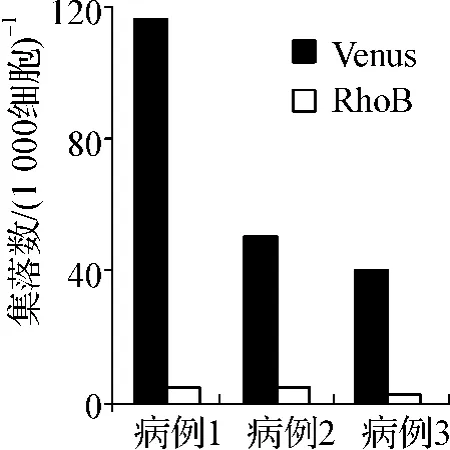
圖3 RhoB對CML患者CD34+細胞增殖能力的影響Fig 3 The effect of RhoB on the proliferation of CD34+cells from CML patients
2.4 RhoB抑制MEG-01細胞生長需要脂類修飾
為了驗證RhoB在MEG-01細胞中行使的抑制功能是否與其蛋白的脂類修飾相關,我們構建了RhoB的脂類修飾缺失變體RhoBC193G。結果發現,過表達RhoBC193G后細胞生長速度幾乎與Venus組細胞一致,細胞總數無明顯差異(P>0.05,圖4A)。此外,RhoBC193G與Venus組相比集落生成能力也無顯著差異(P >0.05,圖4B)。
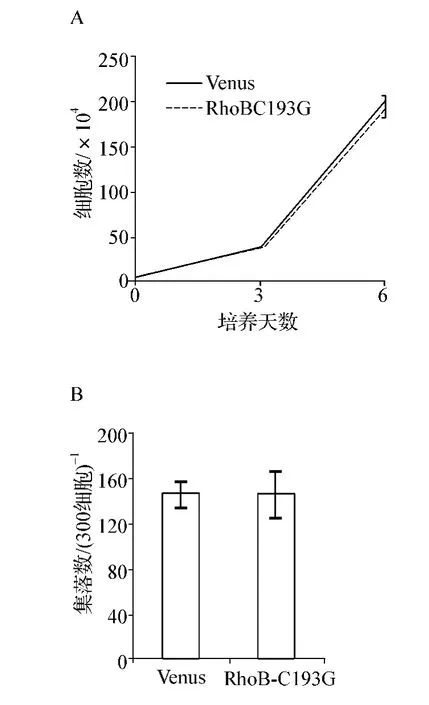
圖4 RhoB抑制功能與其脂類修飾相關性A.Cell liquid culture;B.Colony-forming cell assayFig 4 The association of the inhibitory effect of RhoB with its lipid modification
2.5 F修飾或GG修飾抑制MEG-01細胞的增殖
為了檢測RhoB在MEG-01細胞中行使其抑制作用與哪一種脂類修飾相關,我們在細胞中過表達只進行F修飾或只進行GG修飾的RhoB變體。結果發現,這兩種脂類修飾變體單獨過表達都可以顯著抑制細胞的增殖(P<0.01,圖5A)和集落生成能力(P<0.05,圖5B)。
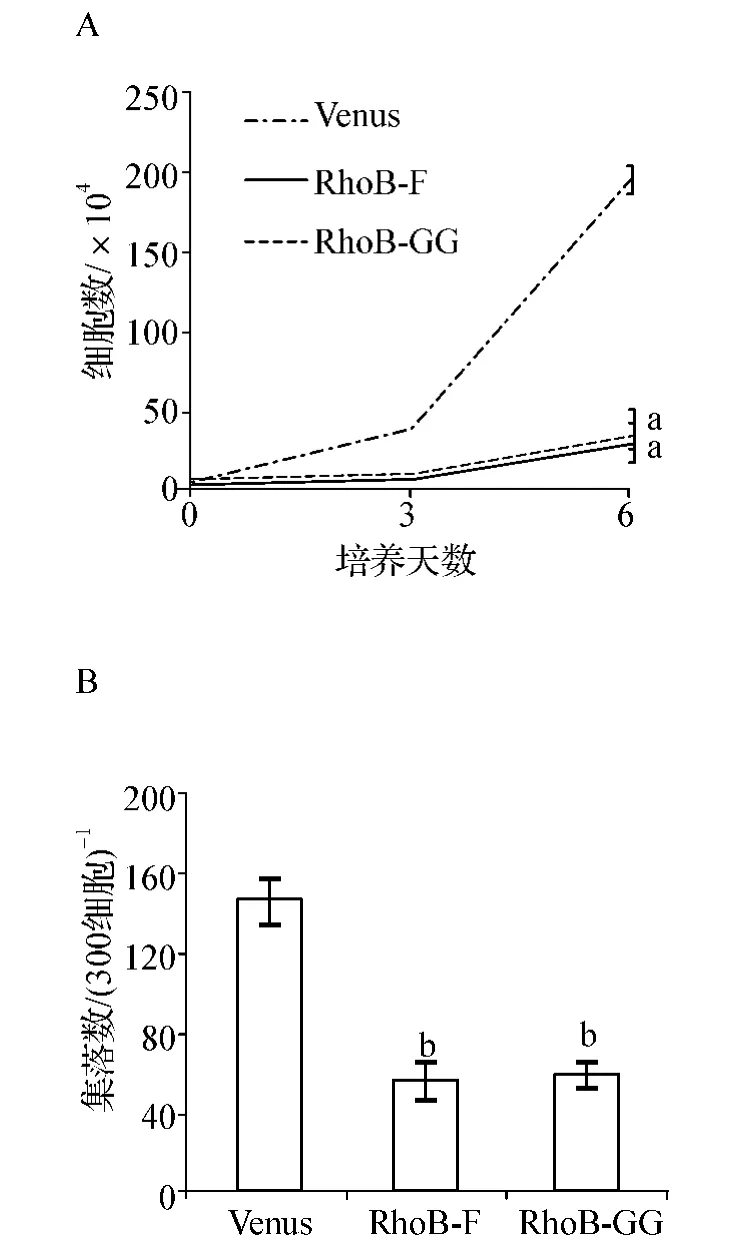
圖5 單一脂類修飾對MEG-01細胞增殖的影響A.Cell liquid culture;B.Colony-forming cell assayFig 5 The effect of mono-lipid modification on the proliferation of MEG-01cells
2.6 RhoB 阻滯MEG-01細胞于G2/M期
為了初步探索RhoB抑制MEG-01細胞生長的機制,我們利用流式細胞儀檢測過表達RhoB及其變體后MEG-01細胞的生長周期進程。結果(圖6)發現,分別過表達RhoB或RhoB-F、RhoB-GG后,處于G0/G1期的細胞與Venus組相比明顯減少(RhoB 34.58%±2.29%,RhoB-F 34.65% ±0.22%,RhoB-GG 39.71% ±1.12%,Venus 46.68% ±0.96%,P<0.05);而處于G2/M期的細胞則比Venus組顯著增多(RhoB 30.16% ±2.05%,RhoB-F 33.3% ±1.58%,RhoB-GG 28.75% ±0.50%,Venus 18.21% ±1.59%,P<0.05)。相比之下,RhoBC193G組無論是處于G0/G1期(47.32% ±0.61%)還是 G2/M期(18.11% ±0.84%)的細胞百分比基本與Venus組一致。這些結果說明RhoB將MEG-01細胞阻滯在G2/M期。
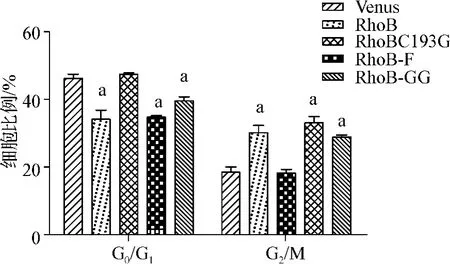
圖6 RhoB及其脂類修飾變體對MEG-01細胞周期的影響Fig 6 The effect of RhoB and its mutants on the cell cycle of MEG-01 cells
3 討 論
Rho家族成員中RhoB較為特殊。許多Rho蛋白在惡性轉化方面起正向調控作用,而RhoB的功能卻恰恰相反。研究發現RhoB在尿路上皮細胞癌、鱗狀細胞癌中表達顯著降低,并且在肺部腫瘤惡化過程中伴隨有RhoB基因的丟失[8-9]。RhoB基因敲除小鼠出生頻率、發育和壽命都正常,但是使用DMBA/TPA進行化學處理誘導形成皮膚腫瘤時,這些小鼠的成瘤率更高[10]。利用裸鼠皮下成瘤模型,在卵巢癌細胞中過表達RhoB可以抑制腫瘤的生長[11]。這些結果說明RhoB在實體瘤中可能是腫瘤抑制基因。但是RhoB是否在惡性血液病CML中同樣行使抑制功能卻不得而知。通過細胞增殖和集落生成實驗發現,CML細胞中過表達RhoB后細胞的增殖受到明顯抑制。結果表明CML細胞中RhoB依然行使抑制功能。
RhoB表達水平因細胞周期而異,RhoB轉錄本半衰期是30 min,這要遠遠比Rho蛋白家族其它成員短[12]。這就表明RhoB的功能要求它的表達受到嚴格的調控。其中一項重要的調控就是RhoB蛋白的脂類修飾,細胞中的RhoB蛋白被特異性地進行了兩種脂類修飾,F修飾和GG修飾。不同類型的脂類修飾可能導致RhoB蛋白在質膜、核膜、晚期內涵體等亞細胞水平不同的定位。RhoB-F蛋白大部分定位在質膜上,RhoB-GG蛋白則主要定位在晚期內涵體中[13]。一些研究表明RhoB-F、RhoB-GG這兩種脂類修飾形式具有不同的生物學功能。在Ras轉化的小鼠NIH3T3成纖維細胞中,RhoB和RhoB-GG起到抗轉化、抗增殖、誘導凋亡的作用[14]。而RhoB-F能夠選擇性地維持內皮細胞和經輻照處理的細胞的存活[15-16]。另外,RhoB,RhoB-F和RhoB-GG能抑制胰腺癌、宮頸癌細胞的生長[5]。本研究中,我們在MEG-01細胞中過表達RhoBC193G,發現失去脂類修飾后,細胞增殖與Venus組相比并無明顯差異,表明脂類修飾是RhoB抑制MEG-01細胞增殖所必需的。MEG-01細胞中分別過表達RhoB-F和RhoB-GG后細胞增殖也同樣受到抑制,表明單種脂類修飾仍然能夠維持RhoB在MEG-01細胞中的抑制功能。另外,細胞周期檢測發現RhoB,RhoB-F,RhoB-GG都可將MEG-01細胞阻滯在G2/M期,而RhoBC193G過表達的細胞周期進程卻與Venus組細胞一致,表明RhoB蛋白的脂類修飾影響MEG-01細胞的周期進程,進而抑制細胞的增殖。
綜上所述,本研究的功能學實驗結果和細胞機制分析說明RhoB抑制CML細胞的生長,并且RhoB蛋白進行任何一種脂類修飾均能在CML細胞中行使抑制功能,同時經過兩種脂類修飾或是單種脂類修飾的RhoB均可阻滯CML細胞于G2/M期。關于RhoB如何在MEG-01細胞中行使抑制功能進一步的分子機制還不明確,深入探索其分子機制將有助于CML治療方法的研究和靶向藥物的篩選。
[1]SHERBENOU D W,DRUKER B J.Applying the discovery of the Philadelphia chromosome[J].J Clin Invest,2007,117:2067-2074.
[2]MULLOY J C,CANCELAS J A,FILIPPI M D,et al.Rho GTPases in hematopoiesis and hemopathies[J].Blood,2010,115:936-947.
[3]PRENDERGAST G C.Actin'up:RhoB in cancer and apoptosis[J].Nat Rev Cancer,2001,1:162-168.
[4]HUANG M,PRENDERGAST G C.RhoB in cancer suppression[J].Histol Histopathol,2006,21:213-218.
[5]CHEN Z,SUN J,PRADINES A,et al.Both farnesylated and geranylgeranylated RhoB inhibit malignant transformation and suppress human tumor growth in nude mice[J].J Biol Chem,2000,275:17974-17978.
[6]吳平平,蘇昀,金治,等.RhoA蛋白通路在DLC-1基因調控人結腸癌HT29細胞周期中的作用及其機制[J].東南大學學報:醫學版,28(4):247-251.
[7]曹倪豪,陳明.BTG1在腎透明細胞癌組織中的表達及對786-O細胞株增殖和凋亡的影響[J].東南大學學報:醫學版,2011,30(4):583-587.
[8]VOLANIS D,ZARAVINOS A,KADIYSKA T,et al.Expression profile of Rho kinases in urinary bladder cancer[J].J BUON,2011,16:511-521.
[9]MAZIERES J,ANTONIA T,DASTE G,et al.Loss of RhoB expression in human lung cancer progression[J].Clin Cancer Res,2004,10:2742-2750.
[10]LIU A X,RANE N,LIU J P,et al.RhoB is dispensable for mouse development,but it modifies susceptibility to tumor formation as well as cell adhesion and growth factor signaling in transformed cells[J].Mol Cell Biol,2001,21:6906-6912.
[11]COUDERC B,PRADINES A,RAFII A,et al.In vivo restoration of RhoB expression leads to ovarian tumor regression[J].Cancer Gene Ther,2008,15:456-464.
[12]ZALCMAN G,CLOSSON V,LINARES-CRUZ G,et al.Regulation of Ras-related RhoB protein expression during the cell cycle[J].Oncogene,1995,10:1935-1945.
[13]WHERLOCK M,GAMPEL A,FUTTER C,et al.Farnesyltransferase inhibitors disrupt EGF receptor traffic through modulation of the RhoB GTPase[J].J Cell Sci,2004,117:3221-3231.
[14]MAZIERES J,TILLEMENT V,ALLAL C,et al.Geranylgeranylated,but not farnesylated,RhoB suppresses Ras transformation of NIH-3T3 cells[J].Exp Cell Res,2005,304:354-364.
[15]ADINI I,RABINOVITZ I,SUN J F,et al.RhoB controls Akt trafficking and stage-specific survival of endothelial cells during vascular development[J].Genes Dev,2003,17:2721-2732.
[16]MILIA J,TEYSSIER F,DALENC F,et al.Farnesylated RhoB inhibits radiation-induced mitotic cell death and controls radiation-induced centrosome overduplication[J].Cell Death Differ,2005,12:492-501.
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
文苑(2018年22期)2018-11-19 02:54:14
中國科技論壇(2017年7期)2017-07-25 08:49:53
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
