橘紅色花菜薹突變體的發(fā)現(xiàn)和研究
2013-02-24 09:01:30
中國蔬菜 2013年4期
(北京市農(nóng)林科學(xué)院蔬菜研究中心,北京 100097)
菜薹〔Brassica campestrisL.ssp.chinensis(L.)Makino var.utilisTsen et Lee〕別名菜心、綠菜薹、菜尖,起源于我國南部,在廣東、廣西栽培歷史悠久,是易抽薹白菜經(jīng)長期選擇和栽培馴化的產(chǎn)物,并演化為不同的類型和品種。菜薹質(zhì)地柔軟,風(fēng)味可口,營養(yǎng)豐富,不僅在南方地區(qū)普遍種植,近年來北方地區(qū)也有栽培。
突變體在植物品種改良上具有重要的應(yīng)用價值。一方面,通過基因突變產(chǎn)生的突變體,可以創(chuàng)造出新的種質(zhì)資源,增加物種的遺傳多樣性;另一方面,突變體作為標(biāo)記性狀,可以輔助辨別親本和一代雜種,顯著提高商品種子的純度和質(zhì)量。同時,突變體在基因定位、克隆和基因表達(dá)、調(diào)控等分子生物學(xué)研究領(lǐng)域也具有重要意義。
菜薹的花色以黃色為主,但也存在其他花色。2011年5月,筆者在1份菜薹育種材料F3群體中發(fā)現(xiàn)了1株突變體,其花色為橘紅色。眾所周知,橘紅心大白菜的花色為橘紅色,是一種特殊的大白菜類型,其資源來源于日本。研究發(fā)現(xiàn),大白菜橘紅心球色為質(zhì)量性狀,控制橘紅心球色的基因?yàn)殡[性單基因or(張德雙 等,2003)。橘紅心大白菜具有較高的營養(yǎng)價值,VC含量306.0 mg·kg-1,β-胡蘿卜素含量2.6 mg·kg-1(FW),居所有類型大白菜品種之首,尤其是β-胡蘿卜素含量,更是普通大白菜品種的6倍以上(張德雙 等,2004)。
劉秀村等(2003)應(yīng)用大白菜DH 群體,獲得了1個與橘紅心大白菜球色基因緊密連鎖的RAPD 分子標(biāo)記OPB01845,遺傳距離為3.8 cM,并將其轉(zhuǎn)化為SCAR 標(biāo)記SCB01-845。隨后,又發(fā)現(xiàn)了1個與橘紅心球色基因連鎖的SCAR 標(biāo)記(王國臣 等,2007;Zhang et al.,2008),并最終將or基因定位在大白菜1號染色體的長臂末端(于拴倉 等,2007)。Feng 等(2010)獲得了與橘紅心基因連鎖的3個SSR 標(biāo)記,其中sau(C)586、syau19 在or基因同側(cè),遺傳距離分別為1.6 cM 和1.3 cM;syau15位于另一側(cè),遺傳距離為3.3 cM。
本試驗(yàn)的目的是研究菜薹橘紅色花的遺傳規(guī)律,并應(yīng)用與大白菜橘紅心球色基因連鎖的SSR分子標(biāo)記對控制菜薹橘紅色花的基因進(jìn)行分析,鑒定菜薹的橘紅色花基因與大白菜的橘紅心球色基因or是否相同,為今后選育橘紅色花菜薹一代雜種和獲得新的橘紅心大白菜種質(zhì)資源奠定基礎(chǔ)。
1 材料與方法
1.1 材料
橘紅色花菜薹11A-47 是筆者于2011年5月發(fā)現(xiàn)的1株突變體,其葉色為翠綠色,葉形為近橢圓 形,莖翠綠色,分枝較旺,花色為橘紅色,鮮艷且較深(圖1)。黃色花菜薹品種為聯(lián)記特選34號甜菜心,該材料已自交7代,遺傳性狀穩(wěn)定、純合,葉色深綠,葉形為圓形,莖綠色,分枝較旺,花色為黃色。供試材料均來源于北京市農(nóng)林科學(xué)院蔬菜研究中心白菜課題組。

圖1 橘紅色花菜薹突變體11A-47
1.2 方法
1.2.1 子葉顏色調(diào)查方法 2011年5月,在本中心對橘紅色花菜薹11A-47 進(jìn)行自交留種;同時以11A-47 為父本,與黃色花菜薹進(jìn)行雜交,獲得F1種子。2012年6月初,F(xiàn)1自交獲得F2,再以F1為母本分別與黃色花菜薹、11A-47 雜交,分別獲得BC1和BC1′。參考耿新翠等(2012)的方法并加以改進(jìn),調(diào)查子葉顏色:7月初,將P1、P2、F1、F2、BC1和BC1′6個世代的種子放置在4℃冰箱中,低溫遮光處理15 d后取出,用肉眼觀測子葉顏色,記錄并對數(shù)據(jù)進(jìn)行方差分析。然后,將6個世代種子分別播種到50 孔穴盤中,直到開花,調(diào)查并記錄各單株花瓣顏色。
1.2.2 根部和髓部顏色調(diào)查方法 2012年7月初,待11A-47 和黃色花菜薹的自交種子收獲結(jié)束后,拔取11A-47 與黃色花菜薹各20株,將菜薹根部洗凈,比較11A-47 與黃色花菜薹的根部顏色。再將根部橫切即獲得髓部,比較二者髓部顏色。
1.2.3 花蕾和花瓣顏色調(diào)查方法 在菜薹盛花期,比較11A-47 與黃色花菜薹的花蕾及花瓣顏色。
1.2.4 分子標(biāo)記方法 以來源于大白菜R9 連鎖群(即大白菜1號染色體)、并與橘紅心球色基因or連鎖的11對SSR 引物對黃色花大白菜、橘紅色花大白菜、黃色花菜薹和11A-47 進(jìn)行標(biāo)記分析。引物分別為Feng 等(2010)的5 對SSR 引物sau(N)033、sau(C)586、syau15、syau19、ENA21,本中心耐寒類蔬菜育種研究室設(shè)計的5 對SSR 引物ba14066、PD7413、PD1355、PD6142、PD7418 和Wang 等(2011)的1對SSR 引物BRID90153。
反應(yīng)體系(12.5 μL):1μL 50 ng 模板 DNA,0.6 μL dNTP(2.5 mmol·mL-1),1.25 μL 10×buffer(含Mg2+),上下游引物各0.8 μL(0.5 μmol·mL-1),0.15 μLTaq酶,7.9 μL ddH2O。PCR 反應(yīng)程序:95℃ 3 min;95℃ 30 s,56℃ 30 s,72℃ 45 s,共35個循環(huán);72℃延伸10 min。擴(kuò)增產(chǎn)物用3%超高分辨率瓊脂糖凝膠電泳檢測。
2 結(jié)果與分析
2.1 橘紅色花菜薹突變體11A-47 的植物學(xué)性狀
2.1.1 橘紅色花菜薹子葉顏色的遺傳規(guī)律 橘紅色花菜薹突變體11A-47 6個世代各單株子葉顏色和卡方檢測結(jié)果見表1和圖2。親本P1為普通黃色花菜薹,子葉顏色全部為綠色;親本P2為橘紅色花菜薹,子葉顏色全部為桔紅色,二者雜交F1的子葉顏色均為綠色,說明橘紅色花對黃色花為隱性;F2世代子葉顏色出現(xiàn)了分離,橘紅色與綠色單株的分離比例符合1∶3,χ2=1.938 9 <χ20.05=3.841;BC1世 代中橘紅色單株數(shù)量很少,約占4.36%;BC1′世代中橘紅色與綠色單株的分離比例符合1∶1,χ2=1.369 7 <χ20.05=3.841。
2.1.2 橘紅色花菜薹根部和髓部顏色 11A-47 與黃色花菜薹根部、髓部顏色見圖3和圖4。11A-47 的根部、髓部顏色均表現(xiàn)為橘紅色,而黃色花菜薹的根部、髓部顏色均表現(xiàn)為白色。據(jù)報道,絕大多數(shù)類胡蘿卜素呈黃色、橘紅色或紅色,是許多花、果實(shí)和胡蘿卜根呈現(xiàn)各種顏色的原因(Bartley & Scolnik,1995)。橘紅心大白菜葉球之所以呈現(xiàn)橘紅色,就是由于高含量β-胡蘿卜素等類胡蘿卜素的積累所致(Bartley & Scolnik,1995;張華敏 等,2009)。本試驗(yàn)中,橘紅色花菜薹11A-47 的根部、髓部顏色均為橘紅色,充分表明這些器官積累了類胡蘿卜素。
2.1.3 橘紅色花菜薹花蕾和花瓣顏色 11A-47 與黃色花菜薹花蕾、花瓣顏色見圖5 和圖6。11A-47 的花蕾顏色為淺綠色,花瓣為桔紅色,花色鮮艷且較深。而黃色花菜薹花蕾顏色為綠色,花瓣為黃色。

表1 黃色花菜薹×11A-47 各世代子葉顏色和卡方檢測結(jié)果
2.2 SSR分子標(biāo)記擴(kuò)增結(jié)果
11對SSR 引物對黃色花大白菜、橘紅色花大白菜、黃色花菜薹和11A-47 的擴(kuò)增結(jié)果見圖7 和表2。其中,BRID90153、PD1355、sau(N)033、sau(C)586、syau15、syau19 等6 對引物可在11A-47 與橘紅色花大白菜基因型中擴(kuò)增出差異產(chǎn)物,而其余5 對引物則沒有擴(kuò)增出差異產(chǎn)物。由此可見,控制菜薹11A-47 橘紅色花的基因與控制大白菜橘紅心球色的基因可能不同。
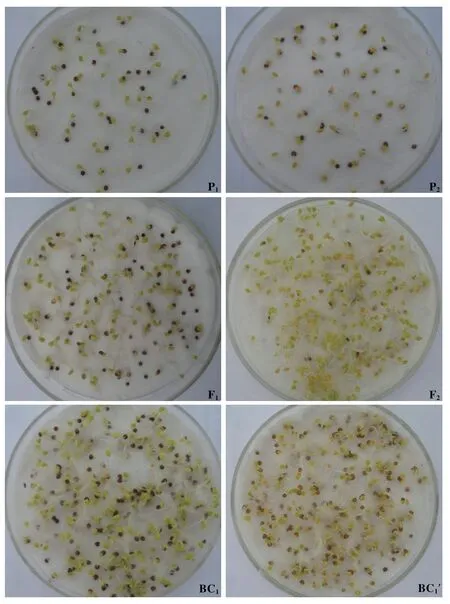
圖2 黃色花菜薹×11A-47 6個世代各單株子葉顏色

圖3 11A-47 與黃色花菜薹的根部顏色

圖4 11A-47 與黃色花菜薹的髓部顏色

圖5 11A-47 與黃色花菜薹的花蕾顏色

圖6 11A-47 與黃色花菜薹的花瓣顏色
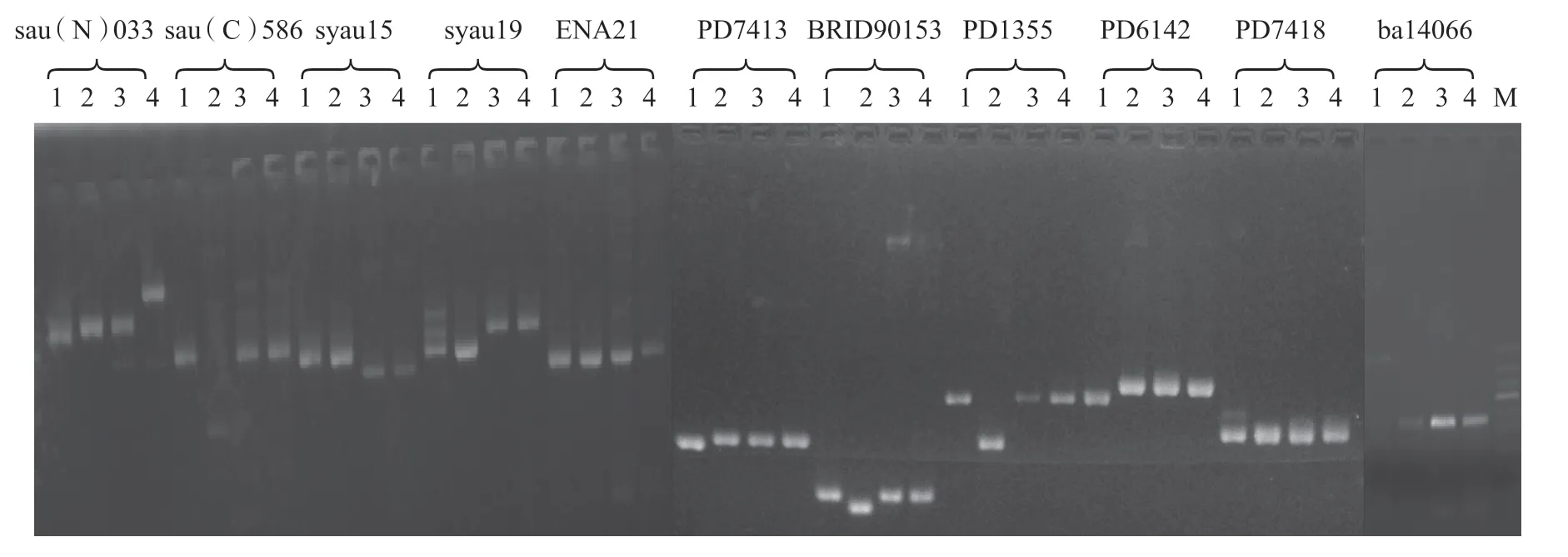
圖7 SSR 引物擴(kuò)增結(jié)果

表2 11對SSR 引物的擴(kuò)增結(jié)果
3 結(jié)論與討論
突變體的創(chuàng)建方法主要有自發(fā)突變、物理誘變、化學(xué)誘變,T-DNA、轉(zhuǎn)座子、反轉(zhuǎn)座子插入等,其中自發(fā)突變是自然界普遍存在的變異。在育種實(shí)踐中,一旦發(fā)現(xiàn)了具有某種有利性狀的突變體,通過研究其遺傳規(guī)律并加以應(yīng)用,最終可選育出具有該性狀的優(yōu)良品種。
3.1 11A-47 突變體的來源
菜薹花色發(fā)生自然變異的可能性比較大。筆者發(fā)現(xiàn)的橘紅色花菜薹突變體11A-47 來源于1份自交2 代的F3菜薹群體。11A-47 突變體除了花色為橘紅色外,其余性狀與黃色花菜薹相似,其葉色翠綠色,葉形近橢圓形,葉柄長,莖翠綠色,腋芽萌發(fā)能力較強(qiáng);且不具有菜薹與大白菜雜交后代產(chǎn)生的中間性狀,如葉翼等。因此,可以肯定地說,菜薹突變體11A-47 的橘紅色花基因不是來源于橘紅心大白菜,而是菜薹自然突變的產(chǎn)物。在調(diào)查菜薹育種材料性狀時,除了花色為橘紅色的突變體11A-47 外,筆者還發(fā)現(xiàn)了以白色為主且嵌有黃色的花色突變體,并可以穩(wěn)定遺傳。
3.2 BC1 群體中橘紅色子葉單株存在的原因
本試驗(yàn)是以子葉顏色來確定橘紅色花或黃色花單株的。BC1群體中存在極少數(shù)橘紅色子葉單株的可能原因主要有:① 在調(diào)查子葉顏色時,將個別原本為綠色子葉的單株誤定為橘紅色子葉;② 因F1花粉去雄不徹底而自交,產(chǎn)生少部分F2種子,混在BC1回交群體中。為了重復(fù)本試驗(yàn),筆者還播種了另外2份BC1回交群體,仍然發(fā)現(xiàn)具有橘紅色子葉的個別單株,橘紅色子葉與綠色子葉的比例分別是6∶85 和2∶101。同時,本試驗(yàn)還發(fā)現(xiàn),在盛花期,BC1回交群體依然存在橘紅色花的植株,因此,今后的重點(diǎn)是嚴(yán)格拔除F1雄蕊,重新獲得BC1群體,并進(jìn)一步驗(yàn)證上述試驗(yàn)結(jié)果。
3.3 菜薹橘紅色花基因與大白菜橘紅心球色or基因的異同
控制菜薹橘紅色花的基因與大白菜橘紅心球色or基因的遺傳規(guī)律一致,即菜薹橘紅色花、大白菜橘紅心球色均為質(zhì)量性狀,由1對隱性等位基因控制。本試驗(yàn)應(yīng)用11對與大白菜橘紅心球色基因連鎖的SSR分子標(biāo)記對11A-47 進(jìn)行分析,其中6 對引物擴(kuò)增的產(chǎn)物能夠準(zhǔn)確區(qū)分11A-47、橘紅心球色大白菜,但這6 對引物也可在黃色花大白菜或黃色花菜薹中擴(kuò)增出相同的產(chǎn)物,即無法將黃色花大白菜、橘紅色花大白菜、黃色花菜薹和11A-47 同時區(qū)分開。由目前的研究結(jié)果初步分析可見,控制菜薹11A-47 橘紅色花的基因與控制大白菜橘紅心球色的基因可能不同。下一步,將對控制11A-47 橘紅色花的or基因進(jìn)行染色體初步定位,進(jìn)一步證明二者間的關(guān)系。
本試驗(yàn)還發(fā)現(xiàn),PD7413、PD7418、sau(N) 033、syau15、syau19 和ENA21 等6 對引物雖然已被成功用于區(qū)分黃色花和橘紅色花大白菜基因型,但在本試驗(yàn)中并沒有在橘紅色花大白菜基因型中擴(kuò)增出差異產(chǎn)物,可能是由于試驗(yàn)采用的電泳檢測方法(丙烯酰胺凝膠和超高分辨率瓊脂糖凝膠)不同或這些引物與大白菜橘紅心球色or基因連鎖的不緊密等原因造成的。
3.4 菜薹橘紅色花基因的應(yīng)用
筆者已經(jīng)開始利用11A-47 和白心大白菜進(jìn)行雜交,希望獲得表現(xiàn)更優(yōu)的大白菜橘紅心育種資源,并進(jìn)一步育成橘紅心新品種。另外,菜薹的橘紅色花基因可以使其根部和髓部變?yōu)殚偌t色,因此采用生物技術(shù)方法有可能將該基因轉(zhuǎn)到蘿卜等食用根部器官的蔬菜中,創(chuàng)造出更多具有橘紅色性狀的蔬菜新資源。
耿新翠,沈向群,侯華,洪雅婷,馬士芳.2012.橘紅心大白菜形態(tài)標(biāo)記的遺傳及穩(wěn)定性研究.中國蔬菜,(6):21-26.
劉秀村,張鳳蘭,張德雙,楊建平,王美,徐家炳.2003.與大白菜桔紅心基因連鎖的RAPD 標(biāo)記.華北農(nóng)學(xué)報,18(4):51-54.
王國臣,張鳳蘭,余陽俊,張德雙,趙岫云.2007.與大白菜橘紅心基因緊密連鎖的SCAR 標(biāo)記.園藝學(xué)報,34 (1):217-220.
于拴倉,于仁波,戚佳妮,張鳳蘭,余陽俊,張德雙,趙岫云.2007.大白菜桔紅心or基因的定位研究.中國遺傳學(xué)會植物遺傳和基因組學(xué)專業(yè)委員會2007年學(xué)術(shù)研討會.金華.
張德雙,張鳳蘭,徐家炳.2003.大白菜花色和球色遺傳規(guī)律的研究.華北農(nóng)學(xué)報,18(2):81-84.
張德雙,徐家炳,張鳳蘭.2004.不同球色大白菜主要營養(yǎng)成分分析.中國蔬菜,(3):37.
張華敏,武云霞,張魯剛,張亞麗.2009.達(dá)草滅對橙色大白菜愈傷組織中類胡蘿卜素含量的影響.植物生理學(xué)通訊,45(4):340-344.
Bartley G E,Scolnik P A.1995.Plant carotenoid:pigments for photoprotection,visual attraction,and human health.Plant Cell,7:1027-1038.
Feng H,Li Y F,Liu Z Y,Liu J.2010.Mapping ofor,a gene conferring orange color on the inner leaf of the Chinese cabbage(Brassica rapaL.ssp.pekinensis).Molecular Breeding,DOI:10.1007/s11032-010-9542-x.
Wang Y,Sun S L,Liu B,Wang H,Deng J,Liao Y C,Wang Q,Cheng F,Wang X W,Wu J.2011.A sequence-based genetic linkage map as a reference forBrassica rapapseudochromosome assembly.BMC Genomics,DOI:10.1186/1471-2164-12-239.
Zhang F L,Wang G C,Wang M,Liu X C,Zhao X Y,Yu Y J,Zhang D S,Yu S C.2008.Identification of SCAR markers linked toor,a gene inducing beta-carotene accumulation in Chinese cabbage.Euphytica,164:463-471.
