兩用核不育系P88S愈傷組織的誘導及其遺傳轉化研究
2013-09-04 03:40:12袁定陽夏石頭
湖南農業科學 2013年13期
關鍵詞:水稻
崔 看 ,成 平 ,陳 錦 ,袁定陽 ,夏石頭
(1.湖南農業大學生物科學技術學院,植物激素與生長發育湖南省重點實驗室,湖南 長沙 410128;2.湖南省農業信息與工程研究所,湖南 長沙 410125;3.雜交水稻國家重點實驗室,湖南 長沙 410125)
水稻(Oryza sativa L.)是人類最重要的糧食作物之一,傳統遺傳育種技術在提高水稻產量和改善米質等方面做出了巨大貢獻[1]。但由于生態環境惡化和水稻種質資源匱乏的限制,傳統的遺傳改良方法已經不能夠滿足現代農業對新型種質資源的需求。P88S是由湖南雜交水稻研究中心羅孝和研究員在2003年利用雙向系選(在長沙長日低溫的條件下選不育,在海南短日低溫下選相對可育)和溫光加再生強化選擇而選育出的異交結實率高、配合力強的超級雜交水稻兩用核不育系,在超級稻生產上得到了廣泛應用,但其對稻瘟病抗性較弱[2-3]。利用基因工程技術將植物抗病性狀相關基因導入水稻基因組中,可以作為獲得新種質資源的一種重要手段,在實現超級稻高產、優質、多抗、穩產目標中發揮重要作用[4]。文章以兩用核不育系P88S為材料,研究了其愈傷組織的誘導及植物抗病基因snc1在其愈傷組織中的遺傳轉化,以期通過誘導、分化和繼代培養獲得高抗病超級稻新種質。
1 材料與方法
1.1 材料
超級雜交水稻兩用核不育系P88S,由湖南雜交水稻研究中心羅孝和研究員提供;遺傳轉化載體為夏石頭教授課題組改造的pG229質粒,含有Kan+與Basta雙重抗性標記;供體菌為農桿菌EHA105,功能獲得型突變基因Atsnc1[5],由加拿大英屬哥倫比亞大學Michael Smith Laboratories李昕教授實驗室惠贈。
1.2 方法
1.2.1 P88S愈傷組織的誘導 采取完全隨機化設計,參照文獻[6-7]選用4種誘導培養基MB、N6、NB和NMB進行愈傷組織誘導與繼代培養,將P88S成熟胚分別接種于 MB、N6、NB、NMB 4種培養基上進行愈傷組織的誘導,2,4-D濃度分別設為2.0、2.5、3.0 mg/L,6-BA 的濃度分別設為 0.1、0.2、0.3 mg/L,并添加0.5 g/L酸水解酪蛋白(CH)和脯氨酸(Pro)。誘導7 d或繼代培養12 d后,分別統計芽長、愈傷組織重量和誘導率等性狀,以篩選出最適合P88S愈傷組織誘導和繼代培養的最適培養基與最適生長物質濃度配比。
1.2.2 遺傳轉化植株的獲得與檢測 農桿菌的浸染轉化、抗性愈傷組織的篩選、轉化植株的檢測按照成平等[8]方法,轉化植株的檢測采用試劑盒(北京康為世紀生物科技有限公司),提取轉化植株的DNA,進行PCR檢測。正向引物序列為:GTGGAGTTCCCATCTGAACATC,反向引物序列為:CCCATTTTGATTGCTGGAAAG。
2 結果與分析
2.1 P88S愈傷組織誘導培養母液的篩選
將 P88S的成熟胚分別接種于 MB、N6、NB、NMB培養基上(含3.0 mg/L 2,4-D+0.2 mg/L 6-BA+0.5 g/L酸水解酪蛋白+0.5 g/L脯氨酸)誘導7 d后,觀察和統計各培養基上愈傷組織的顏色、重量和芽長等。試驗結果如表1所示,P88S在MB、NMB、N6培養基上誘導時,愈傷組織顏色淡黃,顆粒圓潤飽滿,而在NB培養基上誘導時,愈傷組織顆粒有褐化現象且顆粒較小。如圖1所示,P88S在MB培養基上P88S誘導的芽最長,為2.797 cm,比在 N6、NB、NMB 培養基上分別高出 0.749、0.821、0.887 cm,差異達到極顯著水平;在NB培養基上誘導的P88S愈傷組織重量最輕,為0.004 6 g,極顯著低于其余3種培養基上誘導的愈傷組織;NMB培養基的誘導率最高,達95.66%,極顯著高于MB培養基的愈傷組織誘導率。綜合考慮各因素,NMB培養基是最適合P88S愈傷組織誘導的培養基。
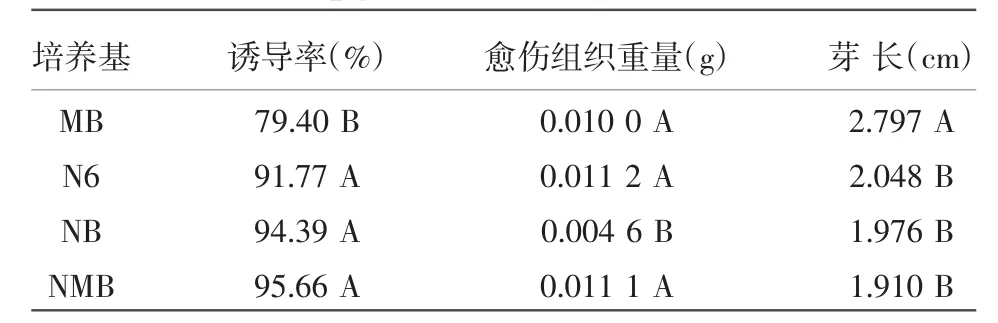
表1 P88S愈傷組織在不同培養基上誘導的芽長、愈傷組織重量和誘導率
2.2 P88S愈傷組織誘導培養基中植物生長物質濃度的選擇
不同濃度2,4-D與6-BA對P88S愈傷組織誘導的影響如表2所示,不同濃度的2,4-D對于P88S芽長的影響不顯著,而對于愈傷組織重量的影響差異達到極顯著水平,當2,4-D質量濃度為3.0 mg/L時,P88S愈傷組織重量最大,平均達0.011 g,極顯著高于其他2個濃度處理的愈傷組織重量。當添加3.0 mg/L的2,4-D時,誘導率達到94.70%,極顯著高于添加2.5 mg/L 2,4-D時的誘導率,表明2,4-D質量濃度對P88S愈傷組織誘導率的有重要影響。而6-BA對芽長的影響明顯,濃度為0.3 mg/L 6-BA處理的芽長極顯著高于其他2個濃度處理的芽長,其愈傷組織誘導率與0.2 mg/L 6-BA處理的愈傷組織誘導率相近,但顯著高于0.1 mg/L 6-BA處理的愈傷組織誘導率。綜合考慮各因素,確定2,4-D最適質量濃度為3.0 mg/L,6-BA最適質量濃度為0.2 mg/L。
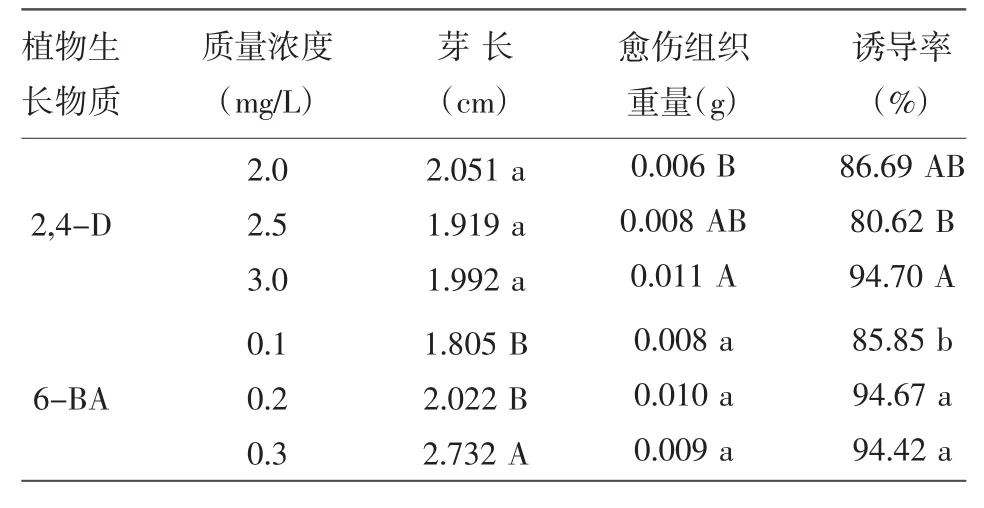
表2 2,4-D和6-BA對P88S愈傷組織誘導的影響
2.3 酸水解酪蛋白和脯氨酸對水稻成熟胚愈傷組織誘導的影響
根據愈傷組織對滲透壓的要求,在試驗中添加了 0.5 g/L酸水解酪蛋白(CH)和脯氨酸(Pro),統計結果表明,添加CH和Pro的培養基,芽長為2.103 cm,愈傷組織重量為0.011 2 g,誘導率為94.75%;不添加CH和Pro的培養基,芽長為2.690 cm,愈傷組織重量為0.006 9 g,誘導率為91.83%。與不添加CH和Pro相比,添加CH和Pro的培養基芽長較短,其誘導率稍高,并能顯著提高愈傷組織的質量。且添加CH和Pro后,愈傷組織生長旺盛,顆粒圓潤,顏色嫩黃鮮艷;而不添加CH和Pro,愈傷組織生長狀態較差,顆粒干癟較小,且有褐化現象。
2.4 P88S愈傷組織最適繼代培養基的確定
愈傷組織一般都要繼代1次,即培養約12d后才進行轉化,且轉化前會將愈傷組織一分為二,造成一定創傷以有利于農桿菌的浸染。在對同一批愈傷組織進行MB、N6、NB、NMB培養基繼代培養,并觀測繼代培養生長和增重情況后,發現NMB培養基是P88S愈傷組織繼代培養的最適培養基,其增重最大,為0.233 g,增重達185%,極顯著高于N6和NB培養基上愈傷組織的增重(圖1)。
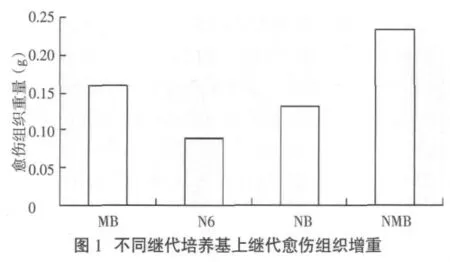
2.5 P88S愈傷組織最適分化培養基的確定
分化培養的目的是讓愈傷組織重新分化成小苗,比較研究了DL和NMB兩種母液對P88S愈傷組織分化的影響,試驗結果表明,不同碳源對愈傷組織分化有一定影響,以麥芽糖作為碳源時,分化啟動較快,愈傷組織生長較迅速,7d左右就可見新的愈傷組織小顆粒,且愈傷組織緊致,綠豆大小的顆粒,出綠點時間為10~15d;以蔗糖作為碳源時,愈傷組織生長緩慢,一般2個星期左右才會有新愈傷組織,且愈傷組織松散,一夾即碎,出綠點時間為15~20d。P88S在DL分化培養基中培養25d后分化出芽,而在NMB分化培養基中培養25d后,愈傷組織有褐化死亡的跡象,且僅有部分愈傷組織出現綠點。因此,選用麥芽糖為碳源的DL分化培養基作為P88S的最適分化培養基。
2.6 遺傳轉化植株的獲得與檢測
當分化培養基中綠芽長到長于2 cm后,將其從愈傷組織上剝離,轉入生根培養基中。大約2周幼苗就長出完整的根系,加水煉苗2~3d后將其移出,洗凈掉根上的培養基,剪掉變黃的葉子,移栽到稻田土中(其轉化流程見圖2)。遺傳轉化植株驗證時,取移栽到稻田土中T0代轉化植株葉片,用試劑盒法提取小量DNA,利用snc1基因的特異引物進行PCR擴增檢測5株P88S轉化植株,結果顯示P88ST0-5中檢測到了目的基因snc1的存在(圖2中h)。
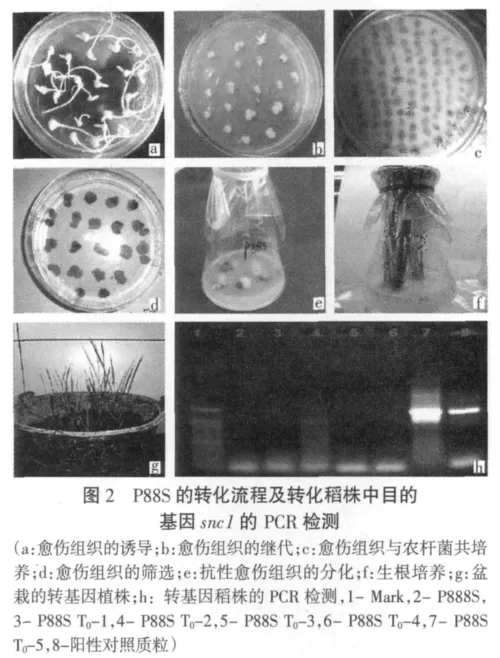
3 結論與討論
以超級雜交水稻兩用核不育系P88S為材料,研究發現P88S成熟胚愈傷組織的最適誘導和繼代培養基是NMB培養基,其中2,4-D和6-BA的最適濃度分別為3.0 mg/L和0.2 mg/L,而添加0.5 g/L的酸水解酪蛋白和脯氨酸有利于愈傷組織的誘導。P88S成熟胚愈傷組織的最適分化培養基是DL分化培養基,麥芽糖為抗性愈傷組織分化的最適碳源。并通過農桿菌介導遺傳轉化法共獲得5株轉化稻株,經PCR檢測其中1株含目的基因s nc1。
擬南芥 SNC1(suppressor of npr1-1,constitutive1)基因[5]是一種重要的水楊酸誘導型廣譜抗病基因,該基因功能獲得型突變導致snc1突變體植株組成性表達病程相關蛋白,能夠組成性激活植物的抗病機制,并對細菌病原體Pseudomonas syringae pv maculicola ES4326(P.s.m.ES4326)和卵菌病原體 Hyaloperonospora arabidopsidis Noco2(H.a.Noco2)等病原物產生廣譜抗性。因為Atsnc1基因是一種獲得功能型突變基因,可以組成性表達致病機理相關(PR)基因,從而激活抗病性反應,P88S轉化稻株中snc1基因是否有同樣的生理功能,還有待進一步研究。
[1]彭雪明,張小平,周文新,等.中國超級稻研究進展與栽培研究展望[J].作物研究,2009(1):1-6.
[2]張志剛,羅孝和,羅 珊.水稻光溫敏核不育系P88S的選育與應用[J].新品種推廣與應用,2008,27(11):123-125.
[3]羅孝和,夏石頭,羅 珊,等.兩用核不育系及其超級稻研究[J],雜交水稻,2010,(25):59-63.
[4]沈志忠.近代中國水稻品種改良探析[J].江海學刊,2010,(6):164-170.
[5]Zhu Z H,Xu F,Zhang YX,et al.Arabidopsis resistance protein SNC1 activates immune responses through association with a transcriptional corepressor[J].Proceedings of the National Academy of Sciences,2010,107(31):13960-13965.
[6]Lin Y J,Zhang Q F.Optimizing the tissue culture conditions for high efficiency transformation of indica rice[J].Plant Cell Reports,2005,(23):540-547.
[7]李明浩,陳 煒,邢莉萍,等.普通小麥品種Alondra’s遺傳轉化體系的建立[J].植物學報,2010,(4):466-471.
[8]成 平,陳 錦,袁定陽,等.植物抗病基因snc1在超級稻父本0293中的遺傳轉化[J].湖南農業大學學報(自然科學版),2012,38(3):267-270.
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
