蛋白質組學技術在細胞信號轉導研究中的應用
2013-10-24 09:18:20王瑋鵬苗芳芳武丹丹楊軍王志鋼
生物技術通報 2013年11期
王瑋鵬 苗芳芳 武丹丹 楊軍 王志鋼
(1.內蒙古大學生命科學學院,呼和浩特 010021;2.內蒙古呼和浩特市環境科學研究所,呼和浩特 010030)
隨著蛋白分離技術與質譜技術的發展,結合基因組學和轉錄組學研究獲得的海量信息,對蛋白質的研究已從蛋白質化學發展到蛋白質組學的研究階段,并成為21 世紀生命科學的重要技術支撐和戰略前沿。蛋白質組學中的蛋白質鑒定、定量以及相互作用的研究已實現了其對細胞信號轉導和相關通路機制的研究作出貢獻的早期設想。新的蛋白質組學技術在很大程度上推進了細胞信號轉導研究的深入,闡明了細胞信號轉導通路是通過蛋白質間相互作用(protein-protein interaction)從而實現信號傳遞的機制。利用蛋白質組學技術進一步確定各種信號轉導通路中的未知分子,實現對這些通路之間的聯系(cross-talk)及所構成的錯綜復雜的細胞內信號網絡系統更深入的認識是目前的研究熱點。
1 蛋白質組學的概念與技術
1.1 蛋白質組學的定義
1994 年澳大利亞Macquarie大學的Wilkins和Williams等在意大利的第一屆Siena會議上首次提出了蛋白質組(Proteome)這個概念,并最初定義為“一個基因組所表達的蛋白質”[1]。目前認為蛋白質組是指一個基因組所編碼的全部蛋白質,蛋白質組學是指利用直接測定和鑒別蛋白質的高通量方法,大規模研究蛋白質組的表達動力學和蛋白質相互作用。蛋白質組學是研究細胞和組織中全部蛋白質的科學,試圖通過蛋白質組學的研究從細胞水平及整體水平上研究蛋白質的組成及其變化規律,從而深入認識有機體的各種生理和病理過程。
1.2 蛋白質組學研究領域與技術
對蛋白質組的功能分析是功能基因組學的核心。蛋白質組學的研究一般可以劃分為3個領域:一是蛋白質鑒定和分析;二是不同生長條件下蛋白質的泛蛋白質組(proteome-wide)差異顯示;三是蛋白質-蛋白質相互作用。其運用的技術主要是蛋白質分離與鑒定技術,同時生物信息學技術也是蛋白質組學研究技術中不可或缺的部分。
傳統蛋白質研究方法的單一應用已經不能滿足目前對蛋白質組研究的要求,在已有方法的基礎上將其整合與創新,已經形成了系統的蛋白質組學分離與鑒定、蛋白質相互作用與修飾及定量研究技術。對蛋白質樣品的制備通常從樣品的預分級開始。常用的預分級方法如二維高效液相色譜、液相等電聚焦及膜電泳,亞細胞分級與單細胞水平樣品制備等,主要用以提高低豐度蛋白質的上樣量和檢測靈敏度,并且可以針對某一細胞器的蛋白質組進行研究。蛋白質的分離與鑒定是預分級處理后蛋白質組學研究中極為重要的一環,雙向凝膠電泳(two-dimensional gel electrophoresis,2-DE)和高效液相色譜(high performance liquid chromatography,HPLC) 是 目 前常用的方法,毛細管電泳(capillary electrophoresis,CE)則是經典電泳技術與現代微柱分離技術相結合的產物,具有極高的分辨率,而毛細管色譜(capillary electro chromatography,CEC)則同時具有毛細管電泳的高效分離性和HPLC的高效選擇性。
蛋白質鑒定的主要手段之一便是生物質譜(Biomass spectrometry,Bio-MS),Bio-MS是通過制備、分離、檢測氣相生物大分子來鑒定化合物的高通量技術,具有較高特異性與靈敏性。液質連用(liquid chromatography-mass spectrometry,LC-MS) 是 以 液相色譜作為分離系統,質譜作為檢測系統對復雜樣品進行實時分析的綜合技術,它綜合了液相色譜較寬的分離范圍特性與MS的高選擇性和高靈敏度的特點,能夠實現很好的鑒定效果。在此基礎上建立的液相色譜串聯質譜聯用(liquid chromatographytandem mass spectrometry,LC-MS/MS)技術則將液相色譜與串聯質譜相結合,檢測樣品即使在液相色譜難分離的情況下,只要通過多級質譜對目標化合物進行的中性碎片掃描,即可發現并突出混和物中的目標化合物,提供更精確的鑒定效果。此外,近年基體輔助激光解吸電離-飛行時間質譜技術(matrixassisted laser desorption ionization-time of flight-mass spectrometry,MALDI-TOF-MS)、Shotgun混 合 蛋 白鑒定技術及蛋白質從頭測序(De-novo)等技術在未知樣品分析及翻譯后修飾鑒定中得到了廣泛應用。同時,蛋白質定量技術近年也逐步發展起來,一是基于傳統雙向凝膠電泳及染色基礎上的定量,如雙向熒光差異凝膠電泳(two-dimensional fluorescence difference gel electrophoresis,DIGE),通過三色熒光染料分別對內標和生物樣本進行標記,再通過圖像分析準確地發現差異表達的蛋白;另一是基于質譜檢測技術的定量,包括采用差異同位素標記的相對定量技術,如ICAT(isotope-coded affinity tag)、SILAC(stable isotope labeling with amino acids in cell)和iTRAQ(isobaric tags for relative and absolute quantitation)等,以及含有特定同位素的目標肽段的絕對定量技術。值得注意的是,高通量的蛋白質組學技術的應用是與生物信息學技術的發展分不開的,蛋白質定性與定量測定結果的分析依賴于生物信息學建立起來的計算模型和數據庫,尤其是未知蛋白的結構和功能預測更離不開生物信息學的支持。
2 細胞信號轉導與分子調控
細胞生長調控是一個受多因素影響的復雜過程,它不僅受到時間和空間的限制,還受到營養條件和細胞內外環境條件的影響。盡管目前對細胞生長和細胞周期的調節機制了解得很少,但它已經成為了現代細胞生物學的研究重點,同時也取得了許多重大進展。細胞內存在許多不同的信號通路來調控細胞內外刺激所引發的反應,并介導細胞的代謝、增殖、分化、遷移、周期阻滯或凋亡等生物學過程。信號分子是細胞的信息載體,種類繁多。信號分子與靶細胞表面受體或胞內受體的特異性結合將受體活化后可以啟動靶細胞內一條或多條信號轉導途徑從而引發細胞代謝或基因表達的改變,進而調控細胞生長與分化[2]。
盡管不同的信號通路具有各自不同的調節機制,但準確的信號傳遞則需要各個通路之間的交叉作用(cross-talk)從而進一步形成復雜的細胞內信號網絡(intracellular network)。隨著細胞內新的信號分子的不斷發現和鑒定,細胞內信號網絡愈加復雜,針對單一信號分子的研究技術和方案已難于滿足對細胞生理活動的全面了解的需要。蛋白質組學技術在細胞信號轉導研究中的應用為我們揭示了細胞內復雜的信號網絡系統,鑒定信號分子復合物,探討蛋白質間相互作用的分子基礎,新伴侶分子的發現,以及已知通路之間的交叉和實現細胞信號轉導的動態變化研究提供了重要手段。
3 蛋白質組學技術在細胞信號轉導研究中的應用
3.1 在蛋白質之間相互作用中的應用
免疫共沉淀(Co-Immunoprecipitation,Co-IP)和酵母雙雜交技術是研究蛋白之間相互作用的基本方法。Co-IP是用抗體將相應特定分子沉淀的同時,與該分子特異性結合的其他分子也會被帶著一起沉淀出來的技術,這種技術常用于驗證蛋白質之間相互特異性結合。酵母雙雜交技術則是在單細胞真核生物酵母在體內利用“誘餌蛋白”(bait)捕獲“獵物蛋白”(prey),二者在細胞內相互作用后形成轉錄激活復合物從而啟動報告基因表達。此外,融合蛋白沉降技術與免疫熒光技術也廣泛適用于蛋白質相互作用的研究。pull-down技術是利用GST對谷朧甘膚偶聯球珠的親和性,從非相互作用蛋白的溶液中純化相互作用蛋白,常采用原核表達純化技術,適用于體外研究蛋白質在溶液中的相互作用。近年單分子操作技術、熒光共振能量轉移(fluorescence resonance energy transfer,FRET)技術也有應用于蛋白質-蛋白質相互作用研究的報道,使得蛋白之間的相互作用檢測實現了直接可測。蛋白質芯片技術目前得以廣泛應用,可將待檢測的蛋白混合樣品與結合到固相基質上的蛋白質相雜交,從而實現對未知蛋白的分離和鑒定。由于蛋白質芯片的高特異性和敏感性,結合質譜、熒光、顯色等方法可以直接或間接地鑒定出與靶蛋白相結合的蛋白質,適用于高通量的蛋白質表達譜分析。
蛋白質復合體在細胞內行使多種功能。鑒定蛋白質復合體的亞基是理解蛋白質復合體功能的基礎。Guerrero等[3]發明了一種體內串聯親和純化交聯蛋白質復合體的定量分析(quantitative analysis of tandem affinity purified in vivo cross-linked(X)protein complexes,QTAX)技術來描述體內蛋白質間相互作用(protein-protein interaction,PPI),并利用這一策略成功繪制了酵母中26S蛋白酶體(細胞內降解蛋白的蛋白質復合體)的相互作用網絡。26S蛋白質復合體中有64對潛在的PPI,其中42對是新發現的相互作用。
較為復雜的研究則包括許多非常規的蛋白質組學手段。例如,酵母是研究G-蛋白偶聯受體(G-protein coupled receptor,GPCR)信號通路的一種模式生物。GPCR研究為理解絲裂原活化蛋白激酶(mitogen activated protein kinase,MAPK)通路包括交配信息素(mating pheromone)的應答提供了基礎。Gruhler等使用13C6-Lysine和13C6-Arginine標記,對賴氨酸和精氨酸營養缺陷型酵母交配信息素誘導的磷酸化改變進行定量蛋白質組學分析,用固定化金屬離子親和層析(immobilized metal ion affinity chromatography,IMAC)富集磷酸化肽段;在離子阱(ion trap)中大量發生中性丟失的離子可被進一步分離、碎裂和分析,這稱為MS/MS/MS(MS3)。準確的母離子質量結合MS3圖譜信息,可以提高幾個數量級的磷酸肽鑒定可信度。該研究以混合線性離子阱-傅立葉變換粒子回旋共振(linear ion trap-Fourier transform ion cyclotron resonance,LTQ-FTICR)質譜儀進行MS/MS和中性丟失導向(neutral lossdirected)的MS3分析,提高對磷酸肽檢測及鑒定的靈敏度和準確性。利用MS/MS掃描自動觸發數據依賴的MS3對中性丟失的母離子進行碎裂,這一中性丟失依賴的MS3運行模式會出現特征性磷酸丟失(-98 Da),通過2次連續的串聯質譜對磷酸肽進行測序。共鑒定了700個磷酸肽,其中139個表達變化在2倍以上,主要是MAPK信號通路成員[4]。
3.2 在分子伴侶研究中的應用
分子伴侶是存在于細胞中,幫助蛋白質正確折疊使之正常行駛功能的小分子蛋白質,對細胞內的信號轉導具有重要意義。近幾年應用蛋白質組學技術在分子伴侶的相關研究中取得了較大的進展(表 1)。
3.3 在信號轉導通路之間的關聯與機制研究中的應用
細胞信號傳導是細胞通過胞外或胞內的刺激后,激活受體而改變細胞內生理機能的過程,這種刺激可以是很小的分子、蛋白、甚至是光子,細胞經過刺激后通過基因表達水平的改變對外界刺激做出回應。通過對調節生長發育起關鍵作用的信號通路分子進行研究發現,應答過程不是一條通路的作用結果,而是不同通路之間交互的調控作用結果[11]。隨著蛋白質組學的發展,對于細胞通路的交叉作用的研究顯得極為重要。通過研究不同通路之間的交叉作用,發現了一些重要通路間的交叉位點(表2)。
4 小結
雖然蛋白質組學技術已經給相關研究帶來了許多指導性的結果,但是相關的技術仍具有很大的發展空間。首先是蛋白質的分析與鑒定方面,將某個蛋白質組樣品中的所有蛋白質分離并檢測仍然是不現實的,目標蛋白質過低的豐度,特殊的蛋白質修飾等都對此技術的發展造成了阻礙。這樣便限制了對未知的細胞信號通路中或已知的通路中未知蛋白質的發現。此外,將一個蛋白質相互作用組與另一個相比較幾乎是不可實現的,兩個相互作用網絡間的比較仍然沒有很好的方法能夠完成,這限制了信號通路間交叉作用的研究。倘若這一技術得到發展,兩個相互作用網絡的差異得以方便地確定,則會大大增進通路間交叉作用的新發現。
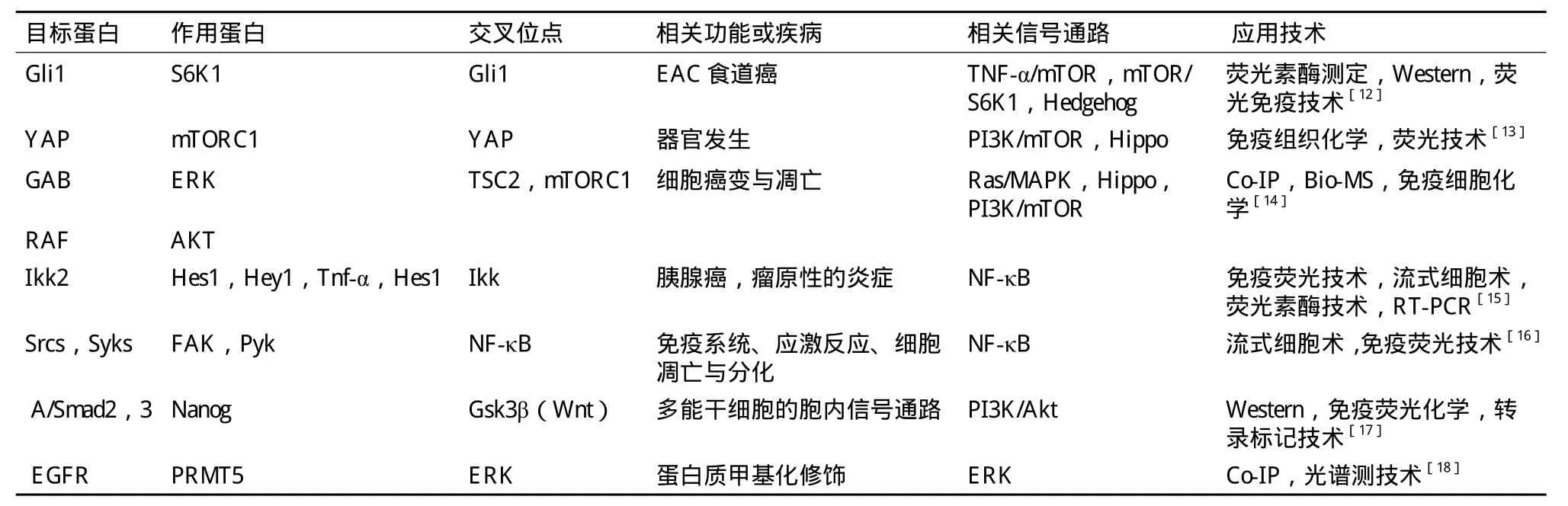
表2 應用蛋白質組學技術發現的重要通路交叉位點及相關功能
未來蛋白質組學在信號轉導研究中可能會更加側重于生命體依賴的復合物、通路以及網絡的形式來協調蛋白質之間的相互作用的研究方向。例如,目前通路間的蛋白質相互作用網絡是一個相對靜態的連線圖,需要將基因敲除及系統擾動等數據加入進來,以期認識信號轉導作用的本質,發現網絡的核心。除此之外,如何在不影響細胞的正常生理條件下的活細胞分析及檢測動態的蛋白間相互作用仍然會是研究的熱點。
[1] Wilkins MR, Sanchez JC, Gooley AA, et al. Progress with proteome projects:why all proteins expressed by a genome should be identified and how to do it[J]. Biotechnol Genet Eng Rev, 1995,13:19-50.
[2] Jayaseelan S, Tenenbaum SA. Neurodevelopmental disorders:Signalling pathways of fragile X syndrome[J]. Nature, 2012, 492:359-360.
[3] Guerrero C, Tagwerker C, Kaiser P, Huang L. An integrated mass spectrometry-based proteomics approach:QTAX to decipher the 26 S proteasome interacting network[J]. Mol Cell Proteomics, 2006, 5:366-378.
[4] Gruhler A, Olsen JV, Mohammed S, et al. Quantitative phosphorproteomics applied to the yeast pheromone signaling pathway[J]. Mol Cell Proteomics, 2005, 4:310-327.
[5] Haupt A, Joberty G, Bantscheff M, et al. Hsp90 inhibition differentially destabilises MAP kinase and TGF-beta signalling components in cancer cells revealed by kinase-targeted chemoproteomics[J].BMC Cancer, 2012, 12:38.
[6] Moulick K, Ahn JH, Zong H, et al. Affinity-based proteomics reveal cancer-specific networks coordinated by Hsp90[J]. Nat Chem Biol, 2011, 7:818-826.
[7] Witt SN. Molecular chaperones, alpha-synuclein, and neurodegeneration[J]. Mol Neurobiol, 2013, 47:552-560.
[8] Kato Y, Kajiwara C, Ishige I, et al. HSP70 and HSP90 differentially regulate translocation of extracellular antigen to the cytosol for crosspresentation[J]. Autoimmune Dis, 2012:745962.
[9] Simet SM, Pavlik JA, Sisson JH. Proteomic analysis of bovine axonemes exposed to acute alcohol:role of endothelial nitric oxide synthase and heat shock protein 90 in cilia stimulation[J].Alcohol Clin Exp Res, 2013, 37(4):609-615.
[10] Morgan ED, Eunice HL, Elizabeth AS, et al. Operational plasticity enables Hsp104 to disaggregate diverse amyloid and nonamyloid clients[J]. Cell, 2012, 151:778-793.
[11] Vert G, Chory J. Crosstalk in cellular signaling:background noise or the real thing?[J]. Dev Cell, 2011, 6:985-991.
[12] Wang Y, Ding QQ, Yen CJ, et al. The crosstalk of mTOR/S6K1 and Hedgehog pathways[J]. Cancer Cell, 2011, 21 :374-387.
[13] Tumaneng K, Schlegelmilch K, Russell RC, et al. YAP mediates crosstalk between the Hippo and PI(3)K-TOR pathways by suppressing PTEN via miR-29[J]. Nature Cell Biology, 2012,14:1322-1329.
[14] Mendoza MC, Er EE, Blenis J. The Ras-ERK and PI3K-mTOR pathways:cross-talk and compensation[J]. Trends Biochem Sci, 2011, 6:320-328.
[15] Maniati E, Bossard M, Cook N, et al. Crosstalk between the canonical NF-κB and Notch signaling pathways inhibits Pparγ expression and promotes pancreatic cancer progression in mice[J]. J Clin Invest, 2011, 121:4685-4699.
[16] Oeckinghaus A, Hayden MS, Ghosh S. Crosstalk in NF-κB signaling pathways[J]. Nature Immunology, 2011, 12 :695-708.
[17] Singh AM, Reynolds D, Cliff T, et al. Signaling network crosstalk in human pluripotent cells:a Smad2/3-regulated switch that controls the balance between self-renewal and differentiation[J]. Cell Stem Cell, 2012, 3:312-326.
[18] Hsu JM, Chen CT, Chou CK, et al. Crosstalk between Arg 1175 methylation and Tyr 1173 phosphorylation negatively modulates EGFR-mediated ERK activation[J]. Nature Cell Biology, 2012,13:174-181.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中國生殖健康(2019年3期)2019-02-01 06:12:26
汽車工程學報(2017年2期)2017-07-05 08:13:02
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
