榮昌豬C-Fos原癌基因克隆與序列分析
2013-11-22 05:21:56鄭小波
中國(guó)獸醫(yī)雜志 2013年5期
盧 梅,孫 波,李 建,肖 榕,李 靖,鄭小波
(西南大學(xué)榮昌校區(qū)動(dòng)物醫(yī)學(xué)系,重慶 榮昌402460)
原癌基因是細(xì)胞內(nèi)與細(xì)胞增殖相關(guān)的基因,是維持機(jī)體正常生命活動(dòng)所必須的,在進(jìn)化上高度保守。原癌基因是細(xì)胞的正常基因,其表達(dá)產(chǎn)物對(duì)細(xì)胞的生理功能極其重要,當(dāng)原癌基因的結(jié)構(gòu)或調(diào)控區(qū)發(fā)生變異,基因產(chǎn)物增多或活性增強(qiáng)時(shí),使細(xì)胞過(guò)度增殖,從而導(dǎo)致細(xì)胞癌變形成腫瘤。原癌基因種類很多,涉及到細(xì)胞信號(hào)傳遞的各個(gè)層次[1]。C-Fos是一種原癌基因,屬于快速早期基因。其編碼的Fos蛋白是真核細(xì)胞內(nèi)調(diào)控因子,在信號(hào)傳導(dǎo)過(guò)程中起重要作用。大量研究發(fā)現(xiàn),C-Fos原癌基因及其蛋白不僅參與細(xì)胞的正常生長(zhǎng)、分化過(guò)程,而且參與細(xì)胞內(nèi)信息傳遞過(guò)程和細(xì)胞的能量代謝過(guò)程,在生命活動(dòng)中起著極為基本而重要的作用[2]。
近年來(lái),國(guó)內(nèi)外對(duì)人、野豬、鼠等動(dòng)物C-Fos基因及相關(guān)研究均有報(bào)道,并通過(guò)大量試驗(yàn)證明,在精子發(fā)生過(guò)程中,C-Fos在生殖細(xì)胞內(nèi)呈現(xiàn)出階段性和特異性表達(dá),在生精細(xì)胞發(fā)育過(guò)程中起著重要調(diào)控作用[3]。榮昌豬C-Fos的研究少,本試驗(yàn)對(duì)其進(jìn)行了克隆與序列分析,為進(jìn)一步研究榮昌豬C-Fos基因的結(jié)構(gòu)和功能提供理論依據(jù),同時(shí)也為榮昌豬其他功能基因的研究提供可靠的分子內(nèi)標(biāo)。
1 材料與方法
1.1 材料 本試驗(yàn)所需的睪丸由榮昌畜牧局養(yǎng)殖場(chǎng)提供。EASYspin Plus組織/細(xì)胞RNA試劑盒、反轉(zhuǎn)錄試劑盒、PCR試劑盒、DL-2 000Marker、割膠回收試劑盒、質(zhì)粒抽提試劑盒、E.coliDH5α、pMD18-T載體連接試劑盒等,購(gòu)自寶生物工程(大連)有限公司。
1.2 方法
1.2.1 引物的設(shè)計(jì)與合成 根據(jù)GenBank上登錄的 野 豬 C-Fos序 列 (AJ132510.1),使 用 Primer Primer5軟件設(shè)計(jì)2對(duì)分段特異性引物P1、P2和P1′、P2′,這兩對(duì)引物擴(kuò)增的目的基因片段 C-Fos-1和C-Fos-2長(zhǎng)度分別為827bp和718bp,引物序列如下:
① P1:5′-ATGATGTTCTCCGGCTTCAAC-3′
P2:5′-CTGGGAACAGGAAGTCATCAAA-3′
② P1′:5′-TCCGAAGGGAAAGGAATAAGAT-3′
P2′:5′-TTTTTTTCACAGGGCCAGC-3′
1.2.2 總RNA的提取 組織總RNA的抽提方法按試劑盒說(shuō)明書操作。
1.2.3 cDNA的獲取及RT-PCR 按反轉(zhuǎn)錄試劑盒說(shuō)明操作,以榮昌豬睪丸總RNA為模板反轉(zhuǎn)錄為第一鏈cDNA,利用特異性引物進(jìn)行PCR擴(kuò)增榮昌豬的C-Fos原癌基因。將PCR產(chǎn)物進(jìn)行1%瓊脂糖凝膠電泳鑒定。
1.2.4 純化回收、載體構(gòu)建及陽(yáng)性克隆篩選 按DNA膠回收試劑盒、質(zhì)粒抽提試劑盒、pMD18-T載體連接試劑盒及其說(shuō)明書操作。最后以1%瓊脂糖凝膠電泳陽(yáng)性鑒定。
1.2.5 序列測(cè)定及分析 將PCR檢測(cè)為陽(yáng)性的菌落擴(kuò)大培養(yǎng),保藏,并送TaKaRa公司測(cè)序,拼接。采用DNAman進(jìn)行序列分析后,使用 MEGA5.03軟件按NJ法構(gòu)建系統(tǒng)進(jìn)化樹,Bootstrap值設(shè)為1 000。
2 結(jié)果
2.1 總RNA提取及鑒定 按照EASYspin Plus組織/細(xì)胞RNA快速提取試劑盒說(shuō)明書操作提取榮昌豬睪丸組織總RNA。得到OD260/OD180為2.1,且電泳可見清晰的28s和18s條帶,前者亮度約為后者兩倍,說(shuō)明提取的RNA比較完整且降解較少,符合用于反轉(zhuǎn)錄合成cDNA要求。結(jié)果如圖1。
2.2 C-Fos的PCR擴(kuò)增 用特異性引物以榮昌豬的總RNA為模板進(jìn)行PCR擴(kuò)增,在750bp上下各擴(kuò)增出明顯的單一條帶,它與預(yù)期估計(jì)值827bp、718bp較為接近(圖2)。
2.3 PCR鑒定陽(yáng)性克隆 將純化的C-Fos PCR產(chǎn)物與pMD18-T Vector連接,CaCl2法轉(zhuǎn)化E.coliDH5α,LB/Amp固體培養(yǎng)基37℃培養(yǎng)12~14h,挑取單個(gè)菌落于LB/Amp液體培養(yǎng)基再培養(yǎng)6~7h,然后將菌液經(jīng)PCR擴(kuò)增鑒定陽(yáng)性克隆,結(jié)果為陽(yáng)性(圖3)。
2.4 C-Fos堿基測(cè)序及分析 將經(jīng)過(guò)PCR鑒定陽(yáng)性的菌液送至TaKaRa公司進(jìn)行雙向測(cè)序,根據(jù)各序列的重疊區(qū)域進(jìn)行拼接,得出全長(zhǎng)為1 143bp,最終得到榮昌豬C-Fos ORF序列并上傳至GenBank,獲得序列號(hào)JX861095。將榮昌豬C-Fos序列與野豬(AJ132510.1),馬 (XM _001491972.2),牛(AY322482.1),綿羊(NM_001166182.1),家貓(DQ288168.1),大熊貓(XM_002914704.1),人(NM_004469.4),黑猩猩(AB219119.1),獼猴(AB219121.1),大鼠(DQ089699.1),小鼠(NM_010234.2),家兔(XM_002714687.1),等 C-Fos用DNAman進(jìn)行核酸同源性比對(duì),結(jié)果顯示,同源性分別為99.48%、95.20%、95.20%、94.14%、93.7%、94.14%、93.0%、93.10%、93.10%、88.58%、82.48%、88.58%。用MEGA 5.03按NJ法構(gòu)建系統(tǒng)進(jìn)化樹(圖4)。
3 討論
研究發(fā)現(xiàn)C-Fos原癌基因表達(dá)有組織、細(xì)胞、階段的特異性。在胚胎早期僅在胚胎外組織如羊膜和胎盤中表達(dá)。胚胎后期則出現(xiàn)在發(fā)育中的中樞神經(jīng)系統(tǒng)、胚胎骨、軟骨和牙齒生長(zhǎng)區(qū)[4]。在成體器官中,C-Fos表達(dá)于巨噬細(xì)胞和粒細(xì)胞,另外還在生殖細(xì)胞中表達(dá)。但在其他大多數(shù)的細(xì)胞中,它的表達(dá)水平相對(duì)較低,需要通過(guò)刺激才能維持高表達(dá)[5,16]神經(jīng)系統(tǒng)科學(xué)家認(rèn)為,在神經(jīng)元產(chǎn)生動(dòng)作電位的時(shí)候經(jīng)常表達(dá)[6]。C-Fos已經(jīng)被證明與許多物質(zhì)如酪蛋白激酶2[7],α1、B細(xì)胞淋巴瘤蛋白3抗體[8]、DNA 損傷誘導(dǎo)轉(zhuǎn)錄因子 3[9]、輔因子 BRCA1[10]、TATA 結(jié)合蛋白[11]、核受體輔活化因子1,2[12]、相關(guān)轉(zhuǎn)錄因子1,2[13]、c-jun[14]和 SMAD3[15]等相關(guān)。目前國(guó)內(nèi)外對(duì)C-Fos的研究相對(duì)較少,對(duì)豬的研究更少。GenBank上登陸的C-Fos序列來(lái)自野豬,而榮昌豬作為中國(guó)四大地方豬種之一,卻缺乏此方面的研究。本試驗(yàn)根據(jù)GenBank上登錄的野豬CFos序列,用Primer Primer5軟件進(jìn)行分段設(shè)計(jì)引物。擴(kuò)增出特異性片段后,經(jīng)雙向測(cè)序,獲得兩段序列。根據(jù)各序列的重疊區(qū)域進(jìn)行拼接,得出全長(zhǎng)為1 143bp,編碼381個(gè)氨基酸,其起始密碼子為ATG終止密碼子為TGA。
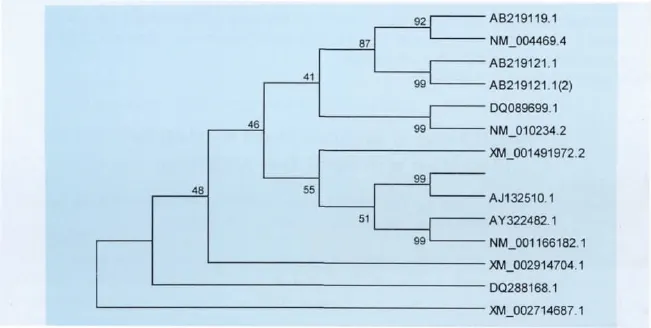
圖4 不同種C-Fos CDS序列構(gòu)建NJ法系統(tǒng)進(jìn)化樹
將榮昌豬C-Fos與野豬、馬、牛等進(jìn)行分析比對(duì),發(fā)現(xiàn)在這些種系中的Fos蛋白序列統(tǒng)一性為69.40%,而且在這些種系中的Fos均存在一個(gè)由88個(gè)完全相同的氨基酸順序組成的區(qū)域。這個(gè)區(qū)域包括一個(gè)能與DNA結(jié)合的基本區(qū)和LZ結(jié)構(gòu)。榮昌豬C-Fos的CDS序列與其他動(dòng)物的CDS進(jìn)行同源性分析比較,結(jié)果顯示,其進(jìn)化關(guān)系與基因水平一致,榮昌豬與野豬的同源性最高,與其他動(dòng)物則較野豬的同源性相對(duì)較低。用 MEGA 5.03軟件按NJ法構(gòu)建系統(tǒng)進(jìn)化樹,結(jié)果顯示,C-Fos在種間、種內(nèi)的距離與寬度和重疊排列上都有所不同,相同的物種聚為一枝,且自展自持率均大于90%,大熊貓、貓、兔獨(dú)立成枝與榮昌豬的親緣關(guān)系更遠(yuǎn)。
[1] 冀群升,章靜波.C-Fos原癌基因的研究進(jìn)展[J].國(guó)外醫(yī)學(xué)分子生物學(xué)分冊(cè),1994,16(4):152-156.
[2] 于芳,張安民.運(yùn)動(dòng)與C-Fos原癌基因的研究進(jìn)展[J].中國(guó)醫(yī)學(xué)研究雜志,2006,6:635-637.
[3] 雷忻,廉振民,李游游,等.C-Fos與生殖細(xì)胞發(fā)育[J].延安大學(xué)學(xué)報(bào)(自然科學(xué)版),2007,26(1):72-74.
[4] 韓濟(jì)生.神經(jīng)科學(xué)綱要[M].北京:北京醫(yī)科大學(xué),中國(guó)協(xié)和醫(yī)科大學(xué)聯(lián)合出版社,1993:535-552.
[5] Van Elzakker M,F(xiàn)evurly R D,Breindel T,etal.Environmental novelty is associated with a selective increase in Fos expression in the output elements of the hippocampal formation and the perirhinal cortex[J].Learn Mem,2008,15(12):899-908.
[6] Day H E,Kryskow E M,Nyhuis T J,etal.Conditioned Fear Inhibits C-Fos mRNA Expression in the Central Extended A-mygdala[J].Brain Res,2008,1229:137-146.
[7] Yamaguchi Y,Wada T,Suzuki F,etal.Casein kinaseⅡinteracts with the bZIP domains of several transcription factors[J].Nucleic Acids Res,ENGLAND,1998,26(16):3854-3861.
[8] Na S Y,Choi J E,Kim H J,etal.Bcl3,an IkappaB protein,stimulates activating protein-1transactivation and cellular proliferation[J].Biol Chem UNITED STATES,1999,274(40):28491-28496.
[9] Ubeda M,Vallejo M,Habener J F.CHOP Enhancement of Gene Transcription by Interactions with Jun/Fos AP-1Complex Proteins[J].Mol Cell Biol UNITED STATES,1999,19(11):7589-7599.
[10]Zhong Hongjun,Zhu Jianhua,Zhang Hao,etal.COBRA1inhibits AP-1transcriptional activity in transfected cells[J].Biochem Biophys Res Commun.United States,2004,325(2):568-573.
[11]Metz R,Bannister A J,Sutherland J A,Hagemeier C,etal.C-Fos-induced activation of a TATA-box-containing promoter involves direct contact with TATA-box-binding protein[J].Mol Cell Biol UNITED STATES,1994,14(9):6021-6029.
[12]Lee,S K,Na S Y,Jung S Y,etal.Activating protein-1,nuclear factor-kappaB,and serum response factor as novel target molecules of the cancer-amplified transcription coactivator ASC-2[J].Mol Endocrinol UNITED STATES,2000,14(6):915-925.
[13]Lee S K,Kim J H,Lee Y C,etal.Silencing mediator of retinoic acid and thyroid hormone receptors,as a novel transcriptional corepressor molecule of activating protein-1,nuclear fac-tor-kappaB,and serum response factor[J].Biol Chem UNITED STATES,2000,275(17):12470-12474.
[14]Pognonec P,BouLukos K E,Aperlo C,etal.Cross-family interaction between the bHLHZip USF and bZip Fra1proteins results in down-regulation of AP1activity[J].Oncogene ENGLAND,1997,14(17):2091-2098.
[15]Zhang Y,F(xiàn)eng X H,Derynck R.Smad3and Smad4cooperate with c-Jun/C-Fos to mediate TGF-beta-induced transcription[J].Nature ENGLAND,1997,394(6696):909-913.
[16]王雪.神經(jīng)細(xì)胞的生理和病理過(guò)程與原癌基因C-Fos的關(guān)系[J].Foregin Medical Science of Pathophysiology and Clinical Medicine,1998,18:358-361.
