脫落酸不同處理時間對丹參毛狀根有效成分積累的影響
2013-12-23 05:27:06盛東峰朱自學
天然產物研究與開發 2013年6期
盛東峰 ,朱自學
周口師范學院生命科學系,周口466000
丹參為唇形科植物丹參(Salvia miltiorrhiza Bge.)的干燥根及根莖,主要活性成分丹參酮類(包括丹參酮Ⅰ、ⅡA、隱丹參酮、二氫丹參酮Ⅰ等),是一類二萜醌化合物,常被用于微循環系統疾病的治療。有研究表明,丹參酮類主要通過細胞質內甲羥戊酸途徑(mevalonate,MVA)[1]和質體內非甲羥戊酸途徑(2-C-methyl-d-erythritol-4-phosphate,MEP)[2]兩條途徑合成。
毛狀根是發根農桿菌Ri 質粒的一段T-DNA 導入植物基因組中并表達的結果。丹參毛狀根由于其培養過程中無需外源植物激素,生長速度快,合成次生代謝物的能力強而穩定。常被用于丹參酮生產及丹參酮合成機理的研究。干旱是影響作物生長發育的重要逆境因子,大量研究表明干旱能降低作物的最終產量。但也有資料顯示干旱能誘導植物次生代謝產物(如萜類)成分的積累[3]。然而迄今為止,人們對干旱脅迫誘導丹參酮積累的機制尚不清楚。脫落酸(ABA)是一種植物激素,因其能抑制生長促使葉子脫落而得名,是干旱脅迫響應的重要信號物質。有研究顯示ABA 能夠誘導植物非甲羥戊酸途徑相關代謝產物的合成[4]。本研究以丹參毛狀根為試材,對ABA 不同處理時間條件下丹參酮類的積累規律進行了研究,以期揭示干旱脅迫對丹參酮類合成的調控機理。
1 材料與方法
1.1 實驗材料
發根農桿菌菌株15834 購自中國林業科學院;無菌丹參苗葉。
1.2 試劑與儀器
SY-360 超聲波提取儀(上海寧商超聲儀器有限公司);360EP 電子天平(上海精科儀器有限公司);Waters 1525 二元高效液相色譜儀(美國Waters 公司);RO-MB-10D 高純水機(杭州永潔達膜分離設備廠);smartcycler 熒光定量PCR(美國Cepheid 公司)。
脫酸酸(高純級,美國Sigma 公司);磷甘霉素(美國Santa Cruz biotechnology 公司);乙腈(分析純,山東齊魯石化);鎢酸鈉(江蘇姜堰貝斯特鉬制品有限公司);洛伐他汀(美國Sigma 公司);丹參酮Ⅰ、隱丹參酮、二氫丹參酮Ⅰ和丹參酮ⅡA 等標準品(中國食品藥品鑒定所);cDNA 合成試劑盒(美國Promega 公司);RNAisoTM Plus 試劑盒(大連寶生物工程有限公司);Brilliant III Ultra-Fast QPCR Master Mix 試劑盒(美國Agilent 公司)。
1.3 試驗方法
1.3.1 毛狀根培養及處理
在無硝酸銨的MS(Murashige and Skoog)培養基中,用發根農桿菌ATCC15834 誘導丹參無菌苗葉生產毛狀根。然后取生長良好的丹參毛狀根0.45 g置于含50 mL MS 液體培養基的三角瓶內,于25 ℃培養箱中黑暗下懸浮繼代培養。
繼代培養20 d 后,用濃度為210 μM 的ABA 處理,并分別于處理后1、3、6、9 d 收獲毛狀根,以研究ABA 處理不同時間對丹參毛狀根生長和丹參酮類積累的影響。在ABA 處理毛狀根第6 d 時,分別使用145 μM 磷甘霉素(FOS)、15 μM 洛伐他汀(MEV)和100 μM 的鎢酸鈉(TUN)處理毛狀根,以研究抑制劑對丹參酮類積累的影響。同時用等試驗組體積的無菌蒸餾水處理的毛狀根作對照。每個處理組重復3 次,并分別于處理結束時立即收獲毛狀根,45 ℃烘干至恒重,稱重,保存。
1.3.2 丹參酮的提取
取干燥至衡重的丹參毛狀根適量研缽磨碎,過0.45 mm 篩,混勻后精密稱取0.1000 g 細粉,加入2 mL 甲醇-水(7∶3)提取液,精密稱重,超聲處理45 min,10000 rpm 高速離心15 min,取上清液過0.45 μm 濾膜,即得。
1.3.3 丹參酮含量的HPLC 定量分析[5]采用RP-HPLC 色譜方法。色譜條件為:色譜柱Waters SunFire C18(250 mm ×4.6 mm,5 μm);柱溫30 ℃,流速1.0 mL/min,上樣體積20 μL,檢測波長270 nm,流動相乙腈和水,線性梯度洗脫:0~5 min,40%乙腈;5~20 min;60%乙腈;20~23 min,60%乙腈;23~25 min,80%乙腈;25 min,100%乙腈。
1.3.4 RNA 分離、cDNA 合成及熒光RT-PCR 分析把處理24 h 的毛狀根置于液氮中研磨成粉,采用購至大連寶生物工程有限公司RNAisoTMPlus 試劑盒提取總RNA。并以提取的RNA(500ng)為模板,采用美國Promega 公司生產的cDNA 試劑盒合成第一鏈cDNA。熒光RT-PCR 實時定量分析采用Brilliant III Ultra-Fast QPCR Master Mix 試劑盒(美國)。PCR 反應體積為25 μL,PCR 反應程序為:95℃~10 min,1 循環;95 ℃~30 s,60 ℃~1 min,72℃~30 s,40 循環。實驗共設置3 個重復,內參基因為β-actin,3-羥基-3-甲基戊二酰輔酶A 還原酶(HMGR)和1-去氧木糖-5-磷酸還原酶(DXR)基因引物Oligo6 軟件設計(表1)。
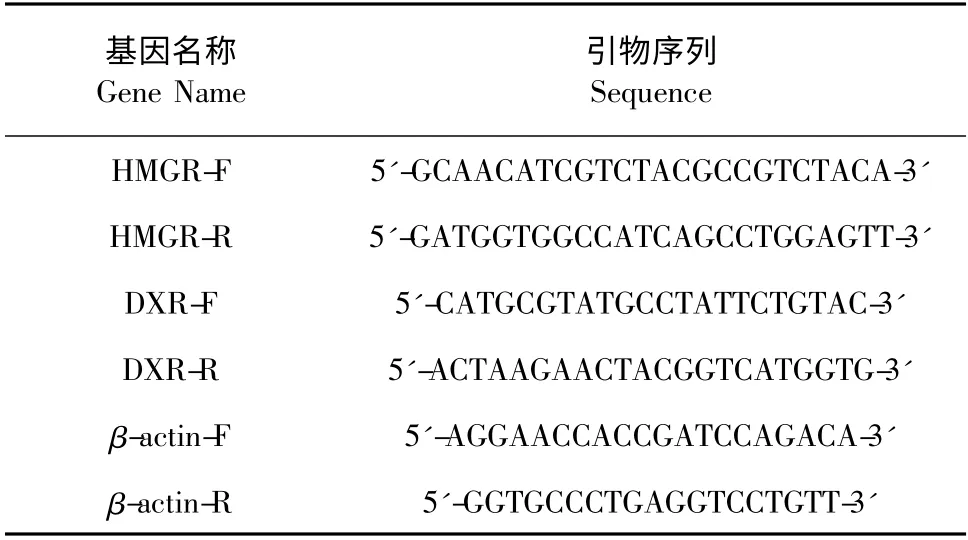
表1 RT-PCR 分析引物設計Table 1 The primers of genes used in RT-PCR
2 結果與討論
2.1 不同ABA 處理時間對丹參毛狀根生長的影響
不同ABA 處理時間對丹參毛狀根的生長的影響見表2,由表2 可知,隨著ABA 處理時間延長,毛狀根的干重逐漸減少。和對照組相比,處理1、3、6、9 d 后,收獲的毛狀根分別減少為94.9%、88.5%、83.3%和80.8%。這說明,隨著ABA 處理時間的增加,丹參毛狀根生長受到抑制逐漸減弱,并在處理6 d 左右變的不顯著。ABA 和三種丹參酮類合成抑制劑均對丹參毛狀根的生長產生抑制效果,和FOS與MEV 相比,TUN +ABA 的抑制作為最弱,FOS +ABA 的抑制作用強于TUN + ABA,但弱于MEV +ABA。這說明MEV 能增強ABA 對丹參毛狀根生長的抑制作用。

表2 不同ABA 處理時間對丹參毛狀根生長的影響Table 2 Effects of ABAtreatment time on growth of S.miltiorrhiza hairy roots
2.2 不同ABA 處理時間對丹參酮類積累的影響
不同ABA 處理時間對丹參毛狀根中丹參酮Ⅰ、隱丹參酮、二氫丹參酮Ⅰ和丹參酮ⅡA 產量和含量的影響見表3 和圖1。從表3 和圖1 可以看出,丹參毛狀根在培養過程中經ABA 處理不同時間后,四種丹參酮的產量和含量均在ABA 處理6 d 左右達到穩定。其中丹參酮Ⅰ、隱丹參酮、二氫丹參酮Ⅰ和丹參酮Ⅱ的產量分別提高為空白對照組的4. 65、7.80、18.33 和2.11 倍。這表明ABA 是促進丹參酮積累的有效誘導因子。
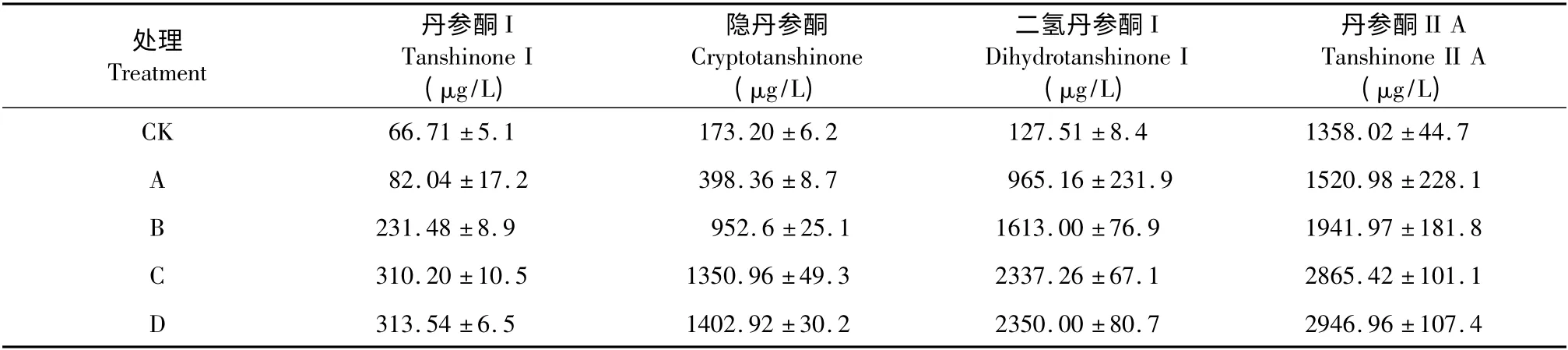
表3 不同ABA 處理時間對毛狀根四種丹參酮產量的影響Table 3 Yields of four tanshinone components in S.miltiorrhiza hairy roots with different ABA treatments time
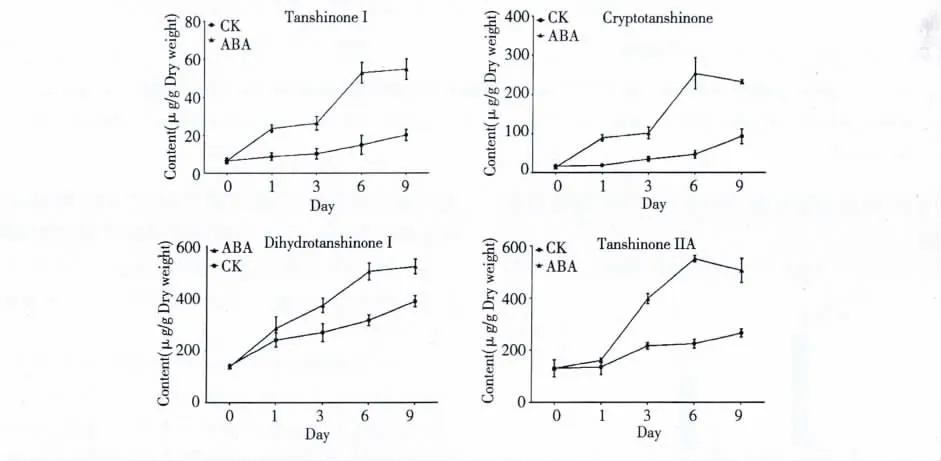
圖1 不同ABA 處理時間對丹參酮含量的影響(CK 為空白對照)Fig.1 Effects of ABA treatment time on tanshinone production in S.miltiorrhiza hairy roots (CK was the blank control)
2.3 抑制劑和ABA 聯合處理對丹參酮類積累的影響
磷甘霉素是丹參酮非甲羥戊酸途徑合成過程中重要酶1-去氧木糖-5-磷酸還原酶(DXR)的專一性抑制劑;洛伐他汀是甲羥戊酸途徑合成途徑限速酶3-羥基-3-甲基戊二酰輔酶A 還原酶(HMGR)的專一性抑制劑;鎢酸鈉是植物內源脫落酸合成的抑制劑之一,能夠抑制干旱脅迫條件下植物中脫落酸的積累。三種抑制劑均可不同程度抑制MEP 和MVA途徑下游產物的積累和內源ABA 的合成。
本研究結果表明(見圖2),FOS 與ABA 聯合處理可顯著抑制四種丹參酮含量的增加。其中,丹參酮Ⅰ、丹參酮ⅡA 和二氫丹參酮Ⅰ的含量降至接近對照組水平,隱丹參酮的含量降至對照組的37%;MEV 與ABA 聯合處理對丹參酮Ⅰ和隱丹參酮的含量變化無顯著影響,但能顯著抑制丹參酮Ⅰ和丹參酮ⅡA 的積累,其抑制效果弱于FOS。這意味著ABA 可能主要通過誘導MEP 途徑促進丹參酮的累積,但從結果也可以看出,MVA 途徑對丹參酮類的積累可能也起著重要作用。
TUN 與ABA 聯合處理對毛狀根四種丹參酮含量均有顯著的抑制作用,其中丹參酮Ⅰ含量降至接近對照組水平,丹參酮ⅡA 和二氫丹參酮Ⅰ含量低于對照組,隱丹參酮的含量則略高于對照組。本研究結果說明,外源ABA 對丹參酮類的誘導作用很可能與其內源ABA 有關。

圖2 抑制劑FOS、MEV 和TUN 對ABA 處理條件下丹參酮積累的影響(CK 為空白對照)Fig.2 Effects of FOS (145μM),MEV (15μM)and TUN (100μM)on ABA (210 μM)-induced tanshinone production (CK was the control)
2.4 ABA 對丹參毛狀根HMGR 和DXR 基因表達的影響
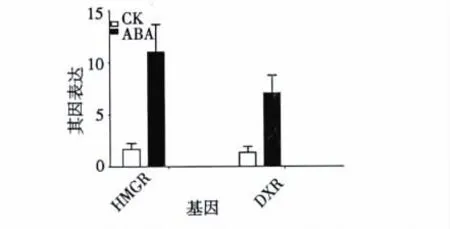
圖3 ABA 對丹參毛狀根HMGR 和DXR 基因表達的影響Fig.3 Effects of ABA (210 μM)on expression of HMGR and DXR.
圖3 顯示了ABA 對丹參毛狀根HMGR 和DXR基因表達的影響,從圖3 可以看出,ABA 夠能顯著誘導丹參毛狀根MVA 中限速酶HMGR 基因和MEP中重要酶DXR 基因的表達,但筆者認為,這種誘導作用可能更多的依賴丹參毛狀根內源ABA 的合成。
2.5 討論
ABA 是植物重要的脅迫響應信號,植物會在脅迫響應敏感部位合成大量ABA[6]。有研究顯示,ABA 能夠誘導植物萜類成分的積累[7],但也有研究顯示,ABA 會抑制植物萜類成分的積累[8]。本研究結果表明,隨著ABA 處理時間的延長,丹參毛狀根的生長被抑制,但其丹參酮類的含量和終產量增加,這說明,ABA 能夠促進毛狀根丹參酮類的積累。
FOS 和MEV 作為植物萜類成分重要合成途徑(MVA 和MEP 途徑)中關鍵酶的抑制劑,已經被廣泛應用于植物萜類合成的調控研究[9]。盡管植物萜類主要通過MEP 途徑合成,但兩條途徑之間的相互作用對萜類合成的影響也不能忽視[10]。本試驗結果表明,FOS 和MEV 均能增大ABA 對丹參毛狀根中丹參酮類積累的抑制作用。筆者認為,ABA 能促進毛狀根中丹參酮類的積累,可能與其能誘導DXR 和HMGR 基因表達,激活毛狀根MVA 和MEP途徑,特別是MEP 途徑有關。TUN 與ABA 聯合處理毛狀根的相關數據也表明,ABA 能夠抑制毛狀根內源ABA 的合成,進而影響外源ABA 對丹參毛狀根的處理效果。
3 結論
本試驗通過研究不同ABA 處理時間對丹參毛狀根的產量、丹參酮類含量及其積累規律的影響,以及ABA 和相關萜類合成途徑抑制劑對丹參毛狀根的聯合抑制效果,得到的結論是:①ABA 對丹參毛狀根的生長抑制作用在處理6 d 左右減弱。ABA 能抑制毛狀根的生長,但能誘導丹參酮類的積累和其主要合成途徑中限速酶HMGR 與DXR 基因的表達;②磷甘霉素(FOS)幾乎能完全抑制ABA 對四種丹參酮的積累作用,但洛伐他汀(MEV)僅能部分抑制丹參酮的積累;TUN 則能顯著抑制內源ABA 的生成;③ABA 對兩種丹參酮合成途徑MVA 和MEP 都有激活作用,但可能主要通過激活MEP 途徑誘導丹參酮的積累。
1 Lichtenthaler HK. Non-mevalonate isoprenoid biosynthesis:enzymes,genes and inhibitors.Biochem Soc Trans,2000,28:785-789.
2 Rohmer M.The discovery of a mevalonate-independent pathway for isoprenoid biosynthesis in bacteria,algae and higher plants.Nat Prod Rep,1999,16:565-574.
3 Wang JW,Wu JY.Tanshinone biosynthesis in Salvia miltiorrhiza and production in plant tissue cultures. Appl Microbiol Biotechnol,2010,88:437-449.
4 Mansouri H,Asrar Z,Szopa J.Effects of ABA on primary terpenoids and Δ9-tetrahydrocannabinol in Cannabis sativa L.at flowering stage.Plant Growth Regul,2009,58:269-277.
5 Gu J (顧娟),Luo HY(羅海燕),An LY(安連英),et al.The study on the extraction of the salvianolic acid by water.Nat Prod Res Dev(天然產物研究與開發),2005,17:781-783.
6 Zhang J,Jia W,Yang J,et al. Role of ABA in integrating plant responses to drought. Field Crops Res,2006,25:111-119.
7 Mansouri H,Asrar Z.Effects of abscisic acid on content and biosynthesis of terpenoids in Cannabis sativa at vegetative stage.Biologia Plantarum,2012,55:153-156.
8 Pan QF,Chen Y,Wang Q,et al.Effect of plant growth regulators on the biosynthesis of vinblastine,vindoline and catharanthine in Catharanthus roseus.Plant Growth Regul,2010a,60:133-141.
9 Skorupinska-Tudek K,Poznanski J,Wojcik J,et al.Contribution of the mevalonate and methylerythritol phosphate pathways to the biosynthesis of dolichols in plants.J Biol Chem,2008,283:21024-21035.
10 Ge XC,Wu JY.Tanshinone production and isoprenoid pathways in Salvia miltiorrhiza hairy roots induced by Ag+and yeast elicitor.Plant Sci,2005b,168:487-491.
猜你喜歡
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
中國衛生(2016年3期)2016-11-12 13:23:26
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
