多不飽和脂肪酸對(duì)大鼠腸道菌群及脂肪代謝相關(guān)基因的影響
2014-01-21 02:32:28喬立君馬文慧蘆庭秀蓋曉英葛銀林
食品科學(xué) 2014年17期
關(guān)鍵詞:質(zhì)量
喬立君,鄭 征,馬文慧,蘆庭秀,蓋曉英,葛銀林
多不飽和脂肪酸對(duì)大鼠腸道菌群及脂肪代謝相關(guān)基因的影響
喬立君,鄭 征,馬文慧,蘆庭秀,蓋曉英,葛銀林*
(青島大學(xué)醫(yī)學(xué)院,生物化學(xué)與分子生物學(xué)教研室,山東 青島 266021)
目的:研究多不飽和脂肪酸(polyunsaturated fat acids,PUFA)飲食對(duì)大鼠腸道菌群及相關(guān)脂肪因子的影響,探討其作用機(jī)理。方法:將30 只5 周齡健康大鼠分為正常對(duì)照組、n-6 PUFA組和n-3 PUFA組,自由攝食飲水8 周,每周記錄大鼠體質(zhì)量,實(shí)驗(yàn)結(jié)束時(shí)取大鼠盲腸糞便和肝臟,用實(shí)時(shí)熒光定量聚合酶鏈?zhǔn)椒磻?yīng)檢測(cè)雙歧桿菌(Bifi dobacterium,Bif)、乳酸桿菌(Lactobacillus,Lac)、腸道細(xì)菌Akkermansia muciniphila(Akk)和脂肪因子脂肪酸合成酶(fatty acid synthetase,F(xiàn)AS)、禁食誘導(dǎo)脂肪因子(fasting-induced adipose factor,F(xiàn)IAF)的水平;并將大鼠盲腸石蠟包埋、切片、蘇木精-伊紅染色測(cè)量其黏膜厚度。結(jié)果:n-3 PUFA組與正常對(duì)照組相比,肥胖程度和FAS水平明顯降低(P<0.05),Bif、Lac、Akk和FIAF水平明顯升高(P<0.05),腸道黏膜厚度明顯增加;n-6 PUFA組大鼠的各項(xiàng)指標(biāo)和正常對(duì)照組相比均無(wú)統(tǒng)計(jì)學(xué)差異。結(jié)論:n-3 PUFA飲食與n-6 PUFA飲食相比,可改變腸道菌群,抑制肥胖的發(fā)生和發(fā)展。
n-3多不飽和脂肪酸;肥胖;Akkermansia muciniphila;腸道黏膜
肥胖是指一定程度的明顯超重與脂肪層過(guò)厚,是體內(nèi)脂肪,尤其是甘油三酯積聚過(guò)多而導(dǎo)致的一種狀態(tài)。由于食物攝入過(guò)多或機(jī)體代謝的改變而導(dǎo)致體內(nèi)脂肪積聚過(guò)多,造成體質(zhì)量過(guò)度增長(zhǎng)并引起人體病理、生理改變或潛伏。歷年來(lái),肥胖一直是人們急需解決的問(wèn)題。有不少研究證明,魚(yú)油中的n-3多不飽和脂肪酸(polyunsaturated fat acids,PUFA)可對(duì)脂肪因子產(chǎn)生影響,有一定的減肥作用。n-3 PUFA,尤其是二十碳五烯酸(eicosapentaenoic acid,EPA)和二十二碳六烯酸(docosahexaenoic acid,DHA)具有很強(qiáng)的生理活性,對(duì)人和動(dòng)物的多種疾病,如癌癥等具有特殊的預(yù)防和治療效果,是一種重要的免疫調(diào)理性營(yíng)養(yǎng)素。n-3 PUFA作為 一種特殊的腸內(nèi)營(yíng)養(yǎng)素,可以減輕腸道黏膜損傷,促進(jìn)腸道炎癥愈合,對(duì)腸道具有保護(hù)作用,但關(guān)于其對(duì)腸道微生態(tài)影響方面的文獻(xiàn)報(bào)道并不多。本實(shí)驗(yàn)通過(guò)實(shí)時(shí)熒光定量聚合酶鏈?zhǔn)椒磻?yīng)(quantity real-time polymerase chain reaction,qRT-PCR)方法,觀察n-3 PUFA對(duì)實(shí)驗(yàn)組大鼠腸道菌群和脂肪因子的影響,探討魚(yú)油對(duì)大鼠腸道生物屏障的影響,為n-3 PUFA作為防治腸道相關(guān)疾病的營(yíng)養(yǎng)素提供新的理論依據(jù)。
1 材料與方法
1.1 動(dòng)物、材料與試劑
SPF級(jí)Wistar雌性大鼠購(gòu)自上海斯萊克實(shí)驗(yàn)動(dòng)物有限責(zé)任公司。
標(biāo)準(zhǔn)實(shí)驗(yàn)動(dòng)物飼料 濟(jì)南康大飼料有限公司;脫脂甜奶粉 瑞士雀巢公司;脫脂大豆粉 青島美辰食品公司;深海魚(yú)油 美國(guó)康隆公司;維生素A、D膠丸青島雙鯨藥業(yè)有限公司;其他試劑均為分析純。
各組飼料配方參照本實(shí)驗(yàn)室飼料制備配比見(jiàn)表1,飼料制備后4 ℃低溫保存。

表1 各組飼料成分配方Table 1 Ingredients of rat diets
1.2 儀器與設(shè)備
DW-86L626超低溫冰箱、BCD-196TE冰箱 青島海爾公司;SIGMA 1-13離心機(jī) 美國(guó)Sigma公司;PYXDHS隔水式電熱恒溫培養(yǎng)箱 上海躍進(jìn)醫(yī)療器械廠;RG-3000熒光定量PCR儀 英國(guó)Corbett Research公司;RM2235切片機(jī) 德國(guó)萊卡公司。
1.3 方法
1.3.1 動(dòng)物分組和實(shí)驗(yàn)設(shè)計(jì)
30 只SPF級(jí)5 周齡雌性Wistar大鼠,體質(zhì)量100~120 g,隨機(jī)分為正常對(duì)照組、n-3 PUFA組和n-6 PUFA組。正常對(duì)照組體質(zhì)量為(116.40±0.10)g;n-3 PUFA組體質(zhì)量為(109.33±0.28)g;n-6 PUFA組體質(zhì)量為(115.62±0.52)g。每組分為3 籠,自由攝食飲水。實(shí)驗(yàn)動(dòng)物房溫度(22±5)℃,相對(duì)濕度(50±10)%,明暗周期12 h/12 h。每周記錄每只大鼠的體質(zhì)量。第8周末時(shí)注射質(zhì)量分?jǐn)?shù)為10%的水合氯醛,處死大鼠,記錄每只大鼠肛門到鼻尖的長(zhǎng)度作為體長(zhǎng),無(wú)菌條件下取大鼠腸道糞便分裝于滅菌的5 mL離心管中,于-70 ℃保存。取大鼠肝臟于-70 ℃保存,取大鼠回腸、結(jié)腸、盲腸浸入體積分?jǐn)?shù)為10%的甲醛固定24 h,進(jìn)行石蠟包埋、切片,以備蘇木精-伊紅(hematoxylin-eosin,HE)染色。
1.3.2 糞便樣品的處理及腸道細(xì)菌總DNA的提取
糞便樣品處理參照參考文獻(xiàn)[1]。處理好的樣品于-20 ℃保存。
腸道細(xì)菌總DNA的提取采用十六烷基三甲基溴化銨法(hexadecyltrimethy ammonium bromide,CTAB)法提取,具體步驟參照參考文獻(xiàn)[2]。提取的DNA于-20 ℃保存。
1.3.3 肝臟總RNA提取
稱取50~100 mg肝臟組織,用Trizol(TaKaRa)提取,按試劑盒步驟進(jìn)行。提取的總mRNA用Roche反轉(zhuǎn)錄試劑盒進(jìn)行cDNA的合成。
1.3.4 聚合酶鏈?zhǔn)椒磻?yīng)(polymerase chain reaction,PCR)引物
雙歧桿菌(Bifidobacterium,Bif)、乳酸桿菌(Lactobacillus,Lac)引物參照參考文獻(xiàn)[3],腸道細(xì)菌Akkermansia muciniphila(Akk)引物參照參考文獻(xiàn)[4],由生工生物工程(上海)股份有限公司合成;禁食誘導(dǎo)脂肪因子(fasting-induced adipose factor,F(xiàn)IAF)和脂肪酸合成酶(fatty acid synthetase,F(xiàn)AS)引物由生工生物工程(上海)股份有限公司設(shè)計(jì)并合成,見(jiàn)表2。

表2 各引物序列Table 2 Sequence of primers used in this study
1.3.5 實(shí)時(shí)定量PCR
腸道菌群的實(shí)時(shí)定量PCR。25 μL反應(yīng)體系包括:12.5 μL SYBR Premix Ex Taq(Roche),200 nmol/L引物,1 μL DNA樣品。Bif、Lac反應(yīng)條件為:95 ℃ 5 min,95 ℃ 15 s,58 ℃ 30 s,72 ℃ 30 s,共40 個(gè)循環(huán),72 ℃延伸5 min,Akk退火溫度為60 ℃。每個(gè)樣本重復(fù)3 次,以2-ΔΔCt表示樣本中mRNA的相對(duì)表達(dá)量。
以提取的DNA樣本為模板,用各基因的特殊引物(表2)進(jìn)行普通PCR,得到PCR產(chǎn)物經(jīng)DNA片段純化試劑盒純化后與T載體連接,構(gòu)建質(zhì)粒轉(zhuǎn)化大腸桿菌。質(zhì)粒用DNA提取試劑盒提取后作為qRT-PCR的對(duì)照。
脂肪基因的實(shí)時(shí)定量PCR。25 μL反應(yīng)體系包括:12.5 μL SYBR Premix Ex Taq(Roche),200 nmol/L引物,500 ng RNA樣品。反應(yīng)條件為:95 ℃ 5 min,95 ℃ 15 s,60 ℃ 30 s,72 ℃ 30 s,共40 個(gè)循環(huán),72 ℃延伸5 min,每個(gè)樣本重復(fù)2 次,以2ΔΔ-Ct表示樣本中mRNA的相對(duì)表達(dá)量。
1.3.6 HE染色測(cè)量腸道黏膜厚度
取近結(jié)腸處約1 cm長(zhǎng),固定在體積分?jǐn)?shù)為10%的甲醛溶液中24 h后,用磷酸鹽緩沖液清洗24 h,分別浸入55%、65%、75%、85%、95%、100%乙醇中脫水,再放入二甲苯中,之后浸蠟包埋,切片進(jìn)行HE染色。每個(gè)部位最少測(cè)量與黏液層垂直的20 個(gè)厚度值,隨機(jī)選取每個(gè)結(jié)腸的5 個(gè)部位,用圖像分析系統(tǒng)(Image-Pro Plus 6.3)測(cè)量。
1.4 數(shù)據(jù)處理
結(jié)果以x±s表示,采用單因素方差分析(One-Way ANOVA)對(duì)每周體質(zhì)量增加量、體質(zhì)量/體長(zhǎng)等作統(tǒng)計(jì)學(xué)分析,用t檢驗(yàn)進(jìn)行各組間的兩兩比較。顯著性水平設(shè)為P<0.05。
2 結(jié)果與分析
2.1 體質(zhì)量、體長(zhǎng)的測(cè)定結(jié)果

圖1 3 組大鼠每周體質(zhì)量平均增加量和體質(zhì)量/體長(zhǎng)Fig.1 Comparisons of average body weight and weight/length ratio between rats fed high fat diet and those fed n-3 or n-6 PUFA-containing diet or common diet
由圖1可知, 3 組大鼠的每周體質(zhì)量增加量的平均值(F=5.374,P=0.013<0.05)差異有統(tǒng)計(jì)學(xué)意義;n-3 PUFA組大鼠的每周體質(zhì)量增加量的平均值顯著低于正常對(duì)照組(P<0.05);n-3 PUFA組的體質(zhì)量/體長(zhǎng)值增加趨勢(shì)緩慢。n-6 PUFA組和正常對(duì)照組之間無(wú)顯著性差異。表明n-3 PUFA組大鼠的肥胖程度顯著低于另外兩組。
2.2 實(shí)時(shí)定量結(jié)果
2.2.1 腸道菌群結(jié)果

圖2 3 組大鼠種腸道菌群的變化Fig.2 Changes in gut microbiota between rats fed high fat diet and those fed n-3 or n-6 PUFA-containing diet or common diet
由圖2可知,n-3 P U FA組和正常對(duì)照組相比,Bif、Lac和Akk均顯著增加,差異有統(tǒng)計(jì)學(xué)意義(t=2.229,P=0.047 6<0.05;t=2.653,P=0.021 0<0.05;t=1.680,P=0.001 3<0.05);而n-6 PUFA組的Bif雖也有所增加,但與正常對(duì)照組相比,二者之間仍無(wú)顯著性差異(P=0.099 6);n-6 PUFA組中Akk變化量低于正常對(duì)照組,但沒(méi)有統(tǒng)計(jì)學(xué)意義(P=0.121 2)。
2.2.2 肝臟脂肪代謝相關(guān)基因結(jié)果


圖3 肝臟中2 種脂肪因子的變化Fig.3 Changes in liver adi pocytokines
由圖3可知,3 個(gè)組FAS mRNA和FIAF mRNA之間的相對(duì)表達(dá)量差異有統(tǒng)計(jì)學(xué)意義(F=4.391,P=0.023 7<0.05;F=4.094,P=0.029 5<0.05);n- 3PUFA組FIAF與正常對(duì)照組相比表達(dá)量增加(P=0.040 3<0.05),與n-6 PUFA組相比無(wú)顯著性差異(P=0.0572);n-3 PUFA組FAS表達(dá)量減少與正常對(duì)照組相比差異有統(tǒng)計(jì)學(xué)意義(P=0.041 1<0.05);而n-6 PUFA組FAS表達(dá)量增加,和正常對(duì)照組相比有顯著性差異(P=0.010 3<0.05)。
2.3 腸道黏膜厚度
由圖4可知,在100×顯微鏡下獲取圖片,用圖像分析儀測(cè)量各組結(jié)腸腸道黏膜的厚度,差異有統(tǒng)計(jì)學(xué)意義(F=14.10,P<0. 01);n-3 PUFA組與正常對(duì)照組相比厚度增加(P=0.003 4<0.05);而n-6 PUFA組和正常對(duì)照組之間無(wú)顯著性差異(P=0.056 0)。
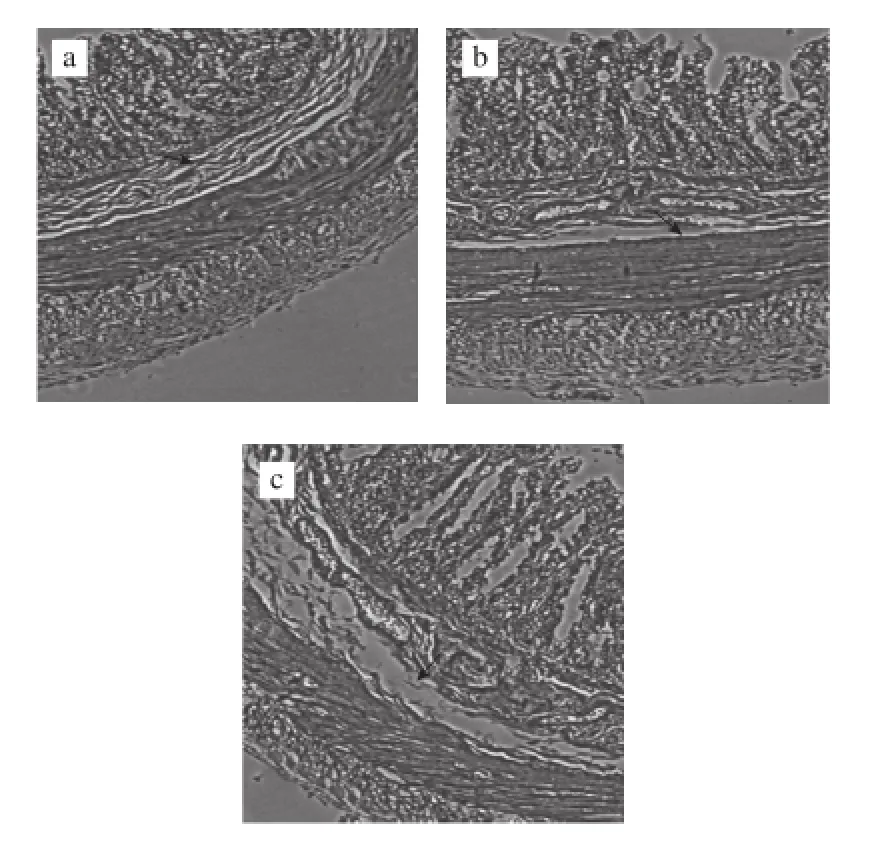
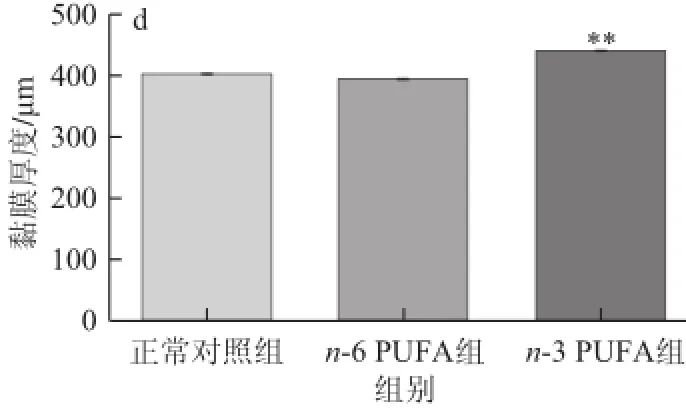
圖4 3 組腸道黏膜厚度的比較Fig.4 Comparison of intestinal mucus thickness among three groups
3 討 論
近年來(lái),n-3 PUFA對(duì)健康的影響越來(lái)越受到人們的重 視。n-3 PUFA是一類重要的多不飽和脂肪酸,主要包括EPA、DHA、二十二碳五烯酸、二十碳四烯酸、α-亞麻酸(alpha-linolenic acid,ALA)。其中EPA和DHA的營(yíng)養(yǎng)保健作用最有效。攝入一定量的EPA和DHA可以降低肥胖成年人體內(nèi)的脂肪含量,促進(jìn)脂肪的利用[5];增強(qiáng)2型糖尿病患者體內(nèi)胰島素的敏感性,緩解胰島素抵抗性[6],顯著降低糖尿病患者血清甘油三酯和血脂水平[7]。本實(shí)驗(yàn)中,長(zhǎng)期給大鼠喂食添加n-3 PUFA的食物后,可有效地控制其肥胖的發(fā)生,降低其肥胖程度,說(shuō)明EPA和DHA對(duì)大鼠有減肥作用,該結(jié)果與相關(guān)報(bào)道一致[5]。
腸道菌群結(jié)構(gòu)對(duì)維持腸道內(nèi)環(huán)境平衡起著非常重要的作用,人體是否健康與腸道內(nèi)的益生菌群結(jié)構(gòu)息息相關(guān),因此,人體在正常情況下,菌群結(jié)構(gòu)相對(duì)穩(wěn)定,對(duì)宿主表現(xiàn)為不致病。相反,如果腸道菌群失調(diào),會(huì)引起各種疾病。有研究指出,有益菌的比例在體格強(qiáng)健的人腸道內(nèi)達(dá)到70%,普通人則是25%,便秘人群減少到15%,而在癌癥病人腸道內(nèi)的比例只有10%。近年來(lái),隨著相關(guān)研究論文的發(fā)表,腸道菌群與腸道內(nèi)環(huán)境平衡和疾病的關(guān)系逐步清晰。有證據(jù)表明,腸道菌群可以調(diào)控脂肪代謝、引發(fā)低度慢性炎癥[8]和破壞腸道屏障[9-11],可以導(dǎo)致胰島素抵抗和肥胖癥的發(fā)生[12-13]。腸道菌群對(duì)腸黏膜系統(tǒng)的機(jī)械屏障、免疫屏障與生物屏障有調(diào)控作用。腸道中的大腸桿菌可以通過(guò)上調(diào)緊密連接蛋白ZO-1的表達(dá),減輕腸上皮的通透性[14]。乳酸桿菌可以對(duì)病原菌的定殖產(chǎn)生阻抗作用[15],能較好地恢復(fù)小鼠的腸黏膜免疫屏障功能[16]。雙歧桿菌可以有效減少乳糜瀉患者中由小麥醇溶蛋白引起的結(jié)腸上皮細(xì)胞的細(xì)胞膜皺褶[17],從而調(diào)控腸道的機(jī)械屏障。腸道菌群會(huì)隨著飲食的變化而變化,研究表明,飲食是決定腸道菌群構(gòu)成的最重要因素。因此,通過(guò)改變飲食,可以讓引起肥胖、糖尿病[18]、冠心病[19]的菌群結(jié)構(gòu)恢復(fù)正常,從而為預(yù)防、緩解甚至逆轉(zhuǎn)這些疾病帶來(lái)新的希望。本實(shí)驗(yàn)中,通過(guò)檢測(cè)n-3、n-6 PUFA對(duì)大鼠腸道菌群的影響發(fā)現(xiàn),n-3 PUFA可以增加腸道內(nèi)一些益生菌如Bif、Lac、Akk的含量,從而影響大鼠腸道內(nèi) 的菌群環(huán)境,改善大鼠的體內(nèi)微環(huán)境,調(diào)控腸道的屏障功能。
Akk是一種革蘭氏陰性菌,是疣微菌門的一種。近日,Akk被確定為駐留在黏液層的黏蛋白降解細(xì)菌,并且它在營(yíng)養(yǎng)豐富的環(huán)境中的人類細(xì)菌中占主導(dǎo)地位[20]。最新研究表明,Akk是一種減肥細(xì)菌,益生元(低聚果糖)能促進(jìn)其在腸 道內(nèi)的豐富度。它通過(guò)增加腸道黏膜的厚度來(lái)提高腸道黏膜屏障功能,維持葡萄糖穩(wěn)態(tài)和提高脂肪組織代謝。在飲食誘導(dǎo)的肥胖小鼠中Akk能改善代謝混亂,增加回腸中內(nèi)源性大麻素的水平,抵消肥胖中飲食引起的結(jié)腸黏膜屏障的功能紊亂[18]。在本實(shí)驗(yàn)中,n-3 PUFA組大鼠的腸道黏膜厚度顯著高于另外2 組,這可能與腸道菌群Akk的含量有關(guān)。進(jìn)一步說(shuō)明,n-3 PUFA可能通過(guò)改善腸道菌群,增加黏膜厚度,來(lái)提高腸道黏膜屏障功能。
FAS是脂肪酸合成的主要限制酶,它在乙酰輔酶A和丙二酸單酰輔酶A從頭合成長(zhǎng)鏈脂肪酸的過(guò)程中起催化作用。有研究指出,n-3 PUFA不僅能在蛋白水平抑制FAS的活性[21],而且還能在轉(zhuǎn)錄水平抑制FAS基因的表達(dá)[22-23]。另外,相關(guān)研究表明,腸道菌群作為內(nèi)化了的環(huán)境因子,可以直接調(diào)控動(dòng)物的基因表達(dá),它們可以增強(qiáng)肝臟合成脂肪的基因FAS的活性,同時(shí)關(guān)閉腸道內(nèi)抑制脂肪積累的基因FIAF[24]。FIAF也稱為血管生成素樣蛋白4,是過(guò)氧化物酶體增殖物激活受體的新靶點(diǎn)[25],是一種新的下丘腦調(diào)節(jié)器,它可以調(diào)節(jié)食物的攝取,從而影響體質(zhì)量[26]。本實(shí)驗(yàn)中,n-3 PUFA組FAS表達(dá)量減少,F(xiàn)IAF表達(dá)量增加,可能一方面通過(guò)n-3 PUFA直接影響基因的轉(zhuǎn)錄,另一方面可能通過(guò)n-3 PUFA影響腸道菌群的量來(lái)間接影響基因的合成。這說(shuō)明n-3 PUFA可能從多方面影響基因合成,從而達(dá)到減肥的目的。
綜上所述,本實(shí)驗(yàn)討論了n-3 PUFA對(duì)腸道菌群的影響及其對(duì)肥胖的抑制作用的可能途徑。這些結(jié)果提示n-3 PUFA可能通過(guò)改變腸道菌群中的有益菌,特別是Akk,改善腸道微環(huán)境,增加腸黏膜厚度,提高腸道黏膜屏障功能,并通過(guò)控制脂肪代謝相關(guān)基因的表達(dá)來(lái)達(dá)到減肥的目的。同時(shí),為人類使用這種腸道細(xì)菌用于預(yù)防或治療肥胖癥和其相關(guān)的代謝紊亂提供了新的治療方法。
[1] 張雪雁, 李琳琳. 一種提取腸道細(xì)菌總基因組DNA的方法[J]. 新疆醫(yī)科大學(xué)學(xué)報(bào), 2007, 30(7): 722-724.
[2] van PASSEL M W J, KANT R, ZOETENDAL E G, et al. The genome of Akkermansia muciniphila, a dedicated intestinal mucin degrader, and its use in exploring intestinal metagenomes[J]. PLoS One, 2011, 6(3): e16876.
[3] CANI P D, BIBILONI R, KNAUF C, et al. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice[J]. Diabetes, 2008, 57(6): 1470-1481.
[4] COLLADO M C, DERRIEN M, ISOLAURI E, et al. Intestinal integrity and Akkermansia muciniphila, a mucin-degrading member of the intestinal microbiota present in infants, adults, and the elderly[J]. Applied and Environmental Microbiology, 2007, 73(23): 7767-7770.
[5] MUNRO I A, GARG M L. Dietary supplementation with long chain omega-3 polyunsaturated fatty acids and weight loss in obese adults[J]. Obesity Research & Clinical Practice, 2013, 7(3): e173-e181.
[6] POPP-SNIJDERS C, SCHOUTEN J A, HEINE R J, et al. Dietary supplementation of omega-3 polyunsaturated fatty acids improves insulin sensitivity in non-insulin-dependent diabetes[J]. Diabetes Research (Edinburgh, Scotland), 1987, 4(3): 141-147.
[7] MONTORI V M, FARMER A, WOLLAN P C, et al. Fish oil supplementation in type 2 diabetes: a quantitative systematic review[J]. Diabetes Care, 2000, 23(9): 1407-1415.
[8] OSBORN O, OLEFSKY J M. The cellular and signaling networks linking the immune system and metabolism in disease[J]. Nature Medicine, 2012, 18(3): 363-374.
[9] CANI P D, AMAR J, IGLESIAS M A, et al. Metabolic endotoxemia initiates obesity and insulin resistance[J]. Diabetes, 2007, 56(7): 1761-1772.
[10] CANI P D, POSSEMIERS S, van de WIELE T, et al. Changes in gut microbiota control inflammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability[J]. Gut, 2009, 58(8): 1091-1103.
[11] MUCCIOLI G G, NASLAIN D, B?CKHED F, et al. The endocannabinoid system links gut microbiota to adipogenesis[J]. Molecular Systems Biology, 2010, 6(1). doi: 10.1038/msb.2010. 46.
[12] CANI P D, DELZENNE N M, AMAR J, et al. Role of gut microflora in the development of obesity and insulin resistance following high-fat diet feeding[J]. Pathologie Biologie, 2008, 56(5): 305-309.
[13] DING Shengli, CHI M M, SCULL B P, et al. High-fat diet: bacteria interactions promote intestinal inflammation which precedes and correlates with obesity and insulin resistance in mouse[J]. PLoS One, 2010, 5(8): e12191. doi:10.1371/journal.pone.0012191.
[14] ZYREK A A, CICHON C, HELMS S, et al. Molecular mechanisms underlying the probiotic effects of Escherichia coli Nissle 1917 involve ZO-2 and PKCzeta redistribution resulting in tight junction and epithelial barrier repair[J]. Cell Microbiol, 2007, 9(3): 804-816.
[15] LINDFORS K, BLOMQVIST T, JUUTI-UUSITALO K, et al. Live probiotic Bifidobacterium lactis bacteria inhibit the toxic effects induced by wheat gliadin in epithelial cell culture[J]. Clinical & Experimental Immunology, 2008, 152(3): 552-558.
[16] CANO P G, AGüERO G, PERDIGON G. Adjuvant effects of Lactobacillus casei added to a renutrition diet in a malnourished mouse model[J]. Biocell, 2002, 26(1): 35-48.
[17] CANDELA M, PERNA F, CARNEVALI P, et al. Interaction of probiotic Lactobacillus and Bifidobacterium strains with human intestinal epithelial cells: adhesion properties, competition against enteropathogens and modulation of IL-8 production[J]. International Journal of Food Microbiology, 2008, 125(3): 286-292.
[18] EVERARD A, BELZER C, GEURTS L, et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls dietinduced obesity[J]. Proceedings of the National Academy of Sciences, 2013, 110(22): 9066-9071.
[19] STOCK J. Gut microbiota: an environmental risk factor for cardiovascular disease[J]. Atherosclerosis, 2013, 229(2): 440-442.
[20] DERRIEN M, VAUGHAN E E, PLUGGE C M, et al. Akkermansia mucinip hila gen. nov., sp. nov., a human intestinal mucin-degrading bacterium[J]. International Journal of Systematic and Evolutionary Microbiology, 2004, 54(5): 1469-1476.
[ 21] CLARKE S D, HEMBREE J. Inhibition of triiodothyronine’s induction of rat liver lipogenic enzymes by dietary fat[J]. The Journal of Nutrition, 1990, 120(6): 625-630.
[22] CLARKE S D. Regulation of fatty acid synthase gene expression: an approach for reducing fat accumulation[J]. Journal of Animal Sci ence, 1993, 71(7): 1957-1965.
[23] 時(shí)皎皎, 糜漫天, 韋娜, 等. 不同脂肪酸構(gòu)成比對(duì)大鼠血脂影響的研究[J]. 第三軍醫(yī)大學(xué)學(xué)報(bào), 2007, 29(9): 824-827.
[24] B?CKHED F, DING Hao, WANG Ting, et al. The gut microbiota as an environmental factor that regulates fat stora ge[J]. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(44): 15718-15723.
[25] KERSTEN S, MANDARD S, TAN N S, et al. Characterization of the fasting-induced adipose factor FIAF, a novel peroxisome proliferatoractivated receptor target gene[J]. Journal of Biological Chemistry, 2000, 275(37): 28488-28493.
[26] KIM H K, YOUN B S, SHIN M S, et al. Hypothalamic Angptl4/Fiaf is a novel regulator of food intake and body weight[J]. Diabetes, 2010, 59(11): 2772-2780.
Effect of PUFA-Containing Diet on Gut Microbiota and Fat Metabolism-Related Genes in Rats
QIAO Li-jun, ZHENG Zheng, MA Wen-hui, LU Ting-xiu, GAI Xiao-ying, GE Yin-lin*
(Department of Biochemistry and Molecular Biology, College of Medical, Qingdao University, Qingdao 266021, China)
Purpose: To determine the effect of PUFA diet on gut microbiota and obesity-related cell factors in rats. Me thods: A total of 30 five-week-old healthy female rats were randomly divided into three groups: control group, n-6 PUFA-containing diet group and n-3 PUFA-containing diet group. The rats had free access to the feeds and water for eight weeks. Body weight was recorded once a week during the trail period and the levels of Bifi dobacterium (Bif), Lactobacillus (Lac), Akkermansia muciniphila (Akk) and obesity-related cell factors, fatty acid synthetase (FAS) and fasting-induced adipose factor (FIAF) were evaluated by qRT-PCR at the end of the trial. Results: The adiposity and FAS level in the n-3 PUFA-containing diet group were significantly lower than that in the control group (P < 0.05), and the levels of Bif, Lac, Akk and FIAF, and the thickness of the intestinal mucosa in the n-3 PUFA-containing diet model group were significantly higher than those in the control group (P < 0.05). Conclusion: An obvious change in gut microbiota was observed in rats fed n-3 PUFA-containing diet; meanwhile the occurrence and development of adiposity were inhibited.
n-3 PUFA; obesity; Akkermansia muciniphila; intestinal mucosa
R378
A
1002-6630(2014)17-0231-05
10.7506/spkx1002-6630-201417044
2013-10-12
山東省自然科學(xué)基金項(xiàng)目(ZR2010CM010)
喬立君 (1988—),女,碩士研究生,研究方向?yàn)榛蛑委煛-mail:qlj19880728@163.com
*通信作者:葛銀林(1957—),男,教授,博士,研究方向?yàn)榛蛟\斷與治療。E-mail:geyinlin@126.com
猜你喜歡
中學(xué)生數(shù)理化·中考版(2022年10期)2022-11-10 09:37:42
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年12期)2022-02-14 07:08:42
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2021年12期)2021-12-31 03:23:08
中學(xué)生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
中國(guó)生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(shè)(2018年6期)2018-04-22 03:16:54
產(chǎn)品可靠性報(bào)告(2017年7期)2017-09-05 09:49:12
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
