稀土金屬鈰對(duì)黃豆種子萌發(fā)及幼苗生理活性的影響
2014-02-18 08:38:24趙倩倩趙朝宇徐秋曼郭紅梅
趙倩倩,趙朝宇,徐秋曼,郭紅梅,陳 宏
(天津師范大學(xué)a.生命科學(xué)學(xué)院,b.天津市動(dòng)植物抗性重點(diǎn)實(shí)驗(yàn)室,天津300387)
稀土元素對(duì)植物的生長(zhǎng)發(fā)育有促進(jìn)作用,主要表現(xiàn)在2個(gè)方面:一方面是稀土元素能夠改善植物的光合器官和活性,提高植物體內(nèi)的酶活性,從而提高植物的光合作用能力[1-3];另一方面能夠促進(jìn)種子萌發(fā),提高種子的出苗率.稀土元素促進(jìn)種子萌發(fā)的作用已成功應(yīng)用在小麥等大田作物上,如小麥發(fā)芽率可提高8%~19%,胡麻發(fā)芽率可提高7%~12%[4].種子萌發(fā)作為種子植物生活史的第一階段對(duì)植物生長(zhǎng)發(fā)育至關(guān)重要[5].有關(guān)稀土金屬對(duì)植物生長(zhǎng)發(fā)育影響的報(bào)道以元素鑭居多,如郜紅建等[6]研究鑭對(duì)水稻根的影響,發(fā)現(xiàn)低濃度的鑭可以使細(xì)胞膜穩(wěn)定,降低電解質(zhì)的外滲率.對(duì)于鈰(Ce)的研究以三價(jià)居多,如王蕊等[7]研究發(fā)現(xiàn)CeCl3可以提高豌豆細(xì)胞清除活性氧的能力,有效緩解銅脅迫對(duì)豌豆的傷害.
本研究采用梯度濃度的Ce(NH4)2(NO3)6溶液處理黃豆種子,測(cè)定種子的萌發(fā)率和活力、幼苗的淀粉酶活性、根系活力、葉綠素含量、相對(duì)電導(dǎo)率以及丙二醛(MDA)含量等指標(biāo),以此研究鈰對(duì)黃豆種子萌發(fā)和生長(zhǎng)的影響.這不但可以為探討不同鈰化合物對(duì)植物生長(zhǎng)發(fā)育的影響提供理論依據(jù),同時(shí)由于黃豆是我國(guó)重要的經(jīng)濟(jì)作物,因此本項(xiàng)研究還具有一定的經(jīng)濟(jì)意義.
1 實(shí)驗(yàn)材料與方法
1.1 實(shí)驗(yàn)處理
選取外形飽滿的黃豆種子(國(guó)審中黃19),用質(zhì)量分?jǐn)?shù)為0.1%的升汞消毒10min,蒸餾水沖洗,吸干.在25℃下分別用不同質(zhì)量濃度(0.1、1.0、10.0、50.0、100.0mg/L)的Ce(NH4)2(NO3)6溶液浸種24h,以蒸餾水作為對(duì)照,再于28℃下催芽24h.挑選發(fā)芽一致的種子,播種于放有濕潤(rùn)濾紙的培養(yǎng)皿(Φ=10cm)中,每皿15粒,置于25℃/20℃、每天光照12h的培養(yǎng)室中培養(yǎng).第3天統(tǒng)計(jì)生長(zhǎng)勢(shì)和萌發(fā)率,測(cè)定淀粉酶活性.分別在第4、8、12天測(cè)定幼苗的葉綠素含量.從第3天開始每天測(cè)定細(xì)胞膜相對(duì)電導(dǎo)率、MDA含量,連續(xù)測(cè)量至第8天.在發(fā)芽的第7天測(cè)定不同質(zhì)量濃度的Ce(NH4)2(NO3)6溶液對(duì)黃豆幼苗根系活力的影響.每個(gè)測(cè)量重復(fù)5次.
1.2 測(cè)定方法
種子萌發(fā)率、生長(zhǎng)勢(shì)、種子活力的測(cè)定方法參照文獻(xiàn)[8].生長(zhǎng)勢(shì)為測(cè)試種子的平均根長(zhǎng)度,胚芽長(zhǎng)度大于種子長(zhǎng)度1/3者視為萌發(fā),萌發(fā)率與生長(zhǎng)勢(shì)之乘積為種子活力,設(shè)對(duì)照處理的種子活力為100%,其他處理的種子活力與對(duì)照處理的種子活力的比值為活力比.用丙酮提取法測(cè)定葉綠素含量,以分光光度法在645nm和663 nm處測(cè)定提取液的光密度[9].淀粉酶活性采用3,5-二硝基水楊酸還原法測(cè)定[9];細(xì)胞膜相對(duì)透性采用電導(dǎo)法;MDA的測(cè)定參照文獻(xiàn)[10].
1.3 數(shù)據(jù)處理
采用SPSS17.0和Excel軟件對(duì)實(shí)驗(yàn)數(shù)據(jù)進(jìn)行統(tǒng)計(jì)分析.
2 結(jié)果與分析
2.1 鈰對(duì)黃豆種子萌發(fā)及活力的影響
黃豆種子經(jīng)不同質(zhì)量濃度的Ce(NH4)2(NO3)6溶液處理后,種子的萌發(fā)和生長(zhǎng)情況如表1所示.
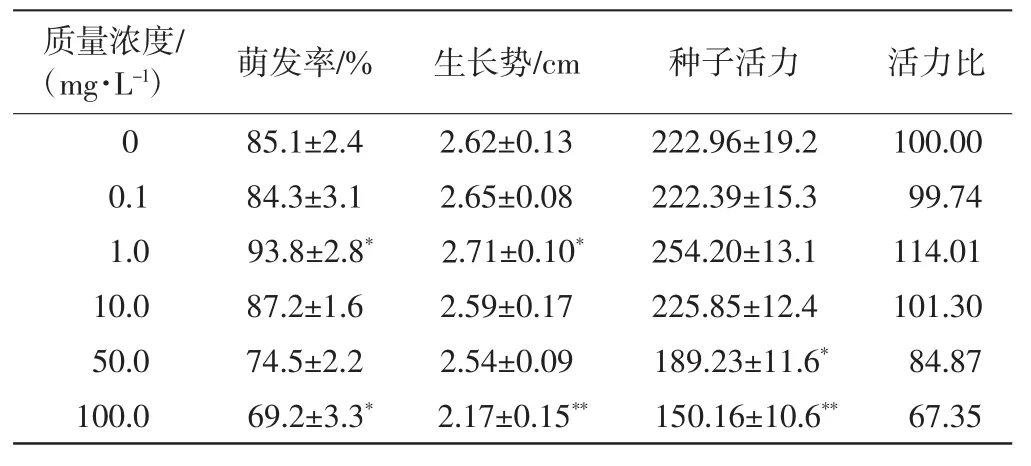
表1 不同質(zhì)量濃度的Ce(NH4)2(NO3)6處理下黃豆種子的萌發(fā)和活力Tab.1 Seed germination and vigor treated with different concentrations of Ce(NH4)2(NO3)6
由表1可以看出,當(dāng)質(zhì)量濃度較低(1.0、10.0mg/L)時(shí),Ce(NH4)2(NO3)6對(duì)黃豆種子的萌發(fā)率以及生長(zhǎng)勢(shì)具有促進(jìn)作用,質(zhì)量濃度為1.0mg/L時(shí)促進(jìn)效果最明顯,萌發(fā)率比對(duì)照組高出10.2%,生長(zhǎng)勢(shì)比對(duì)照組高出3.4%,種子活力比對(duì)照組提高了14.0%.當(dāng)質(zhì)量濃度過高(50.0、100.0mg/L)時(shí),Ce(NH4)2(NO3)6對(duì)黃豆種子的萌發(fā)和活力產(chǎn)生抑制作用,質(zhì)量濃度為100mg/L時(shí),種子的萌發(fā)率和生長(zhǎng)勢(shì)分別比對(duì)照組下降18.7%和17.2%,種子活力也下降了32.7%,萌發(fā)率與對(duì)照組相比差異具有統(tǒng)計(jì)學(xué)意義(p≤0.05),生長(zhǎng)勢(shì)與對(duì)照組相比差異具有高度統(tǒng)計(jì)學(xué)意義(p≤0.01).
2.2 鈰對(duì)黃豆幼苗淀粉酶活性的影響
種子中儲(chǔ)存有大量的淀粉類物質(zhì),種子萌發(fā)時(shí),在淀粉酶的作用下淀粉類物質(zhì)被水解為簡(jiǎn)單的有機(jī)物,成為幼胚生長(zhǎng)的營(yíng)養(yǎng)物質(zhì)來源.淀粉酶的活性會(huì)直接影響幼胚營(yíng)養(yǎng)物質(zhì)供應(yīng)的速度,進(jìn)而影響種子萌發(fā)及其生理活性[11].測(cè)定不同濃度的Ce(NH4)2(NO3)6處理下黃豆種子中的淀粉酶活性,結(jié)果如圖1所示.
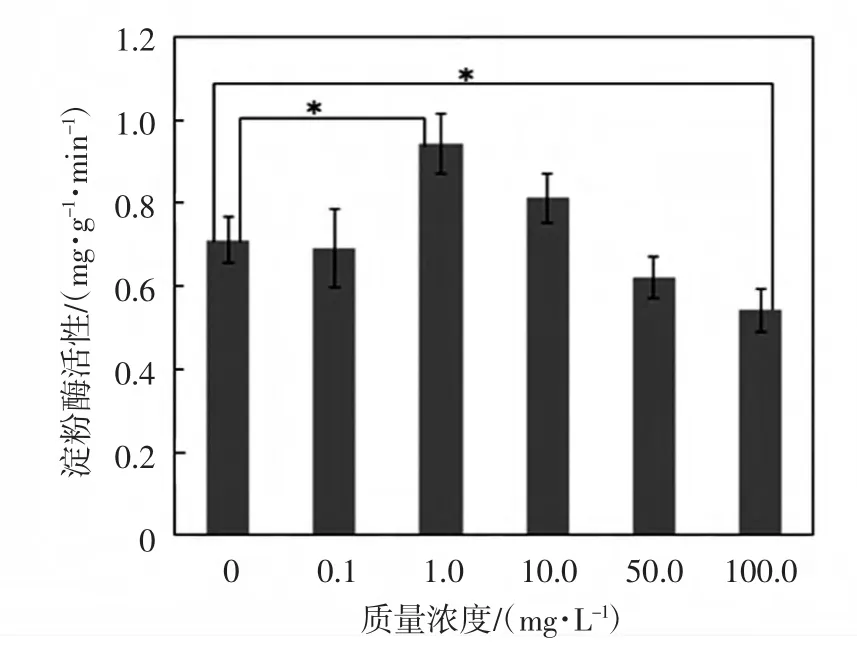
圖1 鈰對(duì)黃豆種子淀粉酶活性的影響Fig.1 Effects of Ce on diastase activity in soybean seed
由圖1可知,Ce(NH4)2(NO3)6質(zhì)量濃度為1.0mg/L和10mg/L時(shí),鈰對(duì)淀粉酶活性有不同程度的促進(jìn)作用,其他濃度的鈰會(huì)抑制淀粉酶活性.當(dāng)Ce(NH4)2(NO3)6質(zhì)量濃度為1.0mg/L時(shí),淀粉酶活性比對(duì)照組高出32.4%,差異具有統(tǒng)計(jì)學(xué)意義(p≤0.05);質(zhì)量濃度為50.0mg/L和100.0mg/L時(shí),鈰對(duì)淀粉酶活性有抑制作用,分別比對(duì)照組降低12.7%和23.9%,100.0mg/L處理組與對(duì)照組之間的差異具有統(tǒng)計(jì)學(xué)意義(p≤0.05).
2.3 鈰對(duì)黃豆幼苗根系活力的影響
根系活力是衡量植物生長(zhǎng)能力大小的重要指標(biāo)[12].在發(fā)芽的第7天,測(cè)定不同濃度的Ce(NH4)2(NO3)6浸種對(duì)黃豆幼苗根系活力的影響,結(jié)果如圖2所示.由圖2可以看出,當(dāng)Ce(NH4)2(NO3)6質(zhì)量濃度為1.0mg/L和10.0mg/L時(shí),黃豆幼苗的根系活力比對(duì)照組高,濃度為1.0mg/L時(shí)根系活力最大,比對(duì)照組高34.3%,差異具有統(tǒng)計(jì)學(xué)意義(p≤0.05);其他濃度下,鈰處理會(huì)降低黃豆幼苗的根系活力,尤其是當(dāng)質(zhì)量濃度為100.0mg/L時(shí),根系活力同對(duì)照組相比降低16.9%,差異具有統(tǒng)計(jì)學(xué)意義(p≤0.05),說明高濃度的鈰對(duì)根系活力會(huì)產(chǎn)生抑制作用.
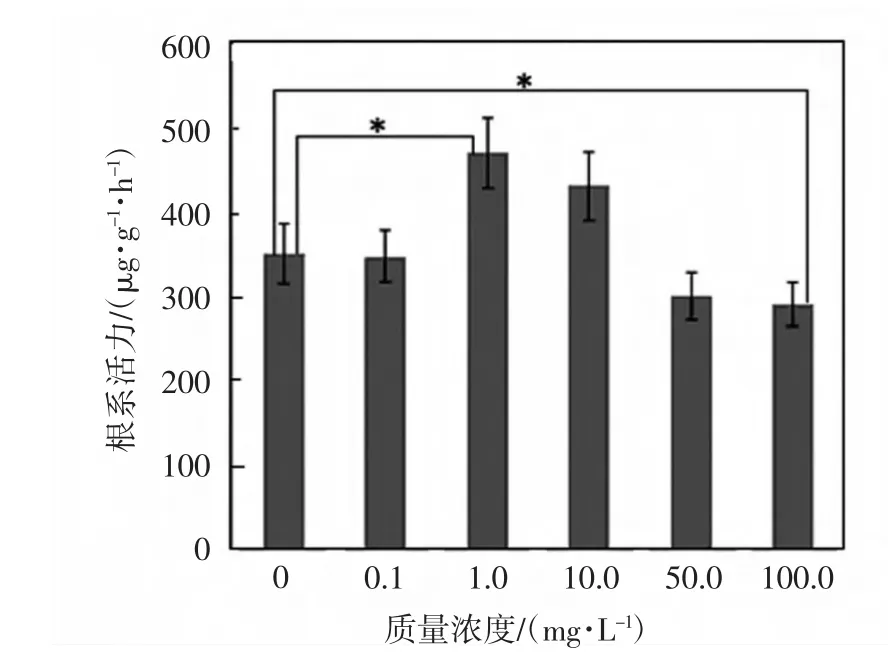
圖2 鈰對(duì)黃豆幼苗根系活力的影響Fig.2 Effects of Ce on root activity of soybean seedlings
2.4 鈰對(duì)黃豆幼苗葉綠素含量的影響
測(cè)定不同濃度的Ce(NH4)2(NO3)6處理下黃豆幼苗中葉綠素的含量,結(jié)果如表2所示.

表2 鈰對(duì)黃豆幼苗葉綠素含量的影響Tab.2 Effects of Ce on chlorophyll content in soybean seedlings
由表2可以看出,黃豆幼苗的葉綠素含量隨培養(yǎng)時(shí)間和鈰處理濃度的變化而不同.從培養(yǎng)時(shí)間來看,除了100.0mg/L處理組在培養(yǎng)時(shí)間為4d時(shí)葉綠素含量最高以外,其他處理組均在8d時(shí)葉綠素a、b和總?cè)~綠素含量達(dá)到最高,隨著培養(yǎng)時(shí)間的延長(zhǎng),黃豆幼苗中的葉綠素含量大幅降低.從鈰處理濃度來看,低濃度的Ce(NH4)2(NO3)6(質(zhì)量濃度≤10.0mg/L)處理會(huì)增加幼苗的葉綠素含量,尤其是質(zhì)量濃度為1.0mg/L的處理,培養(yǎng)時(shí)間為8d時(shí),葉綠素a、b和總?cè)~綠素分別比對(duì)照組增加33.6%、36.9%和34.4%;高濃度的Ce(NH4)2(NO3)6(質(zhì)量濃度≥50.0mg/L)會(huì)降低葉綠素含量,尤其是100.0mg/L的處理組,8d時(shí)葉綠素a、b和總?cè)~綠素含量分別比對(duì)照組降低了38.4%、11.5%、32.3%.
2.5 鈰對(duì)黃豆幼苗相對(duì)電導(dǎo)率的影響
植物細(xì)胞原生質(zhì)膜的穩(wěn)定性一方面可以保證植物細(xì)胞完成物質(zhì)轉(zhuǎn)運(yùn)、能量交換和信息傳遞等功能,同時(shí)也有助于維持細(xì)胞內(nèi)外離子濃度的平衡.種子萌發(fā)過程中,相對(duì)電導(dǎo)率可以反映膜系統(tǒng)的完整性,其值越大,表示電解質(zhì)的滲漏量越多,膜受損程度越大[13].不同濃度的Ce(NH4)2(NO3)6處理下黃豆幼苗相對(duì)電導(dǎo)率隨時(shí)間的變化如圖3所示.
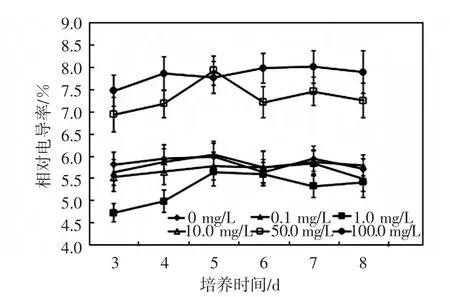
圖3 鈰對(duì)黃豆幼苗相對(duì)電導(dǎo)率的影響Fig.3 Effects of Ce on relative conductivity of soybean seedlings
由圖3可以看出,低濃度的鈰(質(zhì)量濃度≤10.0mg/L)可以降低黃豆幼苗的相對(duì)電導(dǎo)率,尤其是1.0mg/L處理組,幼苗的相對(duì)電導(dǎo)率在培養(yǎng)時(shí)間內(nèi)始終低于其他濃度處理組,其次是10.0mg/L處理組,幼苗的相對(duì)電導(dǎo)率也低于對(duì)照;高濃度的鈰(質(zhì)量濃度≥50.0mg/L)則會(huì)大幅提高幼苗的相對(duì)電導(dǎo)率,尤其是100.0mg/L處理組,顯著高于其他處理組.從培養(yǎng)時(shí)間來看,培養(yǎng)3 d時(shí)幼苗的相對(duì)電導(dǎo)率都處于該處理組下最低水平,隨著培養(yǎng)時(shí)間的延長(zhǎng),相對(duì)電導(dǎo)率增加,通常在4~6d時(shí)出現(xiàn)峰值,此后相對(duì)電導(dǎo)率緩慢下降.
2.6 鈰對(duì)黃豆幼苗MDA含量的影響
植物在逆境條件或者器官衰老階段,細(xì)胞膜脂質(zhì)中的不飽和脂肪酸會(huì)發(fā)生過氧化作用,MDA大量產(chǎn)生,最終導(dǎo)致質(zhì)膜透性的增大.MDA含量可以作為衡量細(xì)胞膜脂質(zhì)過氧化程度的指標(biāo)[14].測(cè)量不同濃度的鈰處理下黃豆幼苗中MDA的含量,結(jié)果如圖4所示.

圖4 鈰對(duì)黃豆幼苗MDA含量的影響Fig.4 Effects of Ce on MDA content in soybean seedlings
由圖4可以看出,所有處理組中,只有1.0mg/L處理組的MDA在整個(gè)培養(yǎng)階段始終低于對(duì)照組,0.1mg/L和10.0mg/L處理組幼苗的MDA與對(duì)照組相近,高濃度處理組(質(zhì)量濃度≥50.0mg/L)的MDA在培養(yǎng)階段則遠(yuǎn)高于對(duì)照組,而且100.0mg/L處理組中MDA含量最多.由此可見,低濃度的鈰會(huì)降低細(xì)胞中MDA含量,即降低質(zhì)膜中不飽和脂肪酸的過氧化作用,減小膜透性;高濃度的鈰則會(huì)增加MDA的含量,使膜透性增加.
3 討論
植物對(duì)稀土元素的吸收有一個(gè)臨界含量,當(dāng)外源稀土施用量達(dá)到適宜濃度時(shí),對(duì)植物的生長(zhǎng)發(fā)育才有促進(jìn)作用,當(dāng)稀土元素濃度超過臨界含量時(shí),植物生長(zhǎng)將受到抑制甚至毒害.如任曉曉[15]研究Ce(Ⅲ)對(duì)UV-B輻射脅迫下大豆礦質(zhì)元素含量與分布的影響,發(fā)現(xiàn)低濃度CeCl3溶液對(duì)大豆生長(zhǎng)有一定的促進(jìn)作用,而高濃度則產(chǎn)生抑制作用;任紅玉等[16]研究鑭和鈰對(duì)大豆開花期葉綠素含量的影響,發(fā)現(xiàn)CeCl3的濃度過高(60~150mg/L)時(shí)會(huì)降低葉綠素合成,低濃度鈰(20mg/L)能夠促進(jìn)葉綠素合成.
本研究采用不同濃度的Ce(NH4)2(NO3)6對(duì)黃豆種子進(jìn)行浸種,觀察不同濃度的鈰對(duì)種子萌發(fā)和幼苗生長(zhǎng)的影響.發(fā)現(xiàn)不同濃度的Ce(NH4)2(NO3)6處理黃豆種子,同樣存在低促高抑現(xiàn)象.質(zhì)量濃度為1.0mg/L時(shí)的促進(jìn)效果最好,黃豆種子的萌發(fā)率、生長(zhǎng)勢(shì)和種子活力分別比對(duì)照組高出10.2%、3.4%和14.0%,黃豆幼苗淀粉酶活性的增幅最大,比對(duì)照組增加32.4%,幼苗葉綠素含量也有所增加,同時(shí)電導(dǎo)率和MDA含量均顯著低于對(duì)照組;當(dāng)質(zhì)量濃度大于50mg/L時(shí),鈰對(duì)黃豆種子萌發(fā)及幼苗生理活性有較強(qiáng)的抑制作用.
[1] 何躍君,薛立.稀土元素對(duì)植物的生物效應(yīng)及其作用機(jī)理[J].應(yīng)用生態(tài)學(xué)報(bào),2005,16(10):1983-1989.
[2] 李永裕,潘騰飛,邱棟梁.稀土元素對(duì)植物生物學(xué)作用機(jī)制的研究進(jìn)展[J].中國(guó)農(nóng)學(xué)通報(bào),2005,21(12):217-221.
[3] 許寶泉,吳水英.稀土金屬對(duì)植物生長(zhǎng)影響的研究進(jìn)展[J].廣西輕工業(yè),2006(5):62-67.
[4] 劉蓉生.稀土農(nóng)用[J].四川稀土,2004(2):30-32.
[5] 呂篤康,李海冰,巴音山,等.重金屬鉻及鹽脅迫對(duì)小香蒲種子萌發(fā)的影響[J].種子,2013,32(2):72-75.
[6] 郜紅建,常江,張自立,等.鑭對(duì)水稻根質(zhì)膜透性和根分泌物中幾種營(yíng)養(yǎng)離子含量的影響[J].應(yīng)用生態(tài)學(xué)報(bào),2004,15(4):651-654.
[7] 王蕊,王應(yīng)軍,馬星宇.鑭、鈰對(duì)銅脅迫下豌豆幼苗抗氧化酶系統(tǒng)的影響[J].核農(nóng)學(xué)報(bào),2013,27(6):873-878.
[8] 陶嘉齡,鄭光華.種子活力[M].北京:科學(xué)出版社,1991:109-113.
[9] 鄒琦.植物生理生化實(shí)驗(yàn)指導(dǎo)[M].北京:中國(guó)農(nóng)業(yè)出版社,1995.
[10]SHIMAZAKI K N,SAKAKI K N,SUGAHARA K.Active oxygen participation in chlorophyll destruction and lipid peroxidation in SO2fumigated leaves of spinach[J].Plant Cell Physiology,1980,21(7):1193-1204.
[11]劉登義,王友保.Cu、As對(duì)作物種子萌發(fā)和幼苗生長(zhǎng)影響的研究[J].應(yīng)用生態(tài)學(xué)報(bào),2002,13(2):179-182.
[12]何奇江,李楠,傅懋毅,等.氯化鈉脅迫對(duì)雷竹根系活力和細(xì)胞膜透性的影響[J].浙江農(nóng)林大學(xué)學(xué)報(bào),2013,30(6):944-949.
[13]徐秋曼,陳宏,程景勝.稀土元素鈰減輕鉛對(duì)玉米毒害的機(jī)理初探[J].西南大學(xué)學(xué)報(bào):自然科學(xué)版,2008,30(3):70-74.
[14]XU Q M,CHEN H.Antioxidant responses of rice seedling to Ce4+under hydroponic cultures[J].Ecotoxicology and Environmental Safety,2011,74(6):1693-1699.
[15]任曉曉.Ce(Ⅲ)對(duì)UV-B輻射脅迫下大豆礦質(zhì)元素含量與分布的影響[D].無錫:江南大學(xué),2008.
[16]任紅玉,劉曦,朱曉鑫,等.鑭和鈰對(duì)大豆開花期葉綠素含量的影響[J].中國(guó)稀土學(xué)報(bào),2013,31(3):363-367.
