饑餓對條紋鋸 仔魚的存活、攝食及生長的影響
2014-03-07 07:12:56賈瑞錦孔祥迪李炎璐宋振鑫吳雷明曲江波趙從明
海洋科學(xué) 2014年4期
賈瑞錦 , 陳 超 , 孔祥迪, 李炎璐 , 宋振鑫, 吳雷明,王 魯 曲江波, 趙從明, 吳 堅
(1. 中國水產(chǎn)科學(xué)研究院 黃海水產(chǎn)研究所, 山東 青島 266071; 2. 上海海洋大學(xué) 水產(chǎn)與生命學(xué)院, 上海 201306; 3. 煙臺開發(fā)區(qū)天源水產(chǎn)有限公司, 山東 煙臺 264000; 4. 天津立達(dá)海水資源開發(fā)有限公司, 天津 300050; 5. 浙江海生源生物科技有限公司, 浙江 臺州 317600)
饑餓對條紋鋸 仔魚的存活、攝食及生長的影響
賈瑞錦1,2, 陳 超1,2, 孔祥迪1,2, 李炎璐1,2, 宋振鑫1,2, 吳雷明1,2,王 魯1, 曲江波3, 趙從明4, 吳 堅5
(1. 中國水產(chǎn)科學(xué)研究院 黃海水產(chǎn)研究所, 山東 青島 266071; 2. 上海海洋大學(xué) 水產(chǎn)與生命學(xué)院, 上海 201306; 3. 煙臺開發(fā)區(qū)天源水產(chǎn)有限公司, 山東 煙臺 264000; 4. 天津立達(dá)海水資源開發(fā)有限公司, 天津 300050; 5. 浙江海生源生物科技有限公司, 浙江 臺州 317600)
為了確定條紋鋸 (Centropristis striata)仔魚的不可逆點(PNR), 作者在溫度(22.9±0.8)℃、鹽度30~32、DO≥5.0mg/L、pH7.8~8.2, 光照強度40~100 Lx條件下, 對人工孵化的條紋鋸 仔魚開展饑餓脅迫對其存活、攝食、生長的變化規(guī)律的試驗研究及影響。結(jié)果顯示, 仔魚在孵化后2 d開口攝食, 仔魚體長開始下降, 從內(nèi)源營養(yǎng)期轉(zhuǎn)入混合營養(yǎng)期, 持續(xù)1 d; 3 d時卵黃囊、油球消失, 仔魚體長增加, 進(jìn)入外源營養(yǎng)期, 初次攝食率可達(dá)到68.25%。3.5 d時初次攝食率最高達(dá)到88.24%, 之后初次攝食率逐漸下降, 5 d時初次攝食率下降為37.5%, 可知條紋鋸 PNR=5 d。6.5 d時試驗組仔魚全部死亡。結(jié)果表明,內(nèi)源營養(yǎng)期對照組與試驗組無顯著差異, 對卵黃囊與油球的利用率相似。混合營養(yǎng)期, 仔魚體長出現(xiàn)負(fù)增長, 且試驗組負(fù)增長速率明顯高于對照組。外源營養(yǎng)期, 試驗組仔魚體長持續(xù)下降, 至 PNR時體長負(fù)增長速率加快死亡率增加, 直至全部死亡; 對照組仔魚體長開始增長, 死亡率逐漸下降并趨于穩(wěn)定。條紋鋸 仔魚孵出后2~5 d為攝食的關(guān)鍵時期。
條紋鋸 (Centropristis striata)仔魚; 饑餓; 攝食; 不可逆點
條紋鋸 (Centropristis striata)屬于 科(Serranidae), 石斑魚亞科(Serraninae), 鋸 屬(Centropristis), 常用名美洲黑石斑魚。條紋鋸 作為名貴海水養(yǎng)殖魚類, 具有肉質(zhì)豐腴, 口感鮮軟清爽[1],適應(yīng)性強等優(yōu)點[2-5], 深受廣大消費者和養(yǎng)殖戶的喜愛。中國自2002年[6]從美國引進(jìn)魚苗和受精卵進(jìn)行養(yǎng)殖和育苗試驗至今, 條紋鋸 的規(guī)模化養(yǎng)殖已經(jīng)在山東、浙江、福建、廣東等地興起, 取得了一定的經(jīng)濟效益。國內(nèi)、外對條紋鋸 都有研究報道,如條紋鋸 胚胎發(fā)育[7]、幼魚配合飼料的研究[8]、魚種對鹽度脅迫的耐受力[9]、海水網(wǎng)箱養(yǎng)殖試驗[10]等, 目前未見有關(guān)饑餓對條紋鋸 仔魚發(fā)育影響的研究報道。
自然界中, 由于食物分布在時間、空間上的不均及環(huán)境劇變等, 魚類經(jīng)常會在生活周期的一定階段面臨食物資源的缺乏而受到饑餓脅迫[11-15]。在人工育苗過程中, 特別是仔魚階段經(jīng)常由于餌料種類、數(shù)量、適口性及其轉(zhuǎn)換等因素引起營養(yǎng)不良導(dǎo)致死亡,因此有關(guān)饑餓對魚類早期生長發(fā)育的研究對育苗生產(chǎn)具有重要的指導(dǎo)意義。自 Blaxter等[16]提出“不可逆點”(the point-of-no-return, PNR)的概念即仔魚攝食前耐受饑餓的時間臨界點, 從生態(tài)學(xué)的角度研究了仔魚對饑餓的耐受能力, 國內(nèi)外對饑餓對牙鲆(Paralichthys olivaceus)[17]、半滑舌鰨(Cynoglossus semilaev is)[18]、條石鯛(Oplegnathus fasciatus)[19-21]、點帶石斑魚(Epinephelus malabaricus)[22]、斜帶石斑魚(Epinephelus coioides)[23]的影響進(jìn)行了大量的研究。作者通過饑餓對條紋鋸 仔魚的存活、攝食及生長的實驗研究及其影響分析, 以期解決條紋鋸 苗種生產(chǎn)過程中的開口餌料的營養(yǎng)利用、內(nèi)外源營養(yǎng)的轉(zhuǎn)換及饑餓對其存活和生長的影響等問題, 為其苗種生產(chǎn)提供理論依據(jù)。
1 材料與方法
1.1 仔魚來源及飼養(yǎng)
試驗材料為2012年5月取自中國水產(chǎn)科學(xué)研究院黃海水產(chǎn)研究所暫養(yǎng)在煙臺開發(fā)區(qū)天源水產(chǎn)有限公司的條紋鋸 親魚自然產(chǎn)卵獲得的受精卵, 收集、孵化、計數(shù)、分組。仔魚孵化后分兩組用80 L長方形塑料桶飼養(yǎng), 每組設(shè)3個平行試驗, 每桶2 000尾初孵仔魚, 第一組用做饑餓試驗(試驗組), 不予投餌;第二組每天正常投餌(對照組), 仔魚在開口前一天開始投喂經(jīng)小球藻(Chlorella vulgaris)強化的ss輪蟲(Brachionus plicatilis), 密度為10~15個/mL。培育水溫(22.9±0.8)℃, 鹽度30~32, DO≥5mg/L, pH7.8~ 8.2,光照強度40~100 Lx。
1.2 試驗方法
試驗開始后, 每隔12 h隨機取仔魚15尾, 用尼康(Nikon)E100顯微鏡測量其全長(L)、卵黃囊長徑(R)、卵黃囊短徑(r)、油球直徑(D); 仔魚開口后每隔12 h從試驗組隨機取20尾仔魚放入1 000 mL燒杯中,加入10~15個/mL用小球藻強化的ss輪蟲恒溫飼養(yǎng), 2 h后開始用尼康E100顯微鏡測量其L、R、r、D, 并逐尾解剖檢查其消化道, 記錄其腸管中含有輪蟲的仔魚的尾數(shù)。初次攝食率為腸管中含有輪蟲的仔魚尾數(shù)占所測定總尾數(shù)的百分比。PNR以仔魚的日齡表示, 當(dāng)所測得的饑餓組仔魚的初次攝食率低于最高初次攝食率的一半時, 仔魚的日齡即為PNR。
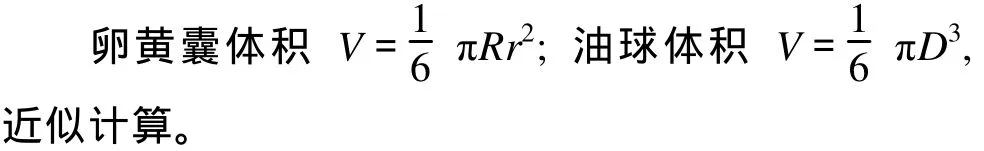
用Excel 2003對數(shù)據(jù)進(jìn)行統(tǒng)計分析、運算和做圖處理。
2 試驗結(jié)果與分析
2.1 仔魚的體長變化和對卵黃囊、油球的吸收
對照組條紋鋸 初孵仔魚平均體長(2.960± 0.21)mm, 通體透明可見部分淡黃色色素, 卵黃囊大而飽滿, 長徑占身體全長的 1/3以上, 體積為 1.27× 10–10m3; 油球較大, 位于卵黃囊的前端, 體積為2.66×10–12m3鰭褶肥厚未分化; 腸管較短, 于身體1/2處, 垂直于身體有一開口。初孵仔魚至開口前的仔魚無攝食能力, 依靠卵黃囊和油球營養(yǎng), 仔魚處于內(nèi)源性營養(yǎng)期, 隨著仔魚的生長發(fā)育, 卵黃囊和油球被迅速吸收(表1)。孵化后1 d時, 仔魚體長進(jìn)一步增加, 卵黃囊體積(1.4×10–11m3)減少(表 1)僅為初孵仔魚的1/9, 油球體積(6.47×10–13m3)為初孵仔魚的1/4(表1)。孵化后2 d時, 仔魚體長進(jìn)一步增加, 仔魚已開口, 但未見攝食, 此時, 卵黃囊體積(1.41×10–12m3)為初孵仔魚卵黃囊體積的 1/90, 油球體積(7.85×10–14m3)為初孵仔魚體積的 1/33; 仔魚開始攝食, 卵黃囊和油球進(jìn)一步消耗, 條紋鋸 仔魚進(jìn)入混合營養(yǎng)期, 此時可見仔魚體長明顯下降(圖1)。孵化3 d之后仔魚體長開始增加, 孵化后3.5 d時仔魚卵黃囊消失, 孵化后4 d時油球消失, 條紋鋸 仔魚進(jìn)入外源營養(yǎng)期。
2.2 初次攝食率和不可逆點的確立
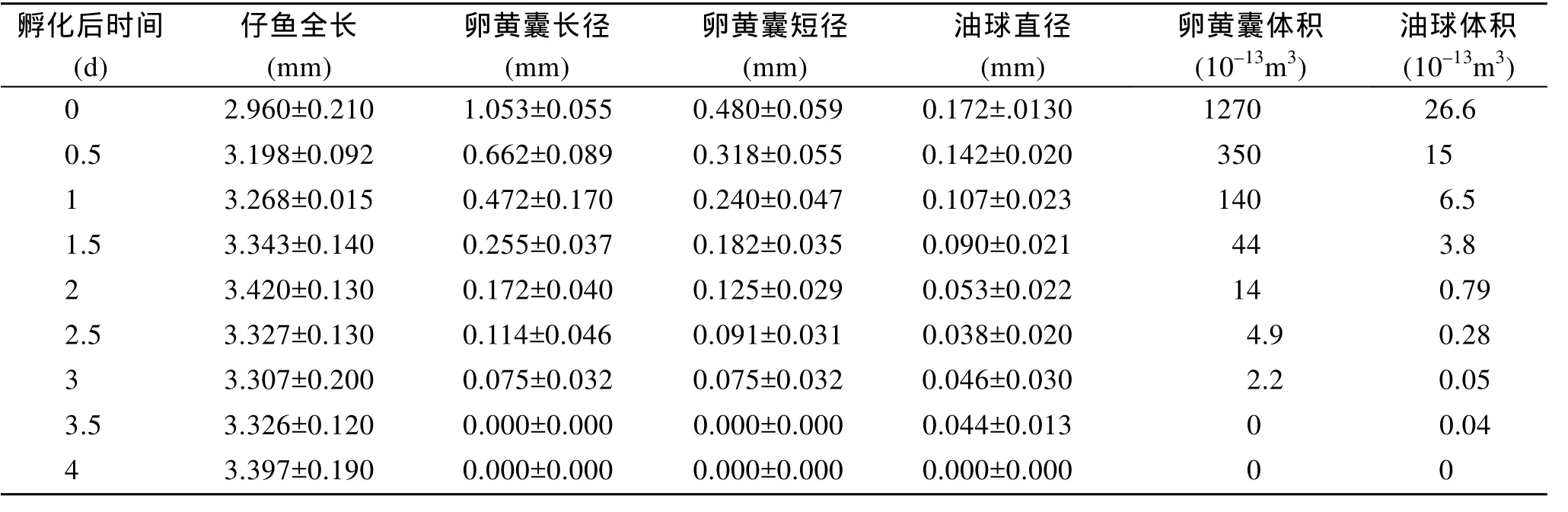
表1 條紋鋸 仔魚全長和卵黃囊、油球的變化Tab.1 Change of total length, size of yolk sac and oil globules of Centropristis striata
條紋鋸 仔魚孵出后第二天即可見口輕微上下開合, 但未見攝食, 孵化后2.5 d時, 可見仔魚攝食,腹部明顯膨大, 腸管進(jìn)一步發(fā)育, 肛門開啟, 仔魚進(jìn)入攝食期。對照組仔魚攝食率為(56.25±3.97)%, 饑餓組仔魚初次攝食率(圖 2)為(68.75±4.47)%, 之后仔魚初次攝食率逐漸增加, 至3.5 d時, 仔魚初次攝食率達(dá)到最大為(88.24±6.38)%, 之后仔魚初次攝食率逐漸下降, 至 5 d時, 仔魚初次攝食率下降為(37.50± 5.45)%, 低于最高初次攝食率的一半, 表明條紋鋸仔魚進(jìn)入不可逆期(PNR期)。5.5 d時, 仔魚初次攝食率僅為(21.43±9.21)%, 6 d時, 仔魚頭部瘦削, 腹部極度萎縮, 已不再攝食, 至6.5 d時, 試驗組(饑餓組)仔魚全部死亡。
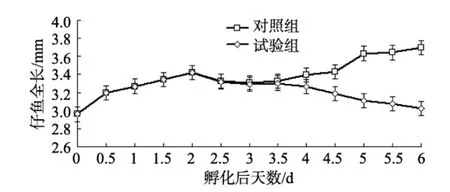
圖1 饑餓仔魚與攝食仔魚全長變化的比較Fig.1 Comparison of total length between the unfed larvae and the fed larvae of Centropristis striata
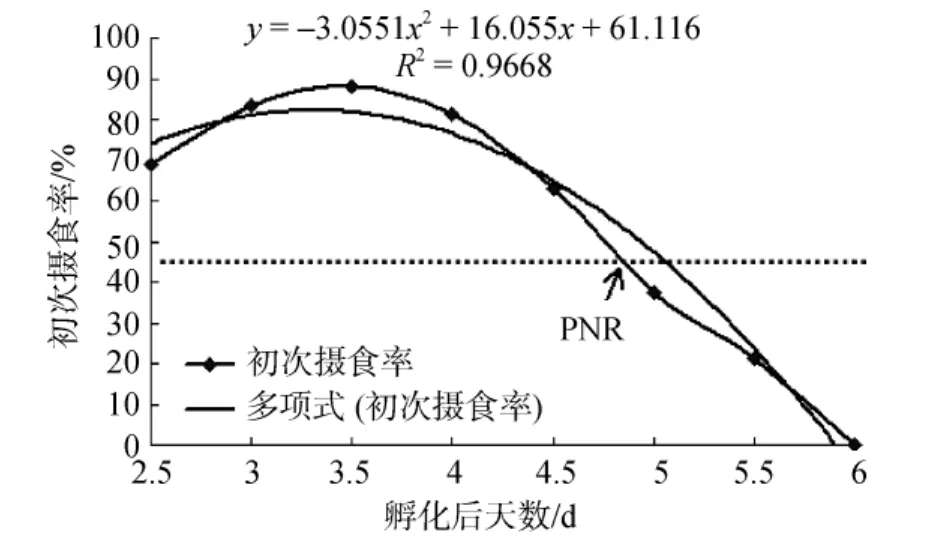
圖2 條紋鋸 初次攝食率的變化曲線Fig.2 Initial feeding rate of Centropristis striata
2.3 攝食強度變化
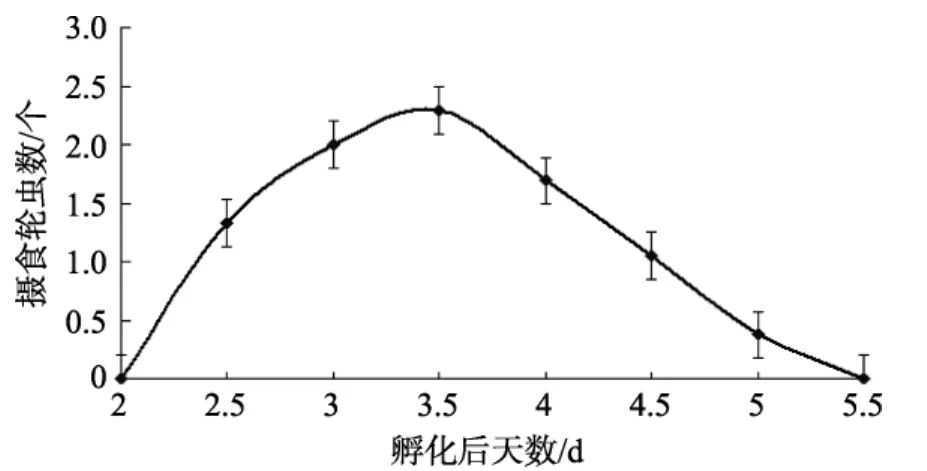
圖3 條紋鋸 攝食強度變化Fig.3 Change of number of rotifers feeding for Centropristis striata
條紋鋸 仔魚開口攝食后, 不同的饑餓時間初次攝食的強度有不同的變化(圖3)。2 d時, 投喂輪蟲2 h后, 解剖發(fā)現(xiàn)仔魚腸道中未見有輪蟲。2.5 d時攝食強度較低, 平均每尾仔魚可攝食輪蟲1.3個/尾, 隨著仔魚饑餓時間的延長, 攝食強度逐漸增加, 至3.5 d時攝食強度達(dá)到最大, 為 2.29個/尾, 解剖可見攝食強度最大的一尾仔魚可攝食 6個輪蟲, 之后攝食強度開始下降, 4 d時, 仔魚平均攝食強度為1.69個/尾, 5 d時, 攝食強度繼續(xù)下降為0.375個/尾, 至5.5 d時,仔魚攝食強度降為0。
3 討論與結(jié)論
3.1 條紋鋸 仔魚不同營養(yǎng)期與PNR
在對條紋鋸 仔魚的培養(yǎng)過程中, 營養(yǎng)是影響仔魚生長、存活的最重要因素之一, 根據(jù)仔魚對內(nèi)源性營養(yǎng)物質(zhì)卵黃囊和油球的吸收利用及仔魚開口攝食的時間, 可以將條紋鋸 對照組仔魚生長分為:內(nèi)源性營養(yǎng)期的快速生長、混合營養(yǎng)期的微弱負(fù)增長、外源性營養(yǎng)期的正常生長3個階段; 試驗組生長分為: 內(nèi)源性營養(yǎng)期的快速生長、混合營養(yǎng)期的微弱負(fù)增長、可建立外源性攝食的負(fù)增長和不可建立外源性攝食的負(fù)增長4個階段。
根據(jù)試驗研究發(fā)現(xiàn), 在(22.9±0.8)℃條件下, 0~2.5 d為內(nèi)源性營養(yǎng)期, 2.5~4 d為混合營養(yǎng)期, 4 d之后為外源性營養(yǎng)期。由表2可知條紋鋸 早期仔魚的混合營養(yǎng)期僅為1.5 d, 仔魚初次攝食到PNR的時間為2.5 d, 真鯛[24]混合營養(yǎng)期為3~4 d, 初次攝食至PNR時間為4~5 d, 黃鯛[25]混合營養(yǎng)期為3 d, 初次攝食至PNR時間為4~5 d, 牙鲆[24]混合營養(yǎng)期為1~2 d, 初次攝食至PNR時間為3~4 d, 點帶石斑魚[26]混合營養(yǎng)期為2 d, 初次攝食至PNR時間為2~3 d, 斜帶石斑魚[27]混合營養(yǎng)期為 2 d, 初次攝食至 PNR時間為 2.5 d, 相比其他海水魚類, 條紋鋸 必須在更短的時間內(nèi)建立外源性營養(yǎng)關(guān)系, 否則只能面對死亡的后果。
3.2 卵黃囊仔魚對內(nèi)源性營養(yǎng)物質(zhì)的利用
卵黃囊仔魚對內(nèi)源性營養(yǎng)的吸收可以分為兩個部分: 對卵黃囊的吸收和對油球的吸收。仔魚對卵黃囊的吸收主要發(fā)生在仔魚孵化后到1 d的時間內(nèi), 可以吸收掉 89%的卵黃物質(zhì), 而在相同時間內(nèi), 卵黃囊仔魚僅吸收了76%的油球物質(zhì), 由表1可見, 卵黃囊仔魚對卵黃囊和油球的快速吸收促進(jìn)了仔魚的快速生長, 隨著卵黃囊和油球的逐漸消耗, 仔魚對內(nèi)源性營養(yǎng)物質(zhì)的吸收逐漸減慢, 仔魚的生長速率也逐漸減慢。分析可知, 卵黃囊仔魚對卵黃物質(zhì)的吸收符合指數(shù)式:y=3276.7e–1.0684x(R2=0.9975), 而對油球的利用符合二項式:y=1.145x2–13.26x+37.714(R2= 0.9872), 可見卵黃囊仔魚對卵黃物質(zhì)和對油球的利用具有差異性, 進(jìn)一步分析發(fā)現(xiàn)油球和卵黃物質(zhì)體積比符合二項式:y=–0.0052x2+0.0431x–0.021(R2= 0.7698), 由圖4分析可知, 卵黃物質(zhì)和油球供應(yīng)仔魚生長過程中, 油球供應(yīng)能量所占的比重先增大后減小, 可以推測: (1)條紋鋸 對營養(yǎng)物質(zhì)的吸收優(yōu)先級為: 卵黃物質(zhì)>油球, 脂肪做為一種重要的儲能物質(zhì), 與蛋白質(zhì)相比, 相同體積的脂肪能提供更多的能量, 因此仔魚優(yōu)先消耗卵黃物質(zhì)而儲存油球以減輕自身的重量和體積而實現(xiàn)節(jié)約能量的目的, 同時延后對油球的消耗可以延長仔魚初次攝食至PNR的時間, 從而有更大的幾率獲取足夠的食物。這與殷名稱[28]所述的仔魚從初次攝食到PNR的時間越長, 建立外源性營養(yǎng)關(guān)系的可能性就越大; 反之則越短相一致;(2)油球提供浮力, 有助于仔魚快速游動以捕獲獵物。
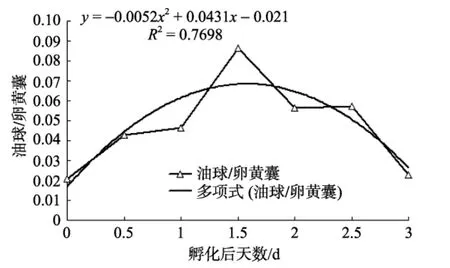
圖4 條紋鋸 油球、卵黃囊體積比的變化Fig.4 Change of volume ratio of yolk sac to oil globule of Centropristis striata

表2 部分海水魚類混合營養(yǎng)期、初次攝食至PNR期時間比較Tab.2 Comparison of mixed nutrition stage time and initial feeding to PNR time
3.3 條紋鋸 仔魚最佳喂食時間
宋兵等[29]認(rèn)為在人工養(yǎng)殖和投喂適口天然餌料的條件下, 饑餓并不是仔魚死亡的主要原因, 過早或過量投喂才是生產(chǎn)中仔魚大量死亡的真正原因。仔魚在剛開口攝食至腸道貫通[30], 包括口與腸道貫通, 腸道與肛門貫通并開口于體外, 需要一定的時間, 若過早投喂會導(dǎo)致食物在胃中無法及時消化吸收導(dǎo)致仔魚的死亡; 若過晚投喂, 仔魚到達(dá) PNR點之后無法及時建立外源性營養(yǎng)期也會導(dǎo)致仔魚的死亡。本研究從攝食生態(tài)學(xué)角度探討了條紋鋸 早期仔魚耐饑餓的特性, 認(rèn)為在(22.9±0.8)℃條件下, 條紋鋸 早期仔魚最佳初次投喂時間為2.5~3.5 d。
[1] 雷霽霖, 盧繼武. 前景廣闊的海水養(yǎng)殖新品種美洲黑石斑魚[J]. 農(nóng)村百事通, 2007, 3: 40-87.
[2] 劉瑞棠. 引進(jìn)美洲黑石斑魚海水網(wǎng)箱養(yǎng)殖試驗[J].河北漁業(yè), 2011, 10: 28-29.
[3] 徐濤, 馬甡, 王志軍. 低溫對條紋鋸 生長及能量收支的影響[J]. 海洋科學(xué), 2007, 31(4): 9-12.
[4] 曹金鳳, 陽剛, 馬甡, 等. 不同溫度條件下條紋鋸耗氧率與窒息點的初步研究[J]. 齊魯漁業(yè), 2009, 27(12): 1-3.
[5] 劉超, 曹金鳳, 馬牲, 等.條紋鋸 對高溫脅迫及脅迫后恢復(fù)方式適應(yīng)的初步研究[J].海洋科學(xué), 2012, 36(9): 54-58.
[6] 王波, 朱明遠(yuǎn), 毛興華.養(yǎng)殖新品種——美洲黑石斑魚[J]. 河北漁業(yè), 2003, 5: 26-27.
[7] 賈瑞錦, 王魯, 趙從明, 等. 條紋鋸 胚胎發(fā)育及卵黃囊仔魚形態(tài)變化的觀察[J]. 漁業(yè)科學(xué)進(jìn)展, 2012, 33(4): 11-17.
[8] 林星.條紋鋸 肌肉營養(yǎng)成分分析與品質(zhì)評價[J]. 養(yǎng)殖與飼料, 2009, 9: 43-46.
[9] 陽鋼, 劉超, 馬甡, 等.條紋鋸 對鹽度脅迫的耐受力及淡水脅迫恢復(fù)的初步研究[J]. 海洋湖沼通報, 2011, 2: 73-78.
[10] 周明濤.引進(jìn)美洲黑石斑魚海水網(wǎng)箱養(yǎng)殖試驗[J]. 現(xiàn)代漁業(yè)信息, 2011, 8: 28-29.
[11] 謝小軍, 鄧?yán)? 張波.饑餓對魚類生理生態(tài)學(xué)影響的研究進(jìn)展[J]. 水生生物學(xué)報, 1998, 22(2): 181-187.
[12] 馬建波, 茅沈麗, 梁桂洪, 等.雜交鱧仔魚饑餓試驗及不可逆點的確定[J]. 水生態(tài)學(xué)雜志, 2012, 3: 90-95.
[13] 黃權(quán), 高峰, 孫曉雨, 等.短期饑餓脅迫對花羔紅點鮭生長率和耗氧率的影響[J]. 吉林農(nóng)業(yè)大學(xué)學(xué)報, 2012, 3: 324-328.
[14] 朱站英, 華雪銘, 于寧, 等.草魚蛋白質(zhì)和脂肪代謝對饑餓脅迫的響應(yīng)[J]. 水產(chǎn)學(xué)報, 2012, 5: 756-763.
[15] 楊明生, 肖漢兵, 曾令兵, 等.溫度對花斑副沙鰍仔魚發(fā)育、攝食及不可逆點的影響[J]. 動物學(xué)雜志, 2012, 4: 114-120.
[16] Blaxter J, Hempel G. The influence of egg size on herring larvae [J]. Cons Int Explor Mer, 1963, 28(3): 211-240.
[17] Dou S, Masuda R, Tanaka M, et al. Feeding resumption, morphological changes and mortality during starvation in Japanese flounder larvae[J]. Fish Biology, 2002, 60(6): 1363-1380.
[18] 吳瑩瑩, 柳學(xué)周, 馬愛軍, 等.饑餓對半滑舌鰨仔魚生長和發(fā)育的影響[J]. 海洋水產(chǎn)研究, 2006, 27(2): 87-93.
[19] 彭志蘭, 柳敏海, 羅海忠, 等.條石鯛仔魚饑餓試驗及不可逆點的確定[J].水產(chǎn)科學(xué), 2010, 3: 152-155.
[20] 宋國, 彭士明, 孫鵬, 等.饑餓與再投喂及投喂頻率對條石鯛幼魚生長和消化酶活力的影響[J].中國水產(chǎn)科學(xué), 2011, 6: 1269-1277.
[21] 施兆鴻, 彭士明, 宋國, 等.饑餓與再投喂對條石鯛幼魚組織和血清中主要代謝酶活性及糖元含量的影響[J].水產(chǎn)學(xué)報, 2012, 9: 1435-1442.
[22] 柳敏海, 施兆鴻, 陳波, 等.饑餓對點帶石斑魚餌料轉(zhuǎn)換期仔魚生長和發(fā)育的影響[J].海洋漁業(yè), 2006, 28(4): 292-298.
[23] 張海發(fā), 劉曉春, 劉付永忠, 等.饑餓對斜帶石斑魚卵黃囊期仔魚攝食、存活及生長的影響[J]. 中山大學(xué)學(xué)報(自然科學(xué)版), 2009, 49(1): 51-55.
[24] 鮑寶龍, 蘇錦祥, 殷名稱.延遲投餌對真鯛、牙鲆仔魚早期階段攝食、存活及生長的影響[J].水產(chǎn)學(xué)報, 1998 , 22(1) : 33 -37.
[25] 夏連軍, 施兆鴻, 陸建學(xué).黃鯛仔魚饑餓試驗及不可逆點的確定[J]. 海洋漁業(yè), 2004, 4: 286-290.
[26] 柳敏海, 施兆鴻, 陳波, 等.饑餓對點帶石斑魚餌料轉(zhuǎn)換期仔魚生長和發(fā)育的影響[J]. 海洋漁業(yè), 2006, 4: 292-298.
[27] 張海發(fā), 劉曉春, 劉付永忠, 等.饑餓對斜帶石斑魚卵黃囊期仔魚攝食、存活及生長的影響[J]. 中山大學(xué)學(xué)報(自然科學(xué)版), 2009, 1: 51-55.
[28] 殷名稱. 北海鯡卵黃囊期仔魚的攝食能力和生長[J].海洋與湖沼, 1991, 6: 554-560.
[29] 宋兵, 陳立僑, 高露姣, 等.延遲投餌對雜交鱘仔魚生長、存活和體成分的影響[J]. 中國水產(chǎn)科學(xué), 2003, 10(3): 222-226.
[30] 駱小年, 李軍, 劉剛, 等.鴨綠江水系唇胚胎發(fā)育、仔魚饑餓及其不可逆點[J].中國水產(chǎn)科學(xué), 2011, 6: 1278-1285.
(本文編輯: 譚雪靜)
Effects of starvation on suriving, feeding and growth of the Yolk-sac larval ofCentropristis striata
JIA Rui-jin1,2, CHEN Chao1,2, KONG Xiang-di1,2, LI Yan-lu1,2, SONG Zhen-xin1,2, WU Lei-ming1,2, WANG Lu1, QU Jiang-bo3, ZHAO Cong-ming4, WU Jian5
(1. Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China; 2. College of Fisheries and Science, Shanghai Ocean University, Shanghai 201306, China; 3. Yantai Tianyuan Aquaculture Co., Ltd, Yantai 264000, China; 4. Tianjin Leadar Marine Resources Development Co, Ltd, Tianjin 300050, China; 5. Zhejiang Haishengyuan Biological Technology Co., Ltd., Taizhou 317600, China)
Jan., 25, 2013
Centropristis striata; larvae; starvation; feeding; the point-of-no-return(PNR)
In order to find the point-of-no-return of black seabass, the yolk-sac larval ofCentropristis striatawas used to explore the variation of suriving, feeding and growth at (22.9±0.8)℃ with salinity 30-32, DO≥5.0 mg/L, pH 7.8~8.2, and light intensity 40~100 Lx. The results showed that, the larval grouper started feeding at the 2nd day past hatching (dph) and the nutrition-mixture stage just lasting one day because the yolk sac and oil globules assimilated down at the 3rd dph. First feeding rate was 68.25% at the 3rd dph and reached the peak of 88.24% at the 3.5th dph. The initial feeding rate reduced to 37.5% which was as low as half of the peak when the larval grouper was 5 days old so the PNR is 5d. All of the fishes died when they were 6.5 days old. The analysis indicates that the endogenous vegetative stage of the grouper had no significant difference between control group and the experimental group. The mixotrophic stage total length had negative growth and a negative growth rate was significantly higher in the experimental group. In exogenous nutrition stage, the total length of experiment group continued to decline until they were died. So we can see, that the growth mode of the yolk sac larvae in control group could be divided into three stages: the rapid growth of endogenous nutrition stage, weak to negative growth of mixed nutrition stage and the normal growth exogenous nutrition stage. The growth mode of the experimental group could be divided into four stages: the rapid growth of endogenous nutrition stage, weak to negative growth of mixed nutrition stage, negative growth stage when the larva could be fed and the negative growth stage when the larva could not be fed. The key period of feeding of yolk-sac larval ofCentropristis striatais in 2nd-5th dph.
S96
A
1000-3096(2014)04-0044-06
10.11759/hykx20130125001
2013-01-25;
2013-09-09
科技部國際合作項目(2012DFA30360); 天津市濱海新區(qū)資助項目(201004070)
賈瑞錦(1988-), 男, 山東臨沂人, 碩士研究生, 主要從事海水魚類苗種繁育與養(yǎng)殖技術(shù)研究, 電話: 15666826817, E-mail: jiaruijin@163.com; 陳超, 通信作者, 電話, 13905328672, E-mail: ysfrichenchao@126.com
猜你喜歡
今日農(nóng)業(yè)(2021年11期)2021-08-13 08:53:34
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
作文周刊·小學(xué)一年級版(2017年17期)2017-06-27 19:32:55
廣東第二課堂·小學(xué)(2017年2期)2017-02-20 15:10:08
