兩種方法在二斑葉螨電壓門控鈉離子通道基因突變檢測中的應(yīng)用
2014-03-26 08:39:04楊順義岳秀利王進軍沈慧敏郭金梅沈一凡周興隆
草業(yè)學(xué)報 2014年5期
關(guān)鍵詞:檢測
楊順義,岳秀利,王進軍,沈慧敏*,郭金梅,沈一凡,周興隆
(1.甘肅農(nóng)業(yè)大學(xué)草業(yè)學(xué)院 草業(yè)生態(tài)系統(tǒng)省部共建教育部重點實驗室 中-美草地畜牧業(yè)可持續(xù)發(fā)展中心,甘肅 蘭州730070;2.西南大學(xué)植物保護學(xué)院 昆蟲學(xué)及害蟲控制工程重點實驗室,重慶400716)
二斑葉螨(Tetranychusurticae)是一種世界性害螨,可危害1100多種寄主植物,除了糧食作物、果樹、花卉、蔬菜等以外,還可以危害城市綠地植物和牧草,且隨著全球變暖,該葉螨在部分寄主植物上的危害有逐漸加重的趨勢[1-3]。二斑葉螨由于體型較小、世代周期短、抗逆性強等特點,抗性發(fā)展迅速[4],對多種農(nóng)藥尤其是甲氰菊酯已產(chǎn)生嚴(yán)重的抗藥性。甲氰菊酯屬擬除蟲菊酯類殺蟲殺螨劑,作用靶標(biāo)為電壓門控鈉離子通道(voltage-gated sodium channels,VGSCs),VGSCs是一種較大的跨膜蛋白,包括4個同源結(jié)構(gòu)域(I-IV),每個結(jié)構(gòu)域由6個跨膜螺旋(S1-S6)組成。VGSCs的不敏感性主要由跨膜螺旋或連接處的點突變引起,主要發(fā)生在ⅡS4-ⅡS6和ⅢS6上[5],并且有的突變與擊倒抗性和超擊倒抗性有著密切聯(lián)系。至今,在蛛形綱蜱螨類以及其他昆蟲抗擬除蟲菊酯種群靶標(biāo)基因VGSCs的點突變已有很多報道。在二斑葉螨抗性種群VGSCs基因中只發(fā)現(xiàn)3個突變,即L1022V、A1215D和F1538I,并且L1022V和F1538I突變對擊倒抗性起著主要作用[6-7]。在朱砂葉螨(T.cinnabarinus)抗甲氰菊酯種群中同樣發(fā)現(xiàn)存在F1538I突變[8];在蜂螨(Varroadestructor)中,VGSCs上也有4個突變F758L,L826P,I982V和M1055I[9],而電生理的研究表明L826P突變對氯氟胺氰戊菊酯敏感性降低起著重要的功能作用[10]。在家蠅(Muscadomestica)的VGSCs中,最常見的L1014F/H/S突變在擊倒抗性和超擊倒抗性種群中都存在,而M918T突變只存在于超擊倒抗性種群中[11-13]。靶標(biāo)VGSCs基因突變是害螨及昆蟲對擬除蟲菊酯類藥劑產(chǎn)生抗性的主要機制。
目前,基因突變檢測的方法很多,主要有焦磷酸直接測序(direct pyrosequencing,DS),單鏈構(gòu)象多態(tài)性分析(single stranded conformation polymorphism,SSCP),限制性片段長度多態(tài)性聚合酶鏈反應(yīng)(polymerase chain reaction-restriction fragment length polymorphism,PCR-RFLP)以及實時熒光定量PCR(real-time quantitive PCR,RT-qPCR)和毛細(xì)管電泳法(capillary electrophoresis,CE)[14]等。高明等[15]通過克隆測序發(fā)現(xiàn)甜菜夜蛾(Spodopteraexigua)抗性種群鈉離子通道基因ⅡS6上存在L1014F突變。各種檢測方法優(yōu)缺點不一,有必要尋找一種經(jīng)濟、靈敏且有效的突變檢測方法。本研究采用PCR產(chǎn)物直接測序法與PCR-RFLP兩種不同的方法對田間采集的二斑葉螨種群VGSCs基因進行突變位點的檢測,旨在建立二斑葉螨對菊酯類農(nóng)藥抗性的分子檢測方法,為二斑葉螨的抗藥性治理提供技術(shù)支持。
1 材料與方法
1.1 供試二斑葉螨
二斑葉螨相對敏感種群(susceptible strain,SS):2007年6月底采自甘肅省蘭州市興隆山國家級森林公園金花忍冬(Lonicerachrysantha)上。在室內(nèi)采用葉碟法(培養(yǎng)皿底部墊有浸水和表面鋪一黑布的海綿,黑布上放用脫脂棉包圍的豇豆葉片)進行單對(挑一雌一雄在葉碟上飼養(yǎng),不接觸任何藥劑)繁殖,待種群擴大后轉(zhuǎn)移到盆栽豇豆(Vignaunguiculata)苗上擴繁。田間抗性種群:LZ-R和WW-R種群于2012年8月分別采集于甘肅省蘭州市和武威市田間種植的茄子(Solanummelongena)和南瓜(Cucurbitamoschata)上,待種群在室內(nèi)擴繁定殖后,測定其對甲氰菊酯的抗性分別達(dá)到27.4和23.1倍。以上二斑葉螨SS、WW-R和LZ-R種群飼養(yǎng)在養(yǎng)蟲室盆栽的豇豆苗上。控制條件:溫度(26±1)℃,相對濕度(75±5)%,光照14h,黑暗10h。
1.2 主要藥劑及儀器
20%甲氰菊酯乳油(紅太陽農(nóng)藥集團有限公司),RNA提取試劑盒RNeasy?Plus Micro Kit(Qiagen公司),反轉(zhuǎn)錄試劑盒PrimerScript?RT Reagent Kit,rTaq酶等PCR相關(guān)試劑(TaKaRa公司),限制性核酸內(nèi)切酶Sau3AI和BclI(Thermo公司),S1000TMThermal Cycling PCR儀(Bio-Rad公司),PowerPac Basic電泳儀(Bio-Rad公司),Universal Hood II凝膠成像儀(Bio-Rad公司),Nanovue核酸濃度測定儀(GE Healthcare公司)。
1.3 二斑葉螨總RNA提取
于2012年9月分別挑取SS、WW-R和LZ-R種群的二斑葉螨雌成螨約500頭,按照Qiagen公司RNA提取試劑盒RNeasy?Plus Micro Kit使用說明書提取總RNA,具體操作步驟如下:
1)預(yù)先將研缽用液氮進行多次冷凍,之后加入待研磨樣品,反復(fù)研磨,再加入350μL Buffer RLT plus繼續(xù)研磨均勻,轉(zhuǎn)移至2mL離心管;
2)將離心管置離心機中13000r/min離心3min,吸取上清液,記住大致體積V,轉(zhuǎn)移到gDNA Eliminator spin column離心管,10000r/min離心30s,丟掉柱子保存液體;
3)在上述液體中加入V體積的70%酒精,混勻;
4)轉(zhuǎn)移上述液體至RNeasy MinElute spin column離心管中,蓋嚴(yán)蓋子,10000r/min離心15s;
5)在RNeasy MinElute spin column離心管中加入700μL Buffer W1,蓋嚴(yán)蓋子,10000r/min離心15s;
6)在RNeasy MinElute spin column離心管中加入500μL Buffer RPE,蓋嚴(yán)蓋子,10000r/min離心15s;
7)在RNeasy MinElute spin column離心管中加入500μL的80%酒精,蓋嚴(yán)蓋子,10000r/min離心2min;
8)空離。將柱子放入一新的2mL離心管里,開蓋,13000r/min離心5min。
9)洗脫RNA。將柱子放入一新的1.5mL的收集管中。在柱子膜中央加入20μL RNase-free water,輕輕蓋上蓋子,13000r/min離心1min,棄柱子,得到的即所需RNA。
對提取得到的總RNA經(jīng)Nanovue核酸濃度測定儀測定其濃度時,依照A260/280和A260/230的值確定RNA純度;RNA純度確定后進行瓊脂糖電泳膠電泳,根據(jù)條帶的數(shù)目及亮度檢測RNA的完整性。最終,選取A260/280在1.8~2.2之間,A260/230在2.0~2.4之間的RNA樣品保存于-80℃冰箱,備用。
1.4 反轉(zhuǎn)錄合成第一鏈cDNA
將提取得到的二斑葉螨SS、WW-R和LZ-R總RNA分別反轉(zhuǎn)錄合成第一鏈cDNA,方法參照Takara公司反轉(zhuǎn)錄試劑盒PrimerScript?RT Reagent Kit Perfect Real time試劑盒說明書進行。反應(yīng)體系10μL:5×Prime-Script?Buffer(for Real Time),2μL;PrimeScript?RT Enzyme Mix I,0.5μL;Oligo dT Primer(50μmol/L),0.5μL;Random 6mers(100μmol/L),0.5μL;總RNA不超過0.5μg;最后補充RNase Free dH2O至10μL。待所有樣品加完后,將其放入PCR儀中37℃反轉(zhuǎn)錄反應(yīng)15min,85℃反轉(zhuǎn)錄酶失活反應(yīng)5s。反應(yīng)完畢后,測定cDNA濃度,將反轉(zhuǎn)錄產(chǎn)物保存于-20℃冰箱,備用。
1.5 引物設(shè)計
根據(jù)二斑葉螨在線基因組數(shù)據(jù)庫(http://bioinformatics.psb.ugent.be/webtools/bogas/overview/Tetur)中VGSCs基因(基因登錄號:Tetur34g00970)全長序列以及該基因序列上報道存在的F1022V、A1215D和F1538I突變等信息,用Primer 5.0軟件分別設(shè)計用于靶標(biāo)位點VGSCs上F1022、A1215和F1538基因片段擴增的特異性引物以及PCR-RFLP反應(yīng)的引物,引物名稱、片段長度及用途見表1。

表1 用于PCR擴增的特異性引物及其用途Table 1 Specific and quantitative primers and their use in the experiment
1.6 PCR反應(yīng)及產(chǎn)物的測序
用于擴增二斑葉螨鈉通道相關(guān)突變基因的PCR反應(yīng)體系為25μL,其中10×PCR buffer 2.5μL,MgCl2(25 mmol/L)2.5μL,dNTPs(2.5mmol/L)2μL,ddH2O 14.8μL,rTaq酶(5U/μL)0.2μL,上下游引物(10 μmol/L)各1μL,cDNA模板(二斑葉螨不同種群的cDNA)1μL。擴增條件:95℃預(yù)變性3min;95℃變性30s,55℃復(fù)性30s,72℃延伸1min,共35個循環(huán);最后72℃延伸10min。PCR反應(yīng)產(chǎn)物經(jīng)1%瓊脂糖凝膠電泳,若檢測到目的條帶,且條帶單一、明亮,則可以直接進行測序。試驗重復(fù)3次。
1.7 突變位點的PCR-RFLP法檢測
A1215D突變位點的檢測:采用PCR-RFLP法擴增目的片段,擴增產(chǎn)物用限制性核酸內(nèi)切酶消化成不同大小的片段,直接在瓊脂糖凝膠電泳上分辨。應(yīng)用Primer 5.0軟件進行酶切位點的尋找,不同種群的PCR擴增產(chǎn)物經(jīng)內(nèi)切酶Sau3AI酶切處理,識別位點為↓GATC。WW-R和LZ-R種群若發(fā)生GCT→GAT突變,就會產(chǎn)生一個酶切位點↓GATC,能被Sau3AI酶切消化產(chǎn)生498和99bp兩條帶,而SS種群由于是正常堿基,不存在相應(yīng)的識別位點,不能被內(nèi)切酶消化,只有597bp的條帶。酶切后產(chǎn)物在2%的瓊脂糖凝膠上電泳,根據(jù)片段的位置、大小即可鑒定出A1215D突變存在與否。試驗重復(fù)3次。
F1538I突變的檢測:由于F1538I突變位點附近沒有合適的酶切位點,根據(jù)創(chuàng)造酶切位點原理[16-17],將引物3′端最后一個堿基錯配引入一個酶切位點T↓GATCA,即將突變堿基前一個堿基T人為的更改為G,并在此處設(shè)計引物(表1)。WW-R和LZ-R種群若發(fā)生TTC→ATC突變,就會產(chǎn)生一個酶切位點T↓GATCA,能被BclI酶切消化產(chǎn)生277和20bp兩條帶,而SS只有297bp一條帶,酶切后在2%的瓊脂糖凝膠上電泳,根據(jù)片段的大小、位置即可鑒定出F1538I突變的存在與否。試驗重復(fù)3次。
2 結(jié)果與分析
2.1 二斑葉螨VGSCs基因片段擴增及測序
2.1.1 突變位點區(qū)域片段擴增 二斑葉螨SS、WW-R和LZ-R種群VGSCs基因L1022、A1215和F1538位點片段擴增序列經(jīng)瓊脂糖凝膠電泳如圖1,3種群擴增產(chǎn)物條帶單一、明亮,均為目的片段,符合PCR產(chǎn)物直接測序的基本要求。
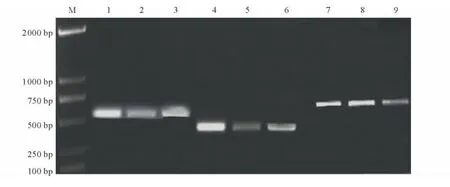
圖1 二斑葉螨不同種群突變位點擴增電泳圖Fig.1 Electrophoretogram of mutation sites in different strains of T.urticae
2.1.2 VGSCs基因PCR產(chǎn)物測序結(jié)果 根據(jù)二斑葉螨SS、WW-R和LZ-R種群A1215和F1538位點部分基因片段測序結(jié)果表明,與二斑葉螨SS種群VGSCs基因核苷酸相比對,WW-R和LZ-R種群均存在點突變,第一個突變位點在結(jié)構(gòu)域ⅡS6和ⅢS1連接處,GCT(丙氨酸)被GAT(天冬氨酸)取代,即存在A1215D突變;第二個突變發(fā)生在結(jié)構(gòu)域ⅢS6處,TTC(苯丙氨酸)被ATC(異亮氨酸)取代,即存在F1538I突變,其中 WW-R種群中F1538I為純合突變,而LZ-R中為雜合突變。然而,L1022V突變在二斑葉螨WW-R和LZ-R種群均未檢測到。
2.2 二斑葉螨VGSCs基因突變PCR-RFLP檢測
2.2.1 A1215D突變位點的PCR-RFLP檢測 采用PCR-RFLP的方法進行酶切鑒定二斑葉螨 WW-R和LZ-R種群A1215D突變。通過Sau3AI酶切處理SS、WW-R和LZ-R種群后,發(fā)現(xiàn) WW-R和LZ-R種群均能被內(nèi)切酶切開,產(chǎn)生498和99bp兩條帶,而SS種群只有一個597bp的條帶(圖2)。酶切前后根據(jù)條帶的位置、數(shù)目可以明顯的區(qū)分突變種群,但由于99bp的條帶太暗凝膠成像顯示不清晰,而498和597bp兩條帶區(qū)分明顯,并不影響突變位點的鑒定,說明二斑葉螨WW-R和LZ-R種群中都存在A1215D氨基酸突變,而根據(jù)酶切結(jié)果電泳圖沒有發(fā)現(xiàn)WW-R和LZ-R種群中存在雜合現(xiàn)象。
2.2.2 F1538I突變位點的檢測 采用錯配堿基引入酶切位點改進的PCR-RFLP方法進行F1538I的突變檢測,結(jié)果表明酶切前后WW-R和LZ-R種群條帶大小、位置沒有發(fā)生變化,與SS種群沒有明顯差異(圖3),即沒有被BclI內(nèi)切酶消化。為了排除是由于酶切前后電泳條帶在2%濃度瓊脂糖凝膠電泳條件下不易區(qū)分引起的,通過增加瓊脂糖凝膠電泳的濃度試驗結(jié)果相同;最后通過PCR產(chǎn)物測序發(fā)現(xiàn),錯配堿基的引入會導(dǎo)致下游片段引物延伸時突變堿基處產(chǎn)生差異,從而不能形成BclI識別的酶切位點,即PCR-RFLP法不能有效地檢測出WW-R和LZ-R種群的F1538I突變。
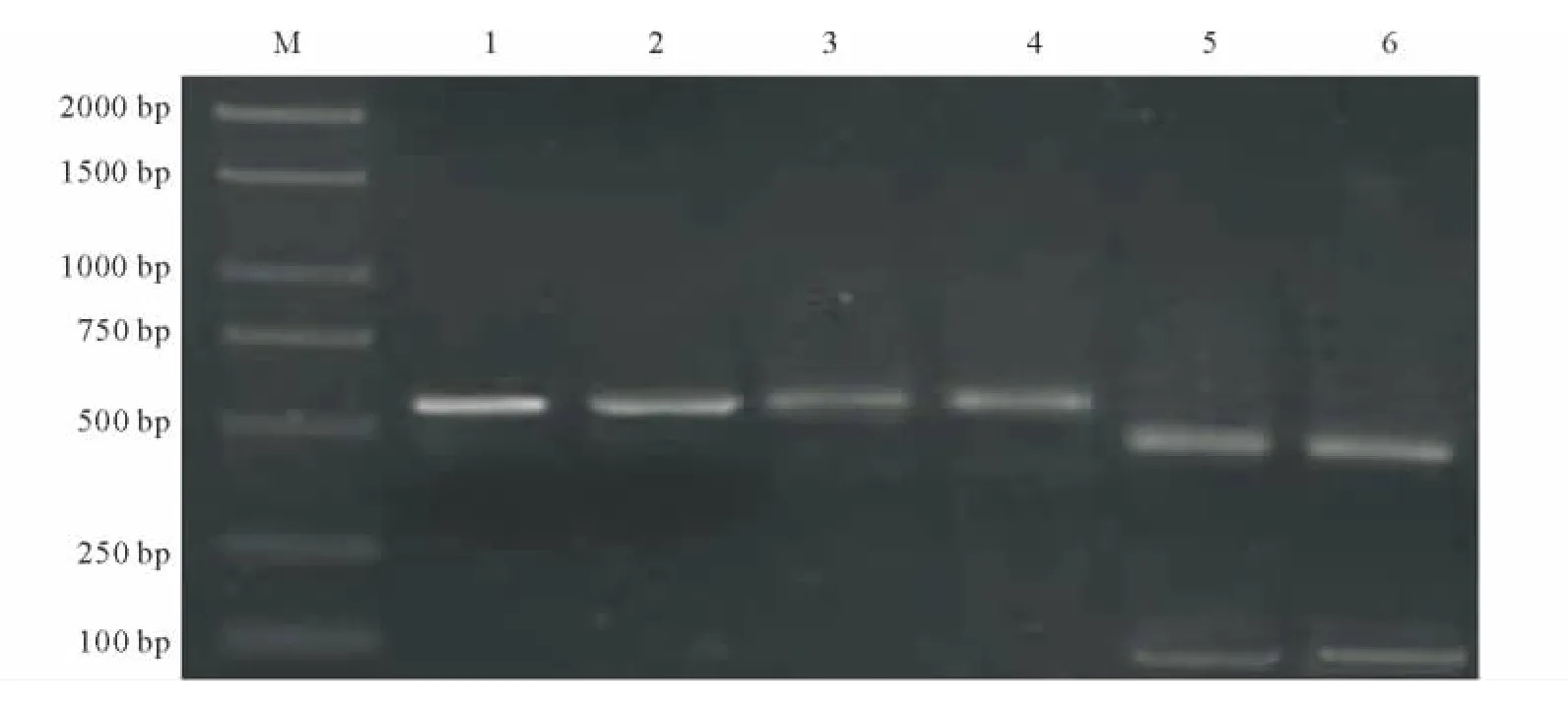
圖2 二斑葉螨不同種群A1215D突變酶切電泳圖Fig.2 Electrophoretogram results of the A1215Dmutation in different T.urticae strains with enzyme digestion

圖3 二斑葉螨不同種群F1538I突變酶切電泳圖Fig.3 Electrophoretogram results of the F1538Imutation in different T.urticae strains with enzyme digestion
3 討論
目前,在二斑葉螨抗性種群VGSCs基因上發(fā)現(xiàn)存在L1022V、A1215D和F1538I共3個氨基酸突變。通過功能分析表明L1022V和F1538I突變在二斑葉螨對擬除蟲菊酯類農(nóng)藥的不敏感性起重要作用[6,18],并且F1538I突變被認(rèn)為是對擬除蟲菊酯具有最高效的抗性位點之一[19-20],是家蠅鈉通道ⅢS6螺旋處的芳香族氨基酸殘基,同時也是擬除蟲菊酯的疏水性結(jié)合位點[21-22];而A1215D突變的功能尚不明確,還需要進一步的研究。本研究發(fā)現(xiàn),田間采集的二斑葉螨WW-R和LZ-R種群對甲氰菊酯抗藥性的產(chǎn)生與F1538I突變有關(guān),F(xiàn)1538I突變是二斑葉螨對甲氰菊酯產(chǎn)生高抗性的重要原因;這與前期本實驗室研究室內(nèi)飼養(yǎng)的二斑葉螨經(jīng)甲氰菊酯抗性培育45代后,同樣會產(chǎn)生F1538I突變的結(jié)果一致[23]。說明田間和室內(nèi)的二斑葉螨在甲氰菊酯藥劑的脅迫作用下,其靶標(biāo)VGSCs基因均會產(chǎn)生同樣的突變,從而對其高抗性的產(chǎn)生起著重要作用。隨著二斑葉螨全基因組的發(fā)布,突變的檢測將更有利于對其抗性分子機理以及抗藥性檢測的研究[24],其中與代謝相關(guān)的二斑葉螨水平基因轉(zhuǎn)移[25]以及二斑葉螨對寄主植物的適應(yīng)性與抗藥性的關(guān)系[26]最近已有報道。因此,從分子水平上檢測擬除蟲菊酯類農(nóng)藥靶標(biāo)VGSCs突變,對相關(guān)的抗性機理研究具有重要意義。
通過比較兩種突變檢測方法,PCR-RFLP法雖然較快速方便,不需要測序,但是由于酶切位點以及內(nèi)切酶價格的限制,并且只能在已知突變的情況下才能采用,這在一定程度上限制了該技術(shù)的使用,并且錯配堿基創(chuàng)造酶切位點的方法在堿基錯配后,有時可能會導(dǎo)致下游1~2個堿基不能正常擴增配對,降低了酶切鑒定的敏感度,本研究采用此方法則不能有效地檢測出F1538I突變。相對于PCR-RFLP法,通過PCR產(chǎn)物正反雙向直接測序,可以檢測出二斑葉螨突變位點A1215D和F1538I,并能明確突變堿基類型,重復(fù)性更好,結(jié)果更準(zhǔn)確,但操作相對繁瑣,測序較耗時;而其他方法如高通量融解曲線法[27]等由于儀器昂貴,成本較高,也不適合一般實驗室采用。熊亮等[28]通過比較PCR產(chǎn)物直接測序法和高通量溶解曲線分析技術(shù)檢測膠質(zhì)瘤表皮生長因子受體(epidermal growth factor receptor,EGFR)基因突變類型發(fā)現(xiàn),這兩種方法在檢測EGFR外顯子19突變率具有較高的一致性。因此,伴隨著新一代高通量分子測序技術(shù)的不斷進步和發(fā)展,測序周期將大大縮短,PCR產(chǎn)物直接測序法將更有利于害螨及昆蟲抗藥性的突變檢測。
[1] Grbic M,Van Leeuwen T,Clark R M,etal.The genome ofTetranychusurticaereveals herbivorous pest adaptations[J].Nature,2011,479:487-492.
[2] 張廷偉,沈慧敏,錢秀娟,等.二斑葉螨刺吸脅迫對白三葉葉綠素含量和兩種保護酶的影響[J].應(yīng)用昆蟲學(xué)報,2013,50(2):395-400.
[3] 賀達(dá)漢,趙曉萍,靳巧紅,等.寧夏地區(qū)二斑葉螨的寄主植物選擇及其季節(jié)轉(zhuǎn)移[J].環(huán)境與應(yīng)用生物學(xué)報,2001,7(5):447-451.
[4] Croft B A,Vandebaan H E.Ecological and genetic-factors influencing evolution of pesticide resistance in tetranychid and phytoseiid mites[J].Experimental and Applied Acarology,1988,4(3):277-300.
[5] Soderlund D M.Pyrethroids,knockdown resistance and sodium channels[J].Pest Management Science,2008,64(6):610-616.
[6] Tsagkarakou A,Van Leeuwen T,Khajehali J,etal.Identification of pyrethroid resistance associated mutations in the para sodium channel of the two-spotted spider miteTetranychusurticae(Acari:Tetranychidae)[J].Insect Molecular Biology,2009,18(5):583-593.
[7] Kwon D H,Clark J M,Lee S H.Cloning of a sodium channel gene and identification of mutations putatively associated with fenpropathrin resistance inTetranychusurticae[J].Pesticide Biochemistry and Physiology,2010,97(2):93-100.
[8] Feng Y N,Zhao S,Sun W,etal.The sodium channel gene inTetranychuscinnabarinus(Boisduval):identification and expression analysis of a mutation associated with pyrethroid resistance[J].Pest Management Science,2011,67(8):904-912.
[9] Wang R W,Liu Z Q,Dong K,etal.Association of novel mutations in a sodium channel gene with fluvalinate resistance in the mite,Varroadestructor[J].Journal of Apicultural Research,2002,41(1-2):17-25.
[10] Liu Z,Tan J,Huang Z Y,etal.Effect of a fluvalinate-resistance-associated sodium channel mutation from varroa mites on cockroach sodium channel sensitivity to fluvalinate,apyrethroid insecticide[J].Insect Biochemistry and Molecular Biology,2006,36(11):885-889.
[11] Ingles P J,Adams P M,Knipple D C,etal.Characterization of voltage-sensitive sodium channel gene coding sequences from insecticide-susceptible and knockdown-resistant house fly strains[J].Insect Biochemistry and Molecular Biology,1996,26(4):319-326.
[12] Williamson M S,Martinez-Torres D,Hick C A,etal.Identification of mutations in the housefly para-type sodium channel gene associated with knockdown resistance(kdr)to pyrethroid insecticides[J].Molecular & General Genetics,1996,252(1-2):51-60.
[13] Rinkevich F D,Leichter C A,Lazo T A,etal.Variable fitness costs for pyrethroid resistance alleles in the house fly,Muscadomestica,in the absence of insecticide pressure[J].Pesticide Biochemistry and Physiology,2013,105:161-168.
[14] 曹曉梅,趙彤言.害蟲抗藥性分子檢測技術(shù)發(fā)展現(xiàn)狀[J].寄生蟲與醫(yī)學(xué)昆蟲學(xué)報,2006,13(1):57-63.
[15] 高明,申瑞平,張鵬,等.競爭性特異等位基因PCR檢測山東甜菜夜蛾對高效氯氰菊酯的抗性[J].植物保護學(xué)報,2013,40(4):363-368.
[16] 趙春江,李寧,鄧學(xué)梅.應(yīng)用創(chuàng)造酶切位點法檢測單堿基突變[J].遺傳,2003,25(3):327-329.
[17] 廖相云,張雅芬,顧學(xué)范.擴增引進限制性酶切位點技術(shù)對兩種CYP21基因突變的檢測[J].中華醫(yī)學(xué)遺傳學(xué)雜志,2003,20(5):449-451.
[18] Van Leeuwen T,Vontas J,Tsagkarakou A,etal.Acaricide resistance mechanisms in the two-spotted spider miteTetranychusurticaeand other important Acari:A review[J].Insect Biochemistry and Molecular Biology,2010,40(8):563-572.
[19] He H,Chen A C,Davey R B,etal.Identification of a point mutation in the para-type sodium channel gene from a pyrethroid-resistant cattle tick[J].Biochemical and Biophysical Research Communications,1999,261(3):558-561.
[20] Tan J G,Liu Z Q,Wang R W,etal.Identification of amino acid residues in the insect sodium channel critical for pyrethroid binding[J].Molecular Pharmacology,2005,67(2):513-522.
[21] O’Reilly A O,Khambay B P,Williamson M S,etal.Modelling insecticide-binding sites in the voltage-gated sodium channel[J].Biochemical Journal,2006,396(2):255-263.
[22] Davies T G E,O’Reilly A O,F(xiàn)ield L M,etal.Knockdown resistance to DDT and pyrethroids:from target-site mutations to molecular modelling[J].Pest Management Science,2008,64(11):1126-1130.
[23] 高新菊.二斑葉螨對甲氰菊酯的抗性分子機理研究[D].蘭州:甘肅農(nóng)業(yè)大學(xué),2012.
[24] Van Leeuwen T,Dermauw W,Grbic M,etal.Spider mite control and resistance management:does a genome help?[J]Pest Management Science,2013,69(2):156-159.
[25] Wybouw N,Balabanidou V,Ballhorn D J,etal.A horizontally transferred cyanase gene in the spider miteTetranychusurticaeis involved in cyanate metabolism and is differentially expressed upon host plant change[J].Insect Biochemistry and Molecular Biology,2012,42(12):881-889.
[26] Dermauw W,Wybouw N,Rombauts S,etal.A link between host plant adaptation and pesticide resistance in the polyphagous spider miteTetranychusurticae[J].Proceedings of the National Academy of Sciences of the United States of America,2013,110(2):113-122.
[27] Whittall R A,Scartezini M,Li K,etal.Development of a high-resolution melting method for mutation detection in familial hyperch-olesterolaemia patients[J].Annals of Clinical Biochemistry,2010,47:44-55.
[28] 熊亮,梁朝峰,陳川,等.qPCR-HRM曲線分析技術(shù)與普通PCR加直接測序法檢測膠質(zhì)瘤EGFR突變比較[J].新醫(yī)學(xué),2012,43(5):327-330.
猜你喜歡
中國設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
