醋酸菌的分類進展
2014-04-12 06:09:14陳福生
中國釀造 2014年12期
關(guān)鍵詞:分類
王 斌,陳福生*
(華中農(nóng)業(yè)大學(xué)食品科技學(xué)院,食品生物技術(shù)與安全實驗室,湖北武漢 430070)
醋酸菌(acetic acid bacteria,AAB)是指能以氧氣為終端電子受體,氧化糖類、糖醇類和醇類生成相應(yīng)的糖醇、酮和有機酸的革蘭氏陰性細菌的總稱[1]。第一個AAB屬——醋酸桿菌屬(Acetobacter,A.),是1898年由BEIJERINCK M[2]提出的。隨后,葡糖桿菌屬(Gluconobacter,G.)和酸單胞菌屬(Acidomonas,Ac.)分別在1935年和1989年由ASAI和UHLIG提出[3]。從1898第一個AAB屬被發(fā)現(xiàn)至1989年的91年中,僅報道了2個新AAB屬,不到10個種。而從1989年至今的25年中,AAB的研究突飛猛進,截至2014年初,已報道的AAB共16個屬,84個種。隨著AAB種類的增加,AAB的功能也逐漸被發(fā)掘。AAB最初因具有生產(chǎn)醋酸的能力而得名,隨后的研究發(fā)現(xiàn),有些AAB并不具有產(chǎn)醋酸能力,如朝井桿菌屬(Asaia,As.)和糖桿菌屬(Saccharibacter,S.)[4-5];而有些AAB除生產(chǎn)醋酸外,還具有產(chǎn)生纖維素、色素、吲哚乙酸、抗壞血酸和固氮等功能[6-10]。本文總結(jié)了AAB常用的分離培養(yǎng)基、AAB分類、鑒定與保藏方法、AAB命名原則與種屬名稱演變,并對AAB的種屬特征進行了歸納。
1 AAB的分離、分類與保藏
1.1 分離AAB常用的培養(yǎng)基
AAB廣泛分布于發(fā)酵食品(如食醋、酒類產(chǎn)品、可可豆和紅茶)、植物器官(如水果、花朵、花粉、莖和根)、動物器官(如果蠅腸道和人體淋巴結(jié))和土壤(如植物根系周圍土壤)、釀造廠周邊土壤和古墓墻壁中。樣品不同,分離AAB的培養(yǎng)基種類不同(見表1),分離得到的AAB特性也不同。
分離AAB的常用培養(yǎng)基主要包括葡萄糖酵母浸膏蛋白胨乙醇(glucose yeast peptone ethanol,GYPE)培養(yǎng)基和葡萄糖酵母浸膏(glucose yeast,GY)培養(yǎng)基。采用這些培養(yǎng)基分離AAB時,通常還需要采用含有100 μg/kg環(huán)己酰亞胺等真菌抑制劑,pH值為3.5的葡萄糖酵母浸膏蛋白胨醋酸乙醇(glucose yeast peptone acetic acid-ethanol,GYPAE)富集培養(yǎng)基進行富集,然后再進行分離;分離具有固氮功能的AAB時,一般選用Dobereiner無氮培養(yǎng)基(又名LGI培養(yǎng)基);而分離高耐酸(醇)的AAB時,常常采用含有一定濃度醋酸和酒精的強化醋酸乙醇(reinforced acetic acidethanol,RAE)培養(yǎng)基和醋酸乙醇(acetic acid-ethanol,AE)培養(yǎng)基。此外,也有學(xué)者采用雙層培養(yǎng)基分離AAB。如ENTANI E等[11]以含0.5%瓊脂的AE作為底層培養(yǎng)基,用1%瓊脂的AE培養(yǎng)基作為上層培養(yǎng)基,并將分離平板置于水分活度達95%~100%的密閉容器中培養(yǎng),成功分離得到了產(chǎn)酸量達10%~15%的AAB。表1列出了分離AAB的主要培養(yǎng)基的名稱、組成成分和適用范圍。

表1 分離醋酸菌的主要培養(yǎng)基及其適用范圍Table 1 Medium to isolate AAB and their applicable scope
1.2 AAB的分類、鑒定方法
關(guān)于AAB的分類與鑒定,早期主要基于其菌落(菌體)形態(tài)與碳(氮)源的同化特征以及輔酶Q的類型和脂肪酸組成等特征進行[3]。隨著分子生物學(xué)技術(shù)的發(fā)展,常常將這些特征與AAB的遺傳特征相結(jié)合來進行分類與鑒定。自從1980年GILLIS M等[15]第一次將DNA堿基組成[16]用于AAB的分類之后,16S rRNA限制性片段長度多態(tài)性分析、16S-23S rDNA ITS[17]、PCR-DGGE[18]、16S-23S-5S rDNA PCR-RFLP[19]、(GTG)5-PCR[20]和Taqman-MGB探針[21]等分子生物學(xué)技術(shù)與手段被先后作為AAB分類鑒定的輔助手段。目前,通常是將上述三方面信息相結(jié)合,對AAB進行分類。
1.3 AAB的保藏
AAB菌種的保藏方法與一般微生物的保藏方法相同。但是研究表明,部分AAB,特別是高產(chǎn)酸AAB的遺傳特征非常不穩(wěn)定,所以這類AAB最好的保藏方法是冷凍干燥保藏,且盡可能減少傳代次數(shù)[22]。高產(chǎn)酸AAB遺傳穩(wěn)定性差的原因可能與AAB基因組中的質(zhì)粒、高可變的重復(fù)序列(hyper mutable tandem repeats)以及轉(zhuǎn)座子(transposons)含量較多相關(guān)[23]。
目前,國際上主要的微生物菌種保藏中心均保藏有AAB,并對外提供菌種保藏、交換與銷售服務(wù)。其中,日本微生物保藏中心(Japan Collectionof Microorganisms,JCM)、日本技術(shù)評價研究所生物資源中心(NITE Biological Re source Center,NBRC)、德國菌種保藏中心(Deutsche Samm lung von Mikroorganismen und Zellkulturen,DSMZ)、比利時微生物綜合保藏中心(Belgian Co-ordinated Collections of Microorganisms,BCCM)、美國典型菌種保藏中心(American Type Culture Collection,ATCC)以及國內(nèi)的工業(yè)微生物菌種保藏中心(Center of Industrial Culture Collection,CICC)和普通微生物菌種保藏中心(General Microbiological Culture Collection Center,CGMCC)等是AAB的主要保藏中心。在這些菌種保藏中心中,JCM、NBRC和DSMZ保藏的AAB的種類與數(shù)量最全面和最多,它們保藏的AAB幾乎覆蓋了目前所有的AAB種屬,保藏的AAB數(shù)量也超過世界AAB菌株總數(shù)量的一半。而我國的上述兩個菌種保藏中心僅保藏4個AAB屬:醋酸桿菌屬(Acetobacter)、葡糖桿菌屬(Gluconobacter)、葡糖醋桿菌屬(Gluconacetobacter)和朝井桿菌屬(Asaia),13種AAB,共137株。
2 AAB的種屬
AAB的分類發(fā)展大致分為兩個階段。第一階段是AAB的發(fā)現(xiàn)與認識階段,從1898年第一個AAB屬,醋酸桿菌屬的提出至1989年的91年中,僅僅報道了醋酸桿菌屬、葡糖桿菌屬和酸單孢菌屬,共3個AAB屬,約10種。第二階段從1989年至今,是AAB分類的發(fā)展階段。在這25年中,基本上平均以每兩年報道1個AAB新屬、3個新種的速度發(fā)展。截至2014年初,已報道了16個AAB屬,共84個種(見表2)。這一方面與分子生物學(xué)技術(shù)用于AAB的輔助分類有關(guān),另一方面,與AAB新的生物學(xué)功能發(fā)現(xiàn)、研究和應(yīng)用也密不可分[24]。如HESTRIN S等[25]1947年就在《Nature》上發(fā)表了[木桿菌(A.xylinum),1997年和2012年先后更名為木葡糖醋桿菌(Ga.xylinus)和木駒形桿菌(K.xylinus)]可生產(chǎn)纖維素的研究結(jié)果。此后的研究發(fā)現(xiàn),柿子葡糖醋桿菌(Ga.persimmonis)、柿醋駒形桿菌(K.kakiaceti)、麥德林駒形桿菌(K.medellinensis)等AAB也能產(chǎn)生纖維素[26-28]。
隨著越來越多的AAB被發(fā)現(xiàn),AAB種屬命名、名稱變更以及種屬特征的描述等帶來了很大的挑戰(zhàn)。在以下的內(nèi)容中將主要敘述AAB命名原則、AAB種屬名稱演變以及AAB的種屬特征。
2.1 AAB命名原則
AAB的命名原則與其他微生物的命名原則基本相同,屬名主要是根據(jù)AAB特性或某位杰出科學(xué)家的姓氏來命名。如糖桿菌屬(Saccharibacter)因具有耐高濃度糖(Saccharon=sugar)的能力且為桿狀(bacter=baktron=rod)而得名;塔堤查仁桿菌屬(Tanticharoenia)以泰國微生物學(xué)家Morakot Tanticharoen的姓氏命名。而AAB種名則主要是根據(jù)菌落顏色、菌體形態(tài)、菌種特性、分離樣品來源、采樣地名稱、菌株分離與命名者或者某位杰出科學(xué)家的姓氏等來命名。以采樣地名稱命名時,一般用采樣地城市或國家名后+后綴“ensis”作為種名,如芝庇儂醋酸桿菌(A.cibinongensis)由芝比儂(Cibinong,位于印尼爪哇島西部的一座城市)+后綴(ensis)組成;加納醋酸桿菌(A.ghanensis)由加納(Ghana,位于非洲西部的一個國家)+后綴(ensis)組成;沖繩醋酸桿菌(A.okinawensis)由沖繩(Okinawa,位于日本九州島和中國臺灣省之間,舊稱琉球)+后綴(ensis)組成。以分離樣品命名時,一般是直接引用或適當修改樣品名稱來命名,如啤酒醋酸桿菌(A.cerevisiae)表明該AAB分離自啤酒(cerevisiae);古墳葡糖醋桿菌(Ga.aggeris)表明該AAB分離自古墳(aggeris=mound);蒲桃醋酸桿菌(A.syzygii)表明該AAB分離自蒲桃屬(Syzygium)的植物。以AAB特性命名時,一般是以AAB特性或特性的縮寫組合來命名,如白葡糖桿菌(G.albidus)表明該AAB菌落為白色(albidus=white);球形葡糖桿菌(G.sphaericus)表明該AAB菌體為球形(spherical);固氮葡糖醋桿菌(Ga.azotocaptans)的種名由氮(azotum=nitrogen)+固定(captans=catching)組成,表明該菌具有固定氮氣的能力(特征)。以微生物學(xué)家姓氏命名時,一般是為紀念在AAB研究領(lǐng)域作出了具大貢獻的微生物家,以其姓氏為主體并作適當修改來命名,如內(nèi)村葡糖桿菌(G.uchimurae)為紀念日本微生物學(xué)家Tai Uchimura(主要從事AAB的系統(tǒng)研究,尤其是朝井桿菌、醋酸桿菌、葡糖桿菌和葡糖醋酸桿菌菌株),以其姓氏內(nèi)村(Uchimura)進行命名;婉貞葡糖桿菌(G.wancherniae)為紀念泰國微生物學(xué)家Wanchern Potacharoen(主要從事AAB系統(tǒng)學(xué)研究);圓谷葡糖醋桿菌(Ga.entanii)為紀念日本微生物學(xué)家Etsuzo Entani(最先從事酒精醋中AAB的分離與繁殖方面的研究工作);約翰娜葡糖醋桿菌(Ga.johannae)為紀念巴西微生物學(xué)家Johanna D?bereiner(第一個分離到葡糖醋桿菌屬中具有固氮功能的AAB)。
2.2 AAB種屬名稱的演變
根據(jù)2005年出版的第九版《伯杰氏系統(tǒng)細菌學(xué)手冊》第2卷,AAB屬于變形桿菌門(Proteobacteria)α-變形菌菌綱(α-Proteobacteria)紅螺菌目(Rhodospirillales)醋桿菌科(Acetobacteraceae),記載了6個AAB屬,共31個種。圖1是第九版《伯杰氏系統(tǒng)細菌學(xué)手冊》第2卷中,采用最大簡值法對收錄的葡糖醋桿菌屬、酸單胞菌屬、朝井桿菌、醋酸桿菌屬和葡糖桿菌屬的31個AAB構(gòu)建的進化樹。其中,醋酸桿菌屬是醋桿菌科的典型屬。
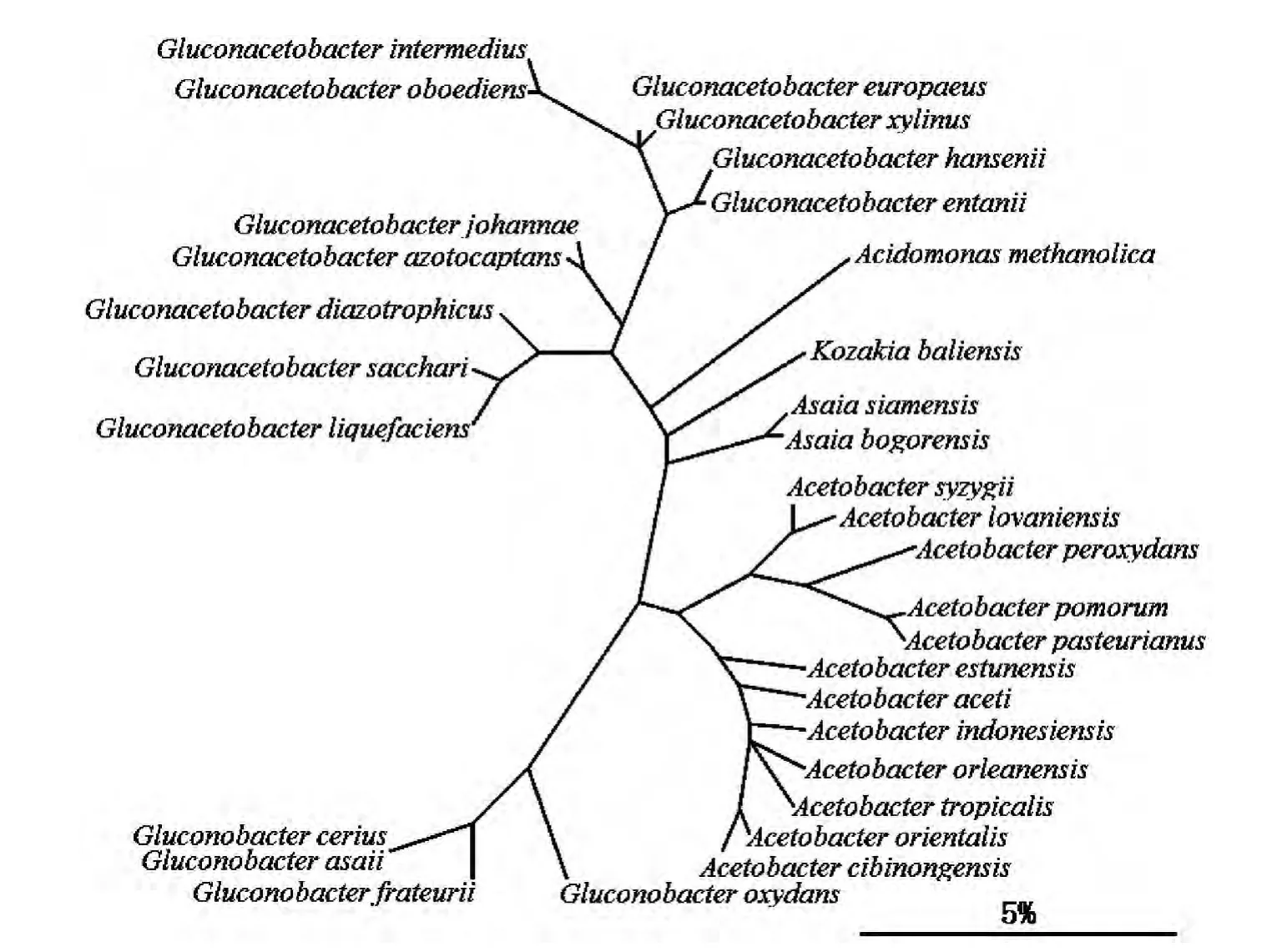
圖1 葡糖醋桿菌屬、酸單胞菌屬、朝井桿菌、醋酸桿菌屬和葡糖桿菌屬的親緣關(guān)系[29]Fig.1 Relationship between Gluconacetobacter, Acidomonas, Asaia, Acetobacter and Gluconobacter
像其他微生物一樣,隨著對AAB分類特征的進一步觀察和分析,各種新的分類方法和手段的不斷引入以及對微生物功能(作用)的拓展與再認識,它們的分類地位和種屬名稱一直在變化之中。如前所述,AAB的分類方法,經(jīng)歷了由以形態(tài)特征為主,到輔以化學(xué)成分特征與遺傳特征,再到三種分類方法相結(jié)合的發(fā)展,而對AAB功能的認識也從原來認為AAB可以分泌醋酸等有機酸到有些AAB具有產(chǎn)生纖維素和固氮等能力。此外,AAB的輔酶Q(ubiquinone,UQ)與菌體脂肪酸組成特性被用作其分類依據(jù)之一,也都推動了其分類地位與種屬名稱的變化。下面將通過對AAB中一些重要的種屬分類地位和名稱變化進行概述。
在AAB的表型分類學(xué)方面,自從1898年第一個AAB屬,醋酸桿菌屬提出后,研究發(fā)現(xiàn),該屬中的有些AAB不但可以分別利用葡萄糖和酒精產(chǎn)生葡萄糖酸和醋酸,還可以進一步以醋酸(鹽)作為碳源,將其分解成CO2并產(chǎn)生能量;而另外一些AAB對酒精的利用能力很弱,或者不能利用酒精作為碳源。依據(jù)這些特點,1935年,ASAI T[30]將醋酸桿菌屬中氧化酒精能力弱,且不能氧化醋酸(鹽)的AAB歸屬為另一個AAB屬——葡糖桿菌屬;1954年LEIFSON E[31]根據(jù)醋酸桿菌屬中AAB菌體的鞭毛著生情況和對醋酸鹽氧化能力,建議將極生鞭毛且不氧化醋酸鹽的AAB獨立為一個新屬——“醋單胞菌屬(Acetomonas)”。由于葡糖桿菌屬和“醋單胞菌屬”的特征極為相似,所以1961年,DE LEY J[32]通過進一步研究,明確了葡糖桿菌屬為第二個AAB屬,并確立了氧化葡糖桿菌(G.oxydans)為該屬典型種的地位,從此,“醋單胞菌屬”逐漸停止了使用。
眾所周知,由于形態(tài)分類方法依據(jù)的菌落特征、菌體形態(tài)等特征不穩(wěn)定,容易受到培養(yǎng)基組成和培養(yǎng)條件變化等的影響,所以輔酶Q類型和細菌膜脂肪酸組成等作為形態(tài)分類的輔助手段被廣泛用于AAB的分類中。輔酶Q是一種廣泛存在于微生物細胞膜中的脂溶性醌類化合物,其結(jié)構(gòu)與維生素K、維生素E與質(zhì)體醌相似,參與呼吸系統(tǒng)的電子傳遞。AAB的輔酶Q主要包括Q-9和Q-10兩種類型(Q-9的側(cè)鏈含有9個異戊二烯單元,Q-10的側(cè)鏈含有10個異戊二烯單元)[33]。1968年和1981年,YAMADA Y等[34-35]先后探討了AAB中輔酶Q類型和脂肪酸組成特點,發(fā)現(xiàn)它們在不同的AAB中存在較大差異,可以用來對AAB進行分類,所以1983年,YAMADA Y等[36]將輔酶Q-10的醋化醋酸桿菌木醋桿菌亞種(A.acetisubsp.xylinum)提升為木醋酸桿菌(A.xylinum);1985年,YAMADA Y等[33]又將醋酸桿菌屬中Q-10型的AAB歸入醋酸桿菌屬葡糖酸醋酸桿菌亞屬(Acetobacter(subgen.Gluconoacetobacter)),這樣醋酸桿菌屬就形成了分別具有輔酶Q-9和Q-10的兩類AAB。1989年,URAKAMI T等[34]通過分析醋酸桿菌屬中AAB和嗜酸菌屬(Acidiphilium)菌株的形態(tài)學(xué)特征、生理生化特征、輔酶Q的類型以及脂肪酸組成,將其中具有甲醇氧化能力的AAB獨立為一個新屬——酸單胞菌屬(Acidomonas,Ac.),并提出了甲醇酸單孢菌種(Ac.methanolica)[32]。但是,葡糖酸醋酸桿菌亞屬和酸單胞菌屬的分類地位曾一度受到質(zhì)疑,直到1997年,YAMADA Y等[57]通過16S rRNA序列分析確定了酸單胞菌屬的分類地位,所以至1989年,醋酸桿菌屬、葡糖桿菌屬和酸單胞菌屬,共3個AAB屬被提出。
隨著分子生物學(xué)方法的快速發(fā)展,遺傳學(xué)信息逐漸成為AAB分類的重要參考依據(jù)。1994年,SIEVERS M等[37]基于16S核糖體DNA序列信息對醋桿菌科(Acetobacteraceae)的四個屬:醋酸桿菌屬、葡糖桿菌屬、紅球狀菌屬和嗜酸菌屬的系統(tǒng)發(fā)育進行了研究,發(fā)現(xiàn)醋酸桿菌屬和葡糖桿菌屬是變形菌門(Proteobacteria)α-亞門中一個獨立分支。1997年,YAMADA Y等[57]基于16S rRNA的序列信息,將葡糖酸醋酸桿菌亞屬提升為葡糖酸醋酸桿菌屬(Gluconoace tobacter),并以液化葡糖酸醋酸桿菌(Gluconoacetobacter liquefaciens)為典型菌株。同年,根據(jù)國際細菌命名法規(guī),Gluconoacetobacter被正式更名為Gluconacetobacter(Ga.),并沿用至今。自此,基于形態(tài)、化學(xué)和遺傳學(xué)特征相結(jié)合的AAB分類方法被普遍采用。隨后,朝井桿菌屬(Asaia,As.)[38]、公崎桿菌屬(Kozakia,Ka.)[39]、斯瓦米納坦桿菌屬(Swaminathania,Sw.)[8]、糖桿菌屬(Saccharibacter,S.)[4]、新朝井桿菌屬(Neoasaia,N.)[40]、顆粒桿菌屬(Granulibacter,Gr.)[41]和塔堤查仁桿菌屬(Tanticharoenia,T.)[42]等新的AAB屬先后被提出。
從21世紀初開始,為了明確AAB的系統(tǒng)進化關(guān)系,一些統(tǒng)計學(xué)方法結(jié)合數(shù)據(jù)處理軟件也開始應(yīng)用于AAB的分類,形成了基于鄰接法(neighbor-joining method)、最大簡約法(maximum parsimony)和最大似然估計(maximum likeli hood method)構(gòu)建系統(tǒng)樹的AAB分類方法[43]。YUKPHAN P等[43]采用這三種方法,基于AAB的16S rRNA序列分析建立其系統(tǒng)樹,再結(jié)合生理生化特性,發(fā)現(xiàn)從泰國清邁紅姜花朵中分離到AAB菌株不同于已報道的AAB屬,于是提出了一個新AAB屬—雨山桿菌屬(Ameyamaea,Am.)。2011年,YUKPHAN P等[44]又對從泰國馬櫻丹和翅莢決明的花中分離的AAB進行了鑒定,根據(jù)細胞膜脂肪酸組成分析和系統(tǒng)樹分析結(jié)果,提出了另外一個AAB新屬——新駒形桿菌屬(Neokomagataea,Ne.)。目前,基于形態(tài)與化學(xué)特征,結(jié)合鄰接法、最大簡約法、最大似然估計等遺傳進化分析結(jié)果的AAB分類方法被普遍采用。YAMADA Y等[45]根據(jù)此分類方法,從葡糖醋桿菌屬中分離出駒形桿菌屬(Komagataeibacter,K.),并以木駒形桿菌(K.xylinus)為典型種。阮桿菌屬(Nguyenibacter,Ng.)[46]和斯溫斯桿菌屬(Swingsia,Sw.)[47]也是在此分類學(xué)方法下被發(fā)現(xiàn)的。2014年初,YAMADA Y[66]又根據(jù)此分類法將曾歸類到葡糖醋桿菌屬的3個AAB(Ga.kakiaceti,Ga.medellinesis和Ga.maltaceti)重新歸類于駒形桿菌屬,命名為柿醋駒形桿菌(K.kakiaceti)、麥德林駒形桿菌(K.medellinesis)和麥芽醋駒形桿菌(K.maltaceti)。至2014年初,已報道的AAB包括16個屬,共84種(見表2)。

表2 AAB的種屬分類Table 2 Species and genus classification of AAB
下面將就這些AAB屬及其代表性種的主要特征和用途進行歸納和分析。考慮到篇幅的限制,關(guān)于AAB各屬典型菌種的生理生化特征、輔酶Q類型、細胞脂肪酸組成以及DNA的G+C摩爾百分含量等更多具體信息可參考其中的相關(guān)文獻。
2.3 AAB的種屬特征
2.3.1 醋酸桿菌屬(Acetobacter,A.)
醋酸桿菌因能產(chǎn)醋(aceti)且為桿菌(bacter=baktron=rod)而得名。醋酸桿菌屬菌株不能運動,能氧化乙酸鹽和乳酸鹽,能氧化乙醇產(chǎn)乙酸,泛醌為Q-9型,其典型菌種為醋化醋酸桿菌(A.aceti)。醋化醋酸桿菌可能是正式報道發(fā)現(xiàn)的第一個AAB屬,廣泛用于食醋的生產(chǎn)。醋化醋酸桿菌的應(yīng)用研究主要包括食醋的液態(tài)發(fā)酵工藝條件優(yōu)化[48]、固定化細胞生產(chǎn)食醋[49]和兩步法生產(chǎn)纖維素[50]。此外,醋化醋酸桿菌的耐酸機制方面,包括質(zhì)子泵、ABC盒轉(zhuǎn)運蛋白、應(yīng)激蛋白、特殊三羧酸循環(huán)、順烏頭酸酶和檸檬酸合成酶也有大量的研究報道[51]。
2.3.2 酸單胞菌屬(Acidomonas,Ac.)
酸單孢菌屬因其嗜酸(acid)且為單細胞(monas=monad)生長而得名。酸單胞菌屬菌株不產(chǎn)芽胞,圓頭桿狀,主要以單個存在,很少成對出現(xiàn),不能運動,能氧化酒精生產(chǎn)醋酸,能在pH 2.0~5.5條件下生長,輔酶Q為Q-10型,DNA的G+C含量為64.4%。該屬典型菌種為甲醇酸單胞菌(Ac.methanolica)。甲醇酸單胞菌是酸單胞菌屬唯一的種,能同化甲醇,其典型菌株TK 0705(=IMET 10945T)分離自酵母發(fā)酵淤泥[34]。
2.3.3 雨山桿菌屬(Ameyamaea,Am.)
雨山桿菌屬以日本山口大學(xué)Minoru Ameyama教授的姓氏命名。該屬菌體呈桿狀,極生鞭毛,能運動,能氧化酒精生產(chǎn)食醋,氧化醋酸鹽能力強而氧化乳酸鹽能力較弱,能在含0.35%乙酸的培養(yǎng)基上生長,輔酶Q為Q-10型,DNA的G+C含量為66.0%~66.1%。典型菌種清邁雨山桿菌(Am.chiangmaiensis),以菌株分類地泰國清邁(Chiang Mai)命名,典型菌株AC04T(=BCC 15744T=NBRC 103196T)分離自泰國紅姜的花朵[43]。
2.3.4 朝井桿菌屬(Asaia,As.)
朝井桿菌屬以日本細菌學(xué)家Toshinobu Asai的姓氏命名。該屬菌株呈桿狀,周生鞭毛,能運動,能氧化乙酸鹽和乳酸鹽,不能氧化酒精生產(chǎn)食醋,泛醌為Q-10型,DNA的G+C含量為59.3%~61.0%,該屬的典型菌種為茂物朝井桿菌(As.bogorensis)。茂物朝井桿菌典型菌株為71T(=JCM 10569T=NRIC 9311T),分離自印度尼西亞的紫荊花,不能或極微弱的氧化酒精產(chǎn)醋酸,不能在含0.35%醋酸的培養(yǎng)基上生長[38]。
朝井桿菌屬菌株主要分離自植物的花朵,普遍耐滲透壓但不能產(chǎn)酸。該屬的一些菌株為昆蟲的共生菌,也是引起飲料腐敗的主要微生物,茂物朝井桿菌(As.bogorensis)和蘭那朝井桿菌(As.lannaensis)也可能是人類的條件致病菌[5,38,52]。朝井桿菌也具有一些潛在的應(yīng)用價值,如用于粘附劑[53]和虐疾控制[54],生物固氮[55]和產(chǎn)生超細纖維素[56]。
2.3.5 葡糖桿菌屬(Gluconobacter,G.)[30]
葡糖桿菌因能氧化葡萄糖(glucose)且為桿菌(bacter)而得名。葡糖桿菌屬菌株具有極生鞭毛,能運動,能氧化酒精產(chǎn)乙酸,但不能氧化乙酸鹽和乳酸鹽,泛醌為Q-10型,該屬典型菌種為氧化葡糖桿菌(G.oxydans)。氧化葡糖桿菌也是最早被發(fā)現(xiàn)的AAB之一,其相關(guān)研究很多,主要包括三個方面:生化產(chǎn)品生產(chǎn)、酶學(xué)研究和傳感器開發(fā)。在生物產(chǎn)品生產(chǎn)方面,應(yīng)用氧化葡糖桿菌可以生產(chǎn)2,5-二酮葡萄糖酸、2-酮-L-葡萄糖酸鹽、抗壞血酸、D-葡萄糖酸、山梨酮糖和半乳糖酸等;酶學(xué)研究包括葡萄糖脫氫酶、葡萄糖酸脫氫酶、2-酮葡萄糖酸脫氫酶、山梨酮糖脫氫酶和乙醛脫氫酶等;傳感器開發(fā)包括用氧化葡糖桿菌純化的酶用于酒精、糖、乳糖和木糖的快速檢測[24]。
2.3.6 葡糖醋桿菌屬(Gluconacetobacter,Ga.)[57]
葡糖醋桿菌因能氧化葡糖酸鹽(gluconate)和醋酸鹽(acetate)且為桿菌(bacter)而得名。葡糖醋桿菌屬菌株具有周生鞭毛,能運動,能氧化酒精產(chǎn)醋,輔酶Q為Q-10型,DNA的G+C百分含量為64.4%。該屬典型菌種為液化葡糖醋桿菌(Ga.liquefaciens),它引起菠蘿發(fā)生紅肉病的病原菌[58],也可以用來降解甲基紅[59]和生產(chǎn)2-酮-古洛糖酸[60]。液化葡糖醋桿菌曾被歸屬于醋化醋桿菌亞種(A.aceti subsp.liquefaciens)(Asai 1935)。1983年,YAMADA Y[69]將其提升為種,命名為液化醋桿菌(A.liquefaciens)。1997年,YAMADA Y等[57]基于16S rRNA序列分析,將液化醋桿菌(A.liquefaciens)更名為液化葡糖醋桿菌(Ga.liquefaciens)。
該屬的相關(guān)種研究較多,并具有多種功能。例如,古墳葡糖醋桿菌(Ga.aggeris)、固氮葡糖醋桿菌(Ga.azotocaptans)、固氮葡糖醋桿菌(Ga.diazotrophicus)、約翰娜葡糖醋桿菌(Ga.johannae)和高松冢葡糖醋桿(Ga.takamatsuzukensis)具有固氮作用[11,61];柿子葡糖醋桿菌(Ga.persimmonis)能生產(chǎn)纖維素[27];約翰娜葡糖醋桿菌(Ga.johannae)、液化葡糖醋桿菌(Ga.liquefaciens)、甘蔗葡糖醋桿菌(Ga.sacchari)和古墓土壤葡糖醋桿菌(Ga.tumulisoli)能生產(chǎn)水溶性棕色素[11,62-63];圓谷葡糖醋桿菌(Ga.entanii)能耐受較高濃度乙醇和乙酸[64];固氮葡糖醋桿菌(Ga.azotocaptans)、固重氮葡糖醋桿菌(Ga.diazotrophicus)、約翰娜葡糖醋桿菌(Ga.johannae)和甘蔗葡糖醋桿菌(Ga.sacchari)能耐受較高濃度葡萄糖[7,28,62]。
2.3.7 顆粒桿菌屬(Granulibacter,Gr.)
顆粒桿菌因產(chǎn)生粒斑或肉芽腫(肉芽腫病患者臨床特征和淋巴結(jié)切片均為顆粒狀)而得名。該屬菌體呈球桿狀,不能運動,菌體為球狀或桿狀,可生產(chǎn)黃色素,能氧化醋酸鹽和乳酸鹽,能氧化酒精產(chǎn)醋酸,適合在含較高濃度葡萄糖的培養(yǎng)基中生長,輔酶Q為Q-10型,DNA的G+C含量為59.1%。該屬典型菌種為貝塞斯達顆粒桿菌(Gr.bethesdensis),以菌株分離地美國貝塞斯達(Bethesda)命名,典型菌株CGDNIH1T(=ATCC BAA-1260T=DSM 17861T)分離自患有慢性肉芽腫病病人的淋巴結(jié),被視為條件致病菌[41]。
2.3.8 駒形桿菌屬(Komagataeibacter,K.)
以日本微生物學(xué)家Kazuo Komagata的姓氏命名。[45]菌體不能運動,能氧化醋酸鹽和乳酸鹽,能化酒精產(chǎn)醋酸,典型菌種木駒形桿菌(K.xylinus),是最早發(fā)現(xiàn)能產(chǎn)纖維素的AAB,所以相關(guān)研究非常透徹[63]。
駒形桿菌屬主要是由葡糖醋桿菌屬分化出來的。早在1999年,F(xiàn)RANKE I H等[62]發(fā)現(xiàn)葡糖醋桿菌屬系統(tǒng)樹中存在二象性,2000年,YAMADA Y等[67]將葡糖醋桿菌屬分成兩個子集。之后也陸續(xù)有報道稱發(fā)現(xiàn)此現(xiàn)象,YAMADA Y等[3]報道稱Ga.liquefaciens(葡糖醋桿菌屬的典型種)和Ga.xylinus在表型特征、遺傳學(xué)特征和生態(tài)學(xué)特征上存在屬級的差異。直到2012年,YAMADA Y等[68]提出由葡糖醋桿菌屬的12個種組成新AAB屬——駒形桿菌屬,但是根據(jù)《細菌學(xué)代碼》27條規(guī)則,該屬典型菌株存在爭議,同年,YAMADA Y等[45]再一次提出此屬,并將該屬詳細信息進行了總結(jié)。
該屬AAB除包括可以產(chǎn)纖維素的木駒形桿菌(K.xylinus)、萊蒂亞駒形桿菌(K.rhaeticus)、斯溫駒形桿菌(K.swingsii)、蔗糖駒形桿菌(K.sucrofermentans)、柿醋駒形桿菌(K.kakiaceti)、麥德林駒形桿菌(K.medellinensis)和hansenii(K.hansenii)種外,還包括高耐酸(醇)的歐洲駒形桿菌(K.europaeus)、中間駒形桿菌(K.intermedius)、溫馴駒形桿菌(K.oboediens)、麥芽醋駒形桿菌(K、maltaceti)(較弱)。
2.3.9 公崎桿菌屬(Kozakia,Ka.)
公崎桿菌屬以日本微生物學(xué)家Michio Kozaki的姓名命名的。該屬菌株為桿狀,不能運動,能利用蔗糖或D-果糖生產(chǎn)類似果聚糖的黏性物質(zhì),能氧化醋酸鹽和乳酸鹽,能利用酒精生產(chǎn)食醋,輔酶Q為Q-10型,DNA的G+C含量為56.8%~57.2%。巴厘島公崎桿菌(Ka.baliensis)是該屬的典型菌種,也是唯一的種。它以采樣地印度尼西亞巴厘島(Bali)命名,其典型菌株Yo-3T(=IFO 16664T=JCM 11301T=DSM 14400T)分離自巴厘島的紅糖中[39]。
2.3.10 新朝井桿菌屬(Neoasaia,N.)
以日本微生物學(xué)家Toshinobu Asai的姓氏命名(Neoasaia=new Asaia)。該屬菌落為粉紅色,能氧化酒精產(chǎn)醋酸,能氧化醋酸鹽和乳酸鹽,輔酶Q為Q-10型,DNA的G+C含量為63.1%。訪屬典型菌種為清邁新朝井桿菌(N.chiangmaiensis),以菌株分離地泰國清邁(Chiang Mai)命名,典型菌株AC28T(=BCC 15763T=NBRC 101099T)分自離泰國紅姜的花朵[40]。
2.3.11 新駒形桿菌屬(Neokomagataea,Ne.)
以日本微生物學(xué)家Kazuo Komagata的姓氏命名。菌體呈桿狀,不能運動,氧化酒精產(chǎn)醋酸能力較弱,不能氧化醋酸鹽和乳醋鹽,能在含30%(w/v)葡萄糖的培養(yǎng)基上生長,但不能在含0.35%(w/v)醋酸的培養(yǎng)基上生長,輔酶Q為Q-10型,DNA的G+C含量為51.2%~56.8%。該屬包括泰國新駒形桿菌(Ne.thailandica)和譚島新駒形桿菌(Ne.tanensis),均以菌種分離地命名。前者能氧化蜜二糖和蜜三糖產(chǎn)生有機酸,而后者不能。兩者的典型菌株分別是AH11T(=BCC 25710T=NBRC 106555T)和AH13T(=BCC 25711T=NBRC 106556T),均分離自泰國的花朵[44]。
2.3.12 阮桿菌屬(Nguyenibacter,Ng.)
以越南微生物學(xué)家Dung Lan Nguyen的姓氏命名。菌體呈桿狀,周生鞭毛,能運動,能氧化醋酸鹽但不能氧化乳酸鹽,不能氧化酒精產(chǎn)食醋,在含30%葡萄糖或0.35%乙酸的培養(yǎng)基上弱生長,能生產(chǎn)水溶性棕色素,有固氮能力,可在不含氮源的LGI培養(yǎng)基上生長,輔酶Q為Q-10型,DNA的G+C含量為68.1%~69.4%。典型菌種為安南阮桿菌(Ng.vanlangensis),以菌種分離地越南(Vietnam,舊稱Vanlang)命名,典型菌株TN01LGIT(=BCC 54744T=NBRC 109046T)分離自越南水稻根部[65]。
2.3.13 糖桿菌屬(Saccharibacter,S.)
糖桿菌屬因其具有耐高濃度糖的能力而得名。該屬菌體為直桿狀,不能運動,能在含2%~40%(w/v)糖類的培養(yǎng)基中生長,利用酒精生產(chǎn)乙酸的能力極弱,不能氧化醋酸鹽,輔酶Q為Q-10型,DNA的G+C含量為52%~53%,該屬典型菌種為花糖桿菌(S.floricola),典型菌株為S-877T(=AJ 13480T=JCM 12116T=DSM 15669T)分離自日本神奈川縣的櫻花花粉[4]。
2.3.14 斯瓦米納坦桿菌屬(Swaminathania,Sw.)
斯瓦米納坦桿菌屬以印度生物學(xué)家Swaminathan的姓名命名。該屬菌體呈圓頭桿狀,周生鞭毛,能運動,能氧化酒精產(chǎn)醋酸,能氧化醋酸鹽和乳酸鹽,能生產(chǎn)水溶性棕色素,能固氮和同化水溶性磷酸鹽,輔酶Q為Q-10型,DNA的G+C含量為57.6%~59.9%。典型菌種為耐鹽斯瓦米納坦桿菌(Sa.salitolerans),其典型菌株P(guān)A51T(=LMG 21291T=MTCC3853T)分離自印度野生水稻根系[8]。
2.3.15 斯溫斯桿菌屬(Swingsia,Si.)
以比利時根特大學(xué)微生物學(xué)家Jean Swings的姓氏命名。該屬菌體不能運動,能利用酒精生產(chǎn)食醋,不能氧化醋酸鹽或乳酸鹽,能在含30%(w/v)葡萄糖的培養(yǎng)基上生長,但不能耐受0.35%(w/v)醋酸,能生產(chǎn)水溶性棕色素,輔酶Q為Q-10型,DNA的G+C含量為46.9%~47.3%。蘇梅斯溫斯桿菌(Si.samuiensis)是其唯一的種,典型菌株AH83T(=BCC 25779T=NBRC 107927T),種名因菌株分離地泰國他尼府蘇梅島(Samui island)而得名[47]。
2.3.16 塔堤查仁桿菌屬(Tanticharoenia,T.)
以泰國微生物學(xué)家Morakot Tanticharoen的姓氏命名。該屬菌體呈桿狀,能氧化酒精生產(chǎn)食醋,能生產(chǎn)水溶性棕色素,不能運動,不能氧化乙酸鹽或乳酸鹽,可以在含30%(w/v)葡萄糖的培養(yǎng)基中生長,耐滲透壓的能力較強,但僅能耐受0.35%的乙酸,輔酶Q為Q-10型,DNA的G+C含量為64.5%~65.6%。典型菌種為薩克塔塔堤查仁桿菌(T.sakaeratensis),以菌種分離地泰國呵叻府薩克塔(Sakaerat)命名,典型菌株為NBRC 103193T(=BCC 15772T=NBRC 103193T)分離自泰國土壤[42]。
3 總結(jié)與展望
AAB是革蘭氏陰性細菌,主要分離自發(fā)酵食品、植物器官、動物器官和土壤。分離時,可將AAB分為三類:普通AAB、耐酸(醇)AAB和固氮AAB。不同類型AAB的分離培養(yǎng)基和分離方法有所不同。隨著分子生物技術(shù)的發(fā)展,目前AAB的鑒定和分類主要是結(jié)合表觀特征(形態(tài)學(xué)特征和營養(yǎng)代謝)、化學(xué)成分特征(輔酶Q類型和細胞膜脂肪酸組成)和遺傳學(xué)特征(基于NJ、MP和ML分析16S rRNA序列)三方面信息來進行。自1898年,BEIJERINCK M[2]定義了第一個AAB屬以來,截至2014年初,已報道的AAB有16個屬,共84種。像其他微生物一樣,AAB的種屬分類一直在變化,而且會一直變化下去。一百多年以來,隨著AAB鑒定和分類方法的發(fā)展,AAB種屬分類了發(fā)生了重大的轉(zhuǎn)變,一方面,不斷有新屬和新種陸續(xù)被發(fā)現(xiàn);另一方面,一些AAB被重新歸類。AAB最初因具有產(chǎn)酸能力而得名,但近年來研究發(fā)現(xiàn):有些AAB并不具有產(chǎn)酸能力,而有些AAB除了產(chǎn)生醋酸外,還具有很多其它功能,如產(chǎn)生纖維素、色素、吲哚乙酸、抗壞血酸和固氮等,正是這些功能和特性促進了AAB研究的發(fā)展。目前,AAB生產(chǎn)纖維素和固氮研究較為透徹,技術(shù)也漸趨成熟。AAB屬各具特色,包括耐滲透壓AAB、耐酸(醇)AAB、固氮AAB、產(chǎn)纖維素AAB等等。AAB的特性(如耐酸、耐醇、耐滲透壓和耐熱性)和功能(如固氮以及生產(chǎn)纖維素、吲哚乙酸、抗壞血酸和水溶性色素)已成為AAB的研究熱門。
[1]MATSUSHITA K,INOUE T,ADACHI O,et al.Acetobacter acetipossesses a proton motive force-dependent efflux system for acetic acid [J].J Bacteriol,2005,187(13):4346-4352.
[2]BEIJERINCK M.Ueber die arten der essigbakterien[J].Centralblatt für Bakteriologie,Parasitenkunde und Infektionskrankheiten,1898,2(4):209-216.
[3]YAMADA Y,YUKPHAN P.Genera and species in acetic acid bacteria[J].Int J Food Microbiol,2008,125(1):15-24.
[4]JOJIMA Y,MIHARA Y,SUZUKI S,et al.Saccharibacter floricolagen.nov.,sp.nov.,a novel osmophilic acetic acid bacterium isolated from pollen[J].Int J Syst Evol Micr,2004,54(6):2263-2267.
[5]MOORE J E,MCCALMONT M,XU J,et al.Asaiasp.,an unusual spoilage organism of fruit-flavored bottled water[J].Appl Environ Microbiol,2002,68(8):4130-4131.
[6]BASTIáN F,COHEN A,PICCOLI P,et al.Production of indole-3-acetic acid and gibberellins A1and A3byAcetobacter diazotrophicusandHerbaspirillum seropedicaein chemically-defined culture media[J].J Plant Growth Regul,1998,24(1):7-11.
[7]FUENTES-RAMáREZ L E,BUSTILLOS-CRISTALES R,TAPIAHERNáNDEZ A,et al.Novel nitrogen-fixing acetic acid bacteria,Gluconacetobacter johannaesp.nov.andGluconacetobacter azotocaptanssp.nov.,associated with coffee plants[J].Int J Syst Evol Micr,2001,51(4):1305-1314.
[8]LOGANATHAN P,NAIR S.Swaminathania salitoleransgen.nov.,sp.nov.,a salt-tolerant,nitrogen-fixing and phosphate-solubilizing bacterium from wild rice(Porteresia coarctataTateoka)[J].Int J Syst Evol Micr,2004,54(4):1185-1190.
[9]NGUYEN V T,FLANAGAN B,GIDLEY M J,et al.Characterization of cellulose production by aGluconacetobacter xylinusstrain from Kombucha[J].Curr Microbiol,2008,57(5):449-453.
[10]REWATKAR A R.Microbial production of L-ascorbic acid from sorbitol[J].Biotechnol Resour,2010,3:268-274.
[11]ENTANI E,OHMORI S,MASAI H,et al.Acetobacter polyoxogenessp.nov.,a new species of an accetic acid bacterium useful for producing vinegar with high acidity[J].J Gen Appl Microbiol,1985,31(5):475-490.
[12]YAMADA Y,HOSONO R,LISDYANTI P,et al.Identification of acetic acid bacteria isolated from Indonesian sources,especially of isolates classified in the genusGluconobacter[J].J Gen Appl Microbiol,1999,45(1):23-28.
[13]SCHULLER G,HERTEL C,HAMMES W P.Gluconacetobacter entaniisp.Nov.,isolated from submerged high-acid industrial binegar fermentations[J].Int J Syst Evol Micr,2000,50:2013-2020.
[14]DUTTA D,GACHHUI R.Nitrogen-fixing and cellulose-producingGluconacetobacter kombuchaesp.nov.,isolated from Kombucha tea[J].Int J Syst Evol Micr,2007,57(2):353-357.
[15]GILLIS M,DE LEY J.Intra-and intergeneric similarities of the ribosomal ribonucleic acid cistrons ofAcetobacterandGluconobacter[J].Int J Syst Bacterio,1980,30(1):7-27.
[16]POBLET M,ROZèS N,GUILLAMóN J M,et al.Identification of acetic acid bacteria by restriction fragment length polymorphism analysis of a PCR-amplified fragment of the gene coding for 16S rRNA[J].Lett Appl Microbiol,2000,31(1):63-67.
[17]TRCEK J.Quick identification of acetic acid bacteria based on nucleotide sequences of the 16S-23S rDNA internal transcribed spacer region and of the PQQ-dependent alcohol dehydrogenase gene[J].Syst Appl Microbiol,2005,28(8):735-745.
[18]DE VERO L,GALA E,GULLO M,et al.Application of denaturing gradient gel electrophoresis(DGGE)analysis to evaluate acetic acid bacteria in traditional balsamic vinegar[J].Food Microbiol,2006,23(8):809-813.
[19]GULLO M,CAGGIA C,DE VERO L,et al.Characterization of acetic acid bacteria in“traditional balsamic vinegar”[J].Int J Food Microbiol,2006,106(2):209-212.
[20]DE VUYST L,CAMU N,DE WINTER T,et al.Validation of the(GTG)5-rep-PCR fingerprinting technique for rapid classification and identification of acetic acid bacteria,with a focus on isolates from Ghanaian fermented cocoa beans[J].Int J Food Microbiol,2008,125(1):79-90.
[21]TORIJA M J,MATEO E,GUILLAMóN J M,et al.Identification and quantification of acetic acid bacteria in wine and vinegar by TaqMan-MGB probes[J].Food Microbiol,2010,27(2):257-265.
[22]SOKOLLEK S J,HERTEL C,HAMMES W P.Cultivation and preservation of vinegar bacteria[J].J Biotechnol,1998,60(3):195-206.
[23]AZUMA Y,HOSOYAMA A,MATSUTANI M,et al.Whole-genome analyses reveal genetic instability ofAcetobacter pasteurianus[J].Nucleic Acids Res,2009,37(17):5768-5783.
[24]GUPTA A,SINGH V K,QAZI G,et al.Gluconobacter oxydans:its biotechnological applications[J].J Mol Microb Biotechnol,2001,3(3):445-456.
[25]HESTRIN S,ASCHNER M,MAGER J.Synthesis of cellulose by resting cells ofAcetobacter xylinum[J].Nature,1947,159(4028):64-65.
[26]CASTRO C,ZULUAGA R,áLVAREZ C,et al.Bacterial cellulose produced by a new acid-resistant strain ofGluconacetobactergenus[J].Carbohyd Polym,2012,89(4):1033-1037.
[27]HUNGUND B S,GUPTA S.Improved production of bacterial cellulose fromGluconacetobacter persimmonisGH-2[J].J Microbial Biochem Technol,2010,2(5):127-133.
[28]IINO T,SUZUKI R,TANAKA N,et al.Gluconacetobacter kakiacetisp.nov.,an acetic acid bacterium isolated from a traditional Japanese fruit vinegar[J].Int J Syst Evol Micr,2012,62(7):1465-1469.
[29]SIEVERS M,SWINGS J.Family II.Acetobacteraceae[M].2th edition.New York:Springer,2005.
[30]ASAI T.Taxonomic studies on acetic acid bacteria and allied oxidative bacteria isolated from fruits:a new classification of the oxidative bacteria[J].J Agr Chem Soc Jpn,1935,11(1):674-708.
[31]LEIFSON E.The flagellation and taxonomy of species ofAcetobacter[J].Anton Leeuw,1954,20(1):102-110.
[32]DE LEY J.Comparative carbohydrate metabolism and a proposal for a phylogenetic relationship of the acetic acid bacteria[J].Microbiology,1961,24(1):31-50.
[33]YAMADA Y.Gluconoacetobacter,a new subgenus comprising the acetate-oxidizing acetic acid bacteria with ubiquinone-10 in the genusAcetobacter[J].J Gen Appl Microbiol,1984,30(4):297-303.
[34]URAKAMI T,TAMAOKA J,SUZUKI K I,et al.Acidomonasgen.nov.,IncorporatingAcetobacter methanolicusasAcidomonas methanolicacomb.nov[J].Int J Syst Bacteriol,1989,39(1):50-55.
[35]YAMADA Y,NUNODA M,ISHIKAWA T,et al.The cellular fatty acid composition in acetic acid bacteria[J].J Gen Appl Microbiol,1981,27(5):405-417.
[36]YAMADA Y.Acetobacter xylinussp.nov.,nom.rev.,for the celluloseforming and cellulose-less,acetate-oxidizing acetic acid bacteria with the Q-10 system[J].J Gen Appl Microbiol,1983,29(5):417-420.
[37]SIEVERS M,LUDWIG W,TEUBER M.Phylogenetic positioning ofAcetobacter,Gluconobacter,RhodopilaandAcidiphiliumspecies as a branch of acidophilic bacteria in the α-subclass ofProteobacteriabased on 16S ribosomal DNA sequences[J].Syst Appl Microbiol,1994,17(2):189-196.
[38]YAMADA Y,KATSURA K,KAWASAKI H,et al.Asaia bogorensisgen.nov.,sp.nov.,an unusual acetic acid bacterium in the alpha-Proteobacteria[J].Int J Syst Evol Micr,2000,50(2):823-829.
[39]LISDIYANTI P,KAWASAKI H,WIDYASTUTI Y,et al.Kozakia baliensisgen.nov.,sp.nov.,a novel acetic acid bacterium in the alpha-proteobacteria[J].Int J Syst Evol Micr,2002,52(3):813-818.
[40]YUKPHAN P,MALIMAS T,POTACHAROEN W,et al.Neoasaia chiangmaiensisgen.nov.,sp.nov.,a novel osmotolerant acetic acid bacterium in the alpha-Proteobacteria[J].J Gen Appl Microbiol,2005,51(5):301-311.
[41]GREENBERG D E,PORCELLA S F,STOCK F,et al.Granulibacter bethesdensisgen.nov.,sp.nov.,a distinctive pathogenic acetic acid bacterium in the familyAcetobacteraceae[J].Int J Syst Evol Micr,2006,56(11):2609-2616.
[42]YUKPHAN P,MALIMAS T,MURAMATSU Y,et al.Tanticharoenia sakaeratensisgen.nov.,sp.nov.,a new osmotolerant acetic acid bacterium in the alpha-Proteobacteria[J].Biosci Biotech Biochem,2008,72(3):672-676.
[43]YUKPHAN P,MALIMAS T,MURAMATSU Y,et al.Ameyamaea chiangmaiensisgen.nov.,sp.nov.,an acetic acid bacterium in the alpha-Proteobacteria[J].Biosci Biotech Biochem,2009,73(10):2156-2162.
[44]YUKPHAN P,MALIMAS T,MURAMATSU Y,et al.Neokomagataeagen.nov.,with descriptions ofNeokomagataea thailandicasp.nov.andNeokomagataea tanensissp.nov.,osmotolerant acetic acid bacteria of the α-Proteobacteria[J].Biosci Biotech Biochem,2011,75(3):419-426.
[45]YAMADA Y,YUKPHAN P,LAN VU H T,et al.Description ofKomagataeibactergen.nov.,with proposals of new combinations (Acetobacteraceae)[J].J Gen Appl Microbiol,2012,58(5):397-404.
[46]MALIMAS T,CHAIPITAKCHONLATARN W,THI LAN VU H,et al.Swingsia samuiensisgen.nov.,sp.nov.,an osmotolerant acetic acid bacterium in the α-Proteobacteria[J].J Gen Appl Microbiol,2013,59(5):375-384.
[47]NISHIJIMA M,TAZATO N,HANDA Y,et al.Gluconacetobacter tumulisolisp.nov.,Gluconacetobacter takamatsuzukensissp.nov.andGluconacetobacter aggerissp.nov.,isolated from Takamatsuzuka Tumulus samples before and during the dismantling work in 2007[J].Int J Syst Evol Microbiol,2013,63(11):3981-3988.
[48]AWAD H,DIAZ R,MALEK R,et al.Efficient production process for food grade acetic acid byAcetobacter acetiin shake flask and in bioreactor cultures[J].E-Journal of Chemistry,2012,9(4):2275-2286.
[49]LOTONG N,MALAPAN W,BOONGORSRANG A,et al.Production of vinegar byAcetobactercells fixed on a rotating disc reactor[J].Appl Microbiol Biotechnol,1989,32(1):27-31.
[50]OKIYAMA A,SHIRAE H,KANO H,et al.Bacterial cellulose I.Twostage fermentation process for cellulose production byAcetobacter aceti[J].Food Hydrocolloid,1992,6(5):471-477.
[51]NAKANO S,FUKAYA M.Analysis of proteins responsive to acetic acid inAcetobacter:Molecular mechanisms conferring acetic acid resistance in acetic acid bacteria[J].Int J Food Microbiol,2008,125(1):54-59.
[52]KREGIEL D,RYGALA A,LIBUDZISZ Z,et al.Asaia lannensis-the spoilage acetic acid bacteria isolated from strawberry-flavored bottled water in Poland[J].Food Control,2012,26(1):147-150.
[53]KREGIEL D.Attachment ofAsaia lannensisto materials commonly used in beverage industry[J].Food Control,2013,32(2):537-542.
[54]FAVIA G,RICCI I,MARZORATI M,et al.Transgenesis and the management of vector-borne disease[M].New York:Springer,2008.
[55]SAMADDAR N,PAUL A,CHAKRAVORTY S,et al.Nitrogen fixation inAsaiasp.[J].Curr Microbiol,2011,63(2):226-231.
[56]KUMAGAI A,MIZUNO M,KATO N,et al.Ultrafine cellulose fibers produced byAsaia bogorensis,an acetic acid bacterium[J].Biomacromolecules,2011,12(7):2815-2821.
[57]YAMADA Y,HOSHINO K,ISHIKAWA T.The phylogeny of acetic acid bacteria based on the partial sequences of 16S ribosomal RNA:the elevation of the subgenusGluconoacetobacterto the generic level[J].Biosci Biotech Biochem,1997,61(8):1244-1251.
[58]GOSSELé F,SWINGS J.Identification ofAcetobacter liquefaciensas causal agent of pink-disease of pineapple fruit[J].J Phytopathol,1986,116(2):167-175.
[59]SO K O,WONG P K,CHAN K Y.Decolorization and biodegradation of methyl red byAcetobacter liquefaciens[J].Environ Toxicol Water Qual,1990,5(3):221-235.
[60]SHINJOH M,SUGISAWA T,MASUDA S,et al.Efficient conversion of l-sorbosone to 2-keto-l-gulonic acid byAcetobacter liquefaciensstrains[J].Journal of Fermentation and Bioengineering,1994,78(6):476-478.
[61]SHRIDHAR B S.Review:nitrogen fixing microorganisms[J].Int J Microbiol Res,2012,3(1):46-52.
[62]FRANKE I H,FEGAN M,HAYWARD C,et al.Description ofGluconacetobacter saccharisp.nov.,a new species of acetic acid bacterium isolated from the leaf sheath of sugar cane and from the pink sugar-cane mealy bug[J].Int J Syst Evol Micr,1999,49(4):1681-1693.
[63]FUENTES-RAMíREZ L E,BUSTILLOS-CRISTALES R,TAPIAHERNáNDEZ A,et al.Novel nitrogen-fixing acetic acid bacteria,Gluconacetobacter johannaesp.nov.andGluconacetobacter azotocaptanssp.nov.,associated with coffee plants[J].Int J Syst Evol Micr,2001,51(4):1305-1314.
[64]SCHüLLER G,HERTEL C,HAMMES W P.Gluconacetobacter entaniisp.nov.,isolated from submerged high-acid industrial vinegar fermentations[J].Int J Syst Evol Micr,2000,50(6):2013-2020.
[65]HI LAN VU H,YUKPHAN P,CHAIPITAKCHONLATARN W,et al.Nguyenibacter vanlangensisgen.nov.,sp.nov.,an unusual acetic acid bacterium in the α-Proteobacteria[J].J Gen Appl Microbiol,2013,59(2):153-166.
[66]YAMADA Y.Transfer ofGluconacetobacter kakiaceti,Gluconacetobacter medellinensisandGluconacetobacter maltacetito the genusKomagataeibacterasKomagataeibacter kakiaceticomb.nov.,Komagataeibacter medellinensiscomb.nov.andKomagataeibacter maltaceticomb.nov.[J].Int J Syst Evol Micr,2014,64(5):1670-1672.
[67]YAMADA Y.Transfer ofAcetobacter oboediensSokollet andAcetobacter intermediusBoesch to the genusGluconacetobacterasGluconacetobacter oboedienscomb.Nov.andGluconacetobacter intermediuscomb.nov[J].Int J Syst Evol Micr,2000,50(6):2225-2227.
[68]YAMADA Y,YUKPHAN P,VU H T L,et al.Subdivision of the genusGluconacetobacterYamada,Hoshino and Ishikawa 1998:the proposal ofKomagatabactergen.nov.,for strains accommodated to theGluconacetobacter xylinusgroup in the α-Proteobacteria[J].Annal Microbiol,2012,62(2):849-859.
[69]YAMADA Y,AKITA M,KODA T,et al.Elevation ofAcetobacter acetisubsp.liquefacienstoAcetobacter liquefacienssp.nov.comprising the peritrichously flagellated intermediate in acetic acid bacteria[J].J Gen Appl Microbiol,1983,29(5):327-333.
猜你喜歡
西北民族大學(xué)學(xué)報(自然科學(xué)版)(2021年4期)2021-12-29 02:54:24
數(shù)學(xué)小靈通(1-2年級)(2021年4期)2021-06-09 06:25:56
大眾健康(2021年6期)2021-06-08 19:30:06
小聰仔(科普版)(2020年12期)2021-01-18 09:16:52
東方少年·布老虎畫刊(2020年4期)2020-06-08 15:48:10
學(xué)生天地(2019年32期)2019-08-25 08:55:22
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年4期)2019-05-20 10:06:32
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2018年6期)2018-06-26 08:36:06
小天使·一年級語數(shù)英綜合(2017年11期)2017-12-05 18:49:56
初中生世界·七年級(2017年9期)2017-10-13 22:27:46
