一種印度產紫菜(Pyropia radi)的生物學特性與部分生活史*
2014-04-16 01:07:15張巖巖嚴興洪
海洋與湖沼 2014年1期
張巖巖 嚴興洪
?
一種印度產紫菜()的生物學特性與部分生活史*
張巖巖 嚴興洪①
(上海海洋大學水產與生命學院 上海 201306)
為積累紫菜的新種質, 本文對一種印度產紫菜—的生活史進行了研究, 結果發現: 該紫菜的葉狀體多為披針形, 基部為楔形、呈褐綠色, 梢部呈紅紫褐色; 藻體由單層細胞構成, 極少含兩層細胞; 一個營養細胞中含1—2個星狀色素體, 多數只含一個; 藻體的中部邊緣具有稀疏的鋸齒狀突起, 基部的齒狀突起較密, 一般的突起含有一至十多個細胞; 成熟前的藻體厚度: 基部為(51.5±4.23)μm, 梢部為(29.4±2.31)μm, 平均厚度41.2μm, 成熟后的藻體逐漸加厚; 成熟前的藻體長度5—10cm、寬度0.5—1cm, 成熟后, 藻體長度可達(45±8.7)cm、寬度為(5±1.21)cm; 藻體的表觀性別多數為雌雄異體, 少數為雌雄同體, 雌雄同體的個體其雌雄生殖細胞分別呈塊狀直線型分布于藻體; 果孢子囊具有16或32個果孢子, 分裂式為♀A2B2C4或♀A4B2C4, 成熟的精子囊器具有64個精子囊, 分裂式為♂A4B4C4; 葉狀體營養細胞和精子囊的核型為單倍, 染色體數目為5個(=5), 絲狀體的營養細胞和膨大細胞的核型為雙倍, 染色體數目為10個(2=10)。檢測該紫菜的5.8S rDNA-ITS區序列發現, 其5.8S保守區的160個堿基序列與壇紫菜相似度為100%。該紫菜的形態結構、染色體數目、性別以及5.8S rDNA的保守序列等結果與壇紫菜極為相似, 暗示它與壇紫菜的親緣關系很近, 但它們是否為同一個紫菜物種還有待深入研究。
紫菜(), 生活史, 絲狀體, 葉狀體, 染色體
紫菜屬()(Sutherland, 2011)在系統分類學上隸屬紅藻門(Rhodophyta), 原紅藻綱(Proto-flo-rideae), 紅毛菜目(Bangiales), 紅毛菜科(Bangiaceae), 廣泛分布在寒溫帶的潮間帶和潮下帶水域(Yoshida, 1997)。紫菜富含蛋白質、維生素、礦物質和食用纖維, 其中, 蛋白質的含量為14.9%—56.1%, 維生素C含量高于橙子(Noda, 1975), 因此被人類食用的歷史很長。此外, 紫菜還含有大約17種游離氨基酸, 其中的牛磺酸具有調控人體血液中膽固醇含量的作用, 具有較高的營養和藥用價值(Sahoo, 2006), 年產值超過18億美元(Yarish, 1999)。紫菜屬重要的商業價值依賴于不斷將新品種和新的培養技術用于栽培生產(Pereira, 2010)。
至今, 全球發現的紫菜種已超過133種(Yoshida, 1997), 但自二十世紀中葉開始, 紫菜生活史的研究主要集中于溫帶種的生活史, 很少涉及亞熱帶和熱帶的紫菜種(López-Vivas, 2011)。1881年, Murray首次報道了生長在印度半島海域的紫菜種, 隨后, 在印度海域陸續發現了其它8個紫菜種, 對印度紫菜的分類學作了初步研究, 但對它們的生活史和相關應用性研究涉及較少(Kumar, 1997, 2005)。
紫菜傳統分類的主要鑒定依據是藻體的形態結構, 包括外形、長度、寬度、厚度、顏色、生殖方式、生殖細胞的分布和分裂式等(張德瑞等, 1960; 曾呈奎等, 1978; 王素娟等, 1980; 鄭寶福, 1981, 1988; 潘國瑛等, 1982; 章景榮等, 1993)。但是, 單憑藻體的形態與結構特征, 許多紫菜物種往往難以被正確鑒定, 于是, 藻類學家對紫菜的染色體數目等細胞學特征進行了研究(戴繼勛, 1999)。近年來, 隨著分子生物學的快速發展, DNA分子標記也逐漸被用于紫菜的系統分類、親緣關系的鑒定以及分子輔助育種(湯曉榮等, 2006)。趙玲敏等(2009)認為5.8S rDNA-ITS區的高度保守區和高變區交替排列的形式可以作為壇紫菜種質鑒定以及系統進化分析。
本文從藻體大小、形態、繁殖類型、邊緣刺特征、生殖結構等方面對產自印度的一種紫菜—的生活史進行了詳細研究, 旨在積累新的紫菜種質。
1 材料與方法
1.1 紫菜來源與保存
采自印度半島, 其自由絲狀體由印度藻類學家(Reddy博士)贈送, 并以自由絲狀體的形式被保存于實驗室內, 保存條件: 溫度為18°C, 光密度為5μmol photons/(m2·s), 光周期為14L:10D。
1.2 絲狀體和葉狀體的培養與觀察
取適量的自由絲狀體, 加入一定量的MES培養液(王素娟等, 1986), 用粉碎機切碎后, 制成絲狀體接種液, 將其均勻噴灑于平鋪的貝殼表面, 暗光培養4d后, 在溫度24±1°C、光密度20μmol photons/(m2·s)、光周期10L : 14D的條件下培養10d, 清洗一次貝殼以除去表面多余的自由絲狀體, 換入培養液并繼續培養。當整個貝殼內表面長滿絲狀體, 使顏色變紅后, 把培養溫度提高到28±1°C, 光密度降到10μmol photons/(m2·s), 光周期縮短至8L : 16D, 培養液改用含N : P=1 : 10的滅菌海水。待貝殼表面長出大量膨大藻絲時, 取3個貝殼, 分別放入含100mL培養液和3—5根棉線(長約5cm)的燒杯(250mL)中充氣刺激。當棉線上附著一定量的殼孢子后, 將它們轉移至培養瓶(250mL)中進行充氣培養, 培養溫度為24±1°C、光密度60—80μmol photons/(m2·s)、光周期為10L : 14D。每5天更換一次培養液, 當殼孢子萌發體的長度達3—5cm, 將它們從棉線上摘下, 繼續充氣培養直至成熟。
絲狀體的形態和膨大細胞、葉狀體的邊緣刺等顯微結構用顯微鏡(Nikon Eclipse 90i)觀察與拍照。
藻體厚度的測定: 隨機取培養30d的葉狀體, 用冰凍切片機(Frigocut 2700, Reichert Jung, LEICA公司)分別對梢、中、基部進行橫切, 取完整切片在顯微鏡下用NIS-Elements軟件分別測量各部位的厚度。每個部位測量30個樣品, 取其平均值。
1.3 染色體觀察
1.3.1 材料固定 取一定量生長旺盛的絲狀體在暗周期開始后3h內, 每隔30min用卡諾固定液(無水乙醇 : 冰乙酸=3 : 1)固定一次材料, 在光亮處放置數日, 使藻體的顏色褪至無色, 以備觀察。
1.3.2 材料的染色與觀察 用鑷子取少量褪色為無色的絲狀體, 在蒸餾水中軟化數秒鐘后, 置于載玻片上, 滴加醋酸鐵蘇木精染色液(Wittmann, 1965), 染色約5min后, 在酒精燈火焰上稍微加熱以軟化材料, 然后輕輕壓片, 用吸水紙吸去多余的染色液, 在顯微鏡下觀察。
葉狀體營養細胞、成熟精子囊以及膨大藻絲的固定方法同絲狀體, 其中用于觀察葉狀體營養細胞和精子囊染色體的藻體在固定前需要先切成小塊, 便于后期壓片、觀察和拍照。
1.4 5.8S rDNA-ITS區片段的序列檢測
隨機選擇日齡30d的葉狀體, 提取DNA(用新型植物基因組提取試劑盒, TIANGEN, 北京), 操作步驟同說明書, 根據NCBI上已報道的壇紫菜5.8S rDNA-ITS區的保守序列設計引物, 擴增包含160bp的5.8S保守區域。上、下游引物分別為: 5′-TTT-G-T-CTTATCCGCTTTGT-3′、5′-TTCATCTGGTATGCC-TCCTA-3′。20μL的PCR反應體系中含10× buffer 2μL, DNA模板50ng, 0.2mmol/L dNTP, 0.5μmol/L引物, 1UDNA聚合酶。PCR擴增程序為: 94°C變性5min, 94°C變性45s, 54°C退火30s, 72°C延伸60s, 共進行30個循環, 最后72°C延伸10min。擴增產物由上海美吉生物公司測序, 所得序列通過DNAMAN軟件與NCBI服務器上已發表的(FJ490524)、(JQ416151)、var(AY766366)和(AY766365)的5.8S保守區進行多重序列比對, 檢測它們的序列同源性。
2 結果
2.1 絲狀體和葉狀體的生物學特征
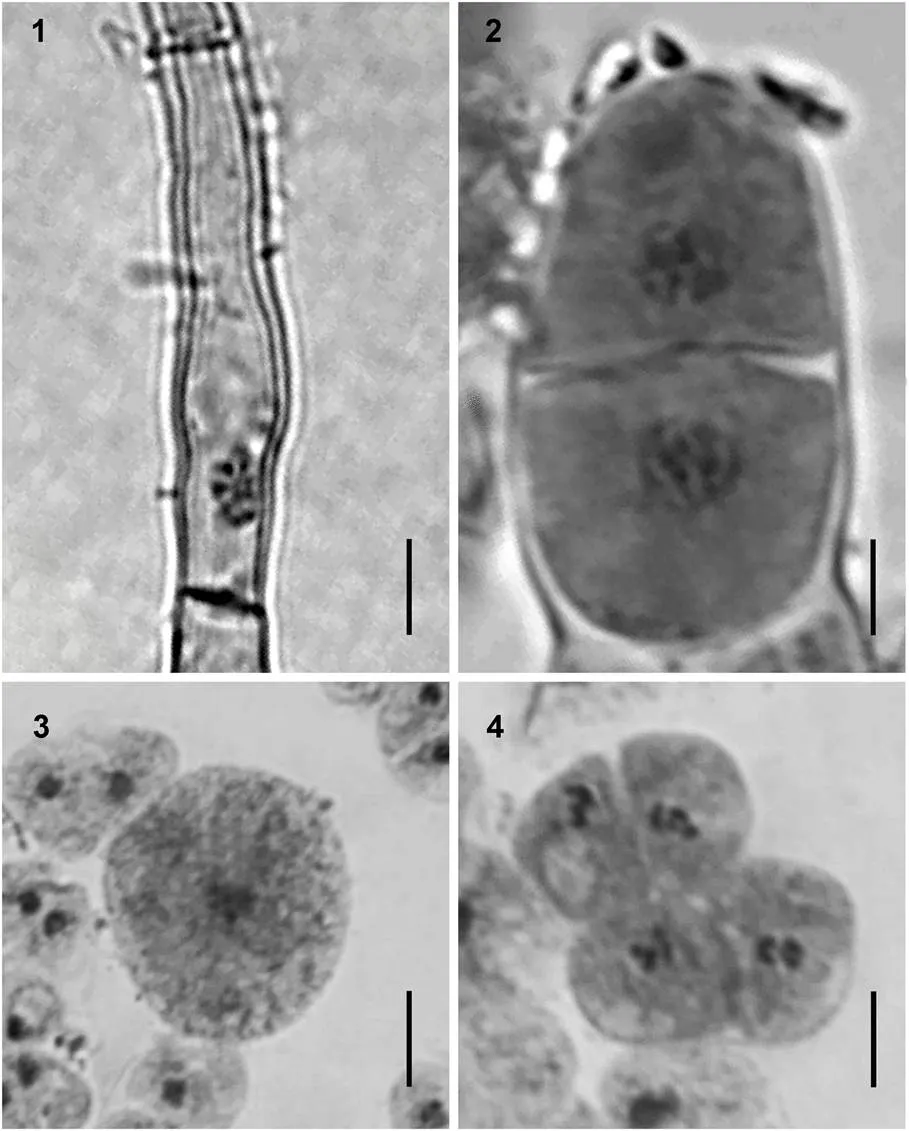
2.1.1 絲狀體的形態學特征 自由絲狀體的顏色為紅紫色(圖版Ⅰ1), 當貝殼表面長滿絲狀體時, 其顏色呈紅紫褐色(圖版Ⅰ2), 在24—30°C范圍內均可發育成膨大藻絲(圖版Ⅰ3)。在此發育過程中, 絲狀體的營養細胞逐漸由細長型變短變粗, 發育成為不定型細胞, 繼續發育成為體積較大的細胞, 形成了膨大藻絲, 當它們成熟時就發育成殼孢子囊枝。在適宜的光照和溫度條件下, 殼孢子囊枝經流水刺激后放散出大量的殼孢子, 并迅速萌發(圖版Ⅰ4), 逐漸長成葉狀體(圖版Ⅰ5)。
2.1.2 葉狀體的形態學特征 藻體呈披針形, 基部為楔形, 基部的根絲細胞圓頭狀, 每個細胞有一個向下延伸的假根絲(圖版Ⅲ9), 構成假根, 葉狀體的梢部較尖(圖版Ⅰ5)。葉狀體的梢部邊緣一般較為平滑, 但中部邊緣存在稀疏的鋸齒狀突起, 基部邊緣的齒狀突起更密(圖版Ⅲ1—3), 齒狀突起的細胞數為一至十多個, 在日齡較大的葉狀體基部邊緣還可見到齒狀突起上又長出細密的次級刺狀突起。藻體一般含單層細胞, 局部含兩層細胞(圖版Ⅲ7)。營養細胞內具有一個明顯的星狀色素體, 少數細胞含有兩個色素體(圖版Ⅲ8)。從梢部到基部, 藻體的厚度逐漸增加, 梢部厚度為(29.4±2.31)μm, 基部為(51.5±4.23)μm, 藻體的梢部、中部和基部三處的厚度平均值為41.2μm。成熟后, 藻體變得更厚。
成熟前的葉狀體體長為5—10cm, 體寬為0.5—1cm。培養至35—40d, 葉狀體開始成熟, 在成熟的早期, 大多數個體的性別為雌雄異體(圖版Ⅰ6a), 極少數為雌雄同體, 雌雄生殖細胞分別呈塊狀直線型分布于藻體(圖版Ⅰ6b)。雄性個體成熟時形成的精子囊區, 其顏色變成淡黃綠色(圖版Ⅱ1), 成熟的單個精子囊器含64個精子囊, 分裂式為♂A4B4C4(圖版Ⅱ2), 成熟后過幾天就可觀察到成熟部位的邊緣放散出大量的精子。雌性個體成熟時, 原來的體細胞顏色變淡, 星狀色素體變得不明顯, 細胞由多角形逐漸變圓, 發育成果胞, 未受精的果胞顏色逐漸變淺甚至變白死亡, 受精后的果胞發育成紅紫色的果孢子囊, 果孢子囊內含有16或32個果孢子, 分裂式為♀A2B2C4或♀A4B2C4(圖版Ⅱ3—4)。部分果孢子在藻體邊緣直接發育成絲狀體, 但大部分脫離后再萌發成絲狀體(圖版Ⅱ5)。雌雄個體充分成熟的梢部和邊緣便開始脫落, 其他未成熟的藻體繼續生長, 當培養至90d, 藻體長度可達(45±8.7)cm, 寬度達(5±1.21)cm。
2.1.3 繁殖方式 大多數葉狀體通過有性生殖即精子與果胞受精后, 發育成果孢子囊, 后者產生的果孢子再萌發成絲狀體, 絲狀體成熟后再釋放殼孢子萌發成葉狀體, 完成生活史。但在單株培養雌性葉狀體時發現, 雌性葉狀體未經受精, 梢部的正常營養細胞逐漸退色, 由褐紅色變成褐綠色, 隨后細胞間隙增大, 藻膠層和細胞壁融化, 細胞以原生質體形式流出, 流出后的細胞迅速變圓并逐漸再生出細胞壁, 細胞中色素體顏色也逐漸變紅, 數小時后, 這些游離出來的細胞陸續萌發成絲狀體(圖版Ⅱ6), 后者產生的殼孢子也可以萌發成葉狀體, 而且葉狀體全部為雌性的, 該種生殖方式被認為是紫菜的單性生殖(嚴興洪等, 2007)。
2.2 染色體數目的觀察結果
熄燈后約2—3h所固定的材料, 其染色體的觀察效果較好, 可看到較清晰的中期染色體。絲狀體的營養細胞和膨大細胞的染色體數為10個, 是雙倍的(2=10)(圖版Ⅳ1—2), 而葉狀體的營養細胞和精子囊細胞的染色體數目均為5個, 是單倍的(=5)(圖版Ⅳ3—4)。在顯微鏡下對1308個精子囊細胞的染色體數目進行了統計, 其中含5條染色體的細胞數占58.9%, 部分處于分裂后期, 未完全分裂成兩個細胞時可以看到10條染色體, 所占比例為28.3%。出現其他條數的原因可能是在壓片過程中的擠壓導致染色體丟失、堆疊, 或計數的誤差。
2.3 葉狀體的5.8S rDNA-ITS區片段的序列分析結果
經PCR擴增, 在4個和3個壇紫菜的葉狀體中均擴增出一條大小均一的DNA 片段, 約500bp(圖1), 與引物設計的目標片段大小一致。
測序結果表明, 擴增片段總長度為477bp, 包含三個部分: ITS1區和ITS2區的部分序列及5.8S區的完整序列。5.8S區片段長度為160bp, 兩端分別有一段polyA和polyT序列。將其與NCBI上已公布的采自不同地區的壇紫菜5.8S保守區序列比對, 發現序列完全一致(圖2), 相似度達到100%, 與、var的5.8S區序列比對, 結果顯示同相似度為95%, 與var和相似度均僅為91%(圖3)。
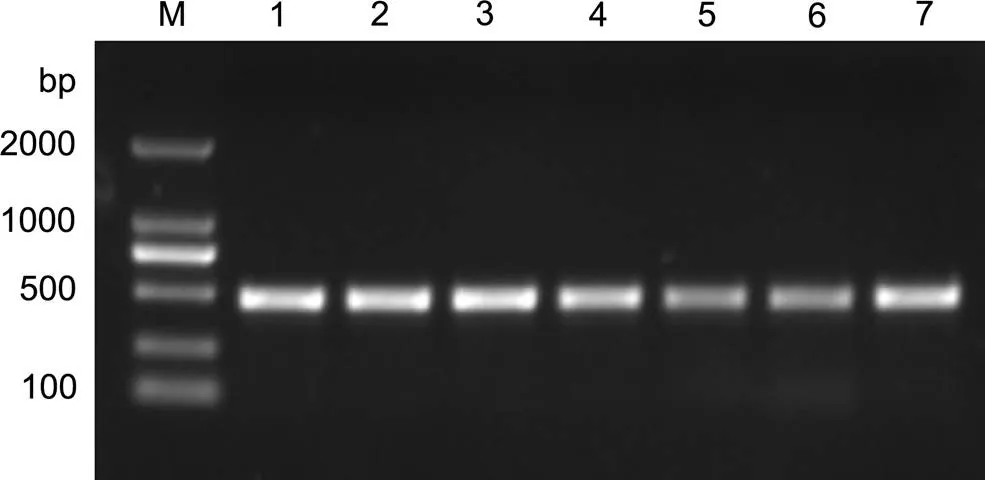
圖1 Pyropia radi和壇紫菜的ITS1-5.8S-ITS2區的PCR擴增產物電泳圖
M: Marker; 泳道1—4:; 泳道5—7: 壇紫菜

圖2 Pyropia radi葉狀體的5.8S rDNA-ITS區測序結果
下劃線部分為上下游引物序列, 陰影部分為5.8S區保守序列
3 討論
紫菜的生活史如圖4所示, 該種紫菜可以通過有性生殖完成生活史過程, 也可以通過單性生殖完成生活史, 但是在整個培養過程中, 葉狀體階段未觀察到葉狀體直接釋放單孢子發育成葉狀體的過程, 即該種紫菜不具有無性生殖行為。

圖3 幾種紫菜5.8S 區的序列比對結果
與壇紫菜之間存在許多相似之處: 藻體的顏色呈暗綠略帶褐紅色, 基部較綠, 梢部較紅; 藻體的外形多呈披針形, 大部分由單層細胞構成, 部分為兩層; 細胞內含星狀色素體一個或兩個; 藻體邊緣從基部到頭部, 齒狀突起由密集逐漸變得稀疏, 突起的細胞數由十幾個逐漸減少至1—2個; 營養細胞和精子囊細胞的染色體數目均為5條(=5), 而絲狀體細胞的染色體均為10條(2=10), 這與壇紫菜(嚴興洪等, 2008)相同; 兩種紫菜的葉狀體性別也基本一致, 即表觀性別在葉狀體的成熟早期以雌雄異體為主, 少數為雌雄同體, 隨著葉狀體進入成熟后期, 雌雄同體的比例增加; 兩種紫菜均可以通過有性生殖產生絲狀體, 也可由葉狀體的單性生殖產生可育的絲狀體(嚴興洪等, 2007)。果孢子囊內的果孢子數目和壇紫菜一樣, 均為16或32個。此外,與壇紫菜相比, 無論是葉狀體還是絲狀體, 其生長發育的適宜生態條件如溫度, 光周期等均非常相似。
與壇紫菜之間也存在不同之處:在成熟前的藻體較小, 成熟后藻體大于壇紫菜。一般藻體成熟前的的體長為5—10cm, 而壇紫菜的體長為12—18cm; 成熟后的藻體會繼續生長, 雖然兩者都存在成熟部分的掉落, 但的未成熟的中基部生長速度快于壇紫菜, 藻體的體長大于壇紫菜, 培養90d,的體長為45cm左右, 而壇紫菜為28cm左右;的藻體比壇紫菜更薄,葉狀體中部厚度一般為27—37μm, 壇紫菜則為65—95μm(張德瑞等, 1960);藻體的成熟結構也存在一些差異,的精子囊數目少于壇紫菜,成熟的精子囊器只含有64個精子囊, 分裂式為♂A4B4C4, 而成熟的壇紫菜精子囊器含有128或256個精子囊, 分裂式為♂A4B4C8或♂A4B4C16。上述差異是實驗觀察誤差還是兩者真實存在的差異還有待進一步驗證。
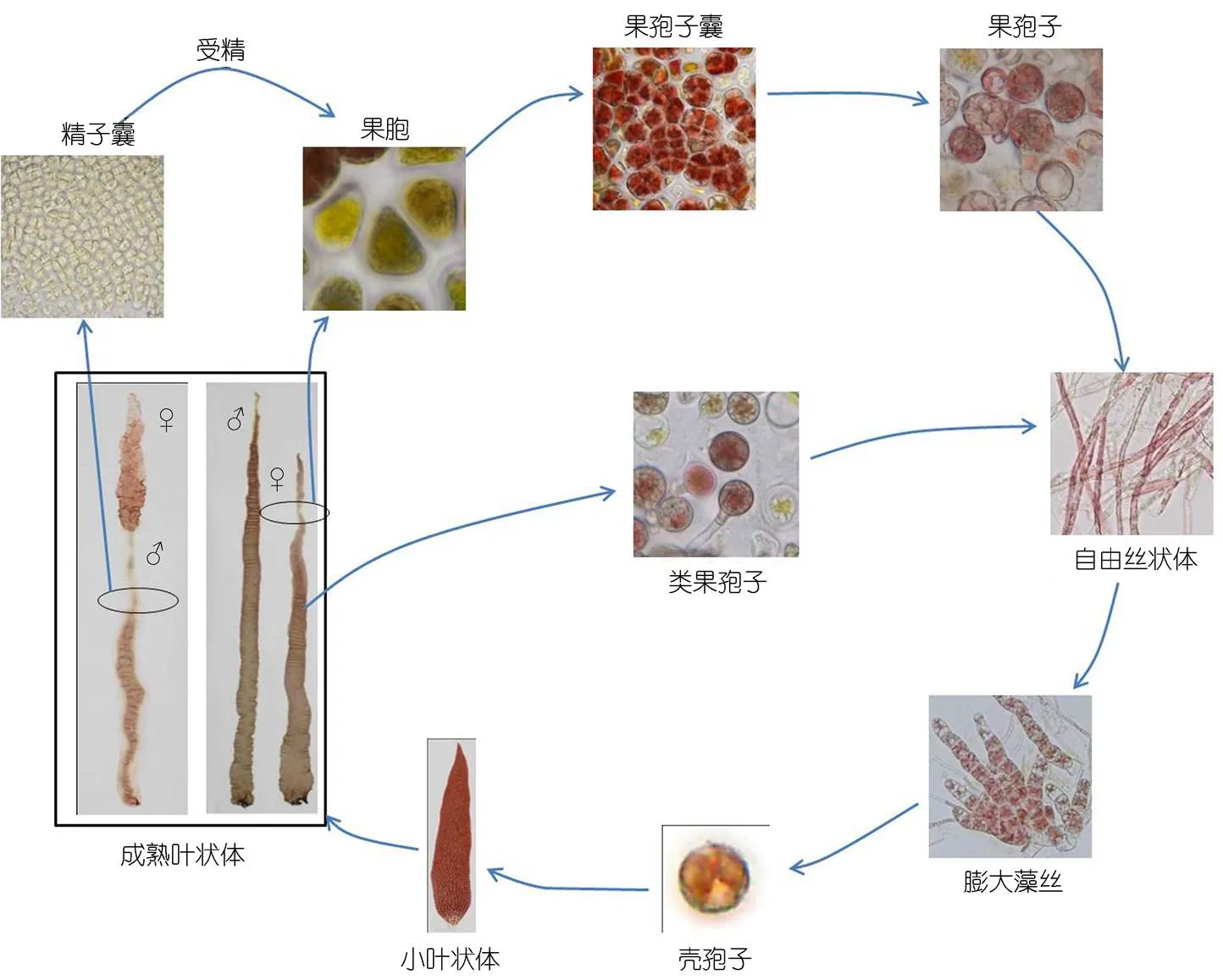
圖4 Pyropia radi的部分生活史示意圖
趙玲敏等(2009)對10個種質材料的野生壇紫菜5.8S rDNA-ITS區進行PCR擴增和測序, 結果表明它們的5.8S區序列完全一致, 但與其他紫菜種的5.8S區序列有很大差異, 序列同源性在79.7%—95%之間。因此5.8S rDNA-ITS區的高度保守區和高變區交替排列的形式可以作為壇紫菜種質鑒定及系統進化分析的強有力工具。從擴增出的5.8S區序列與NCBI上已公布的采自不同地區的20個壇紫菜5.8S區序列相似度達到100%, 這表明在進化上與壇紫菜的親緣關系很近。但在本研究中, 比對的序列有限, 不能全面反應出與壇紫菜是否屬于同一個種。
從地理位置來講,與壇紫菜相隔很遠, 但兩者的生物學、細胞學和分子生物學的諸多特征極為一致, 搞清楚兩者到底是否屬于同一個紫菜物種具有一定的學術價值。
王素娟, 張小平, 徐志東等, 1986. 壇紫菜營養細胞和原生質體培養的研究Ⅰ. 海洋與湖沼, 17(3): 217—221
王素娟, 章景榮, 1980. 紫菜一新種——單孢紫菜的研究. 海洋與湖沼, 11(2): 141—152
湯曉榮, 潘光華, 許東海, 2006. 紫菜分子遺傳學研究進展. 中國海洋大學學報, 36(5): 687—692
嚴興洪, 李琳, 陳俊華等, 2007. 壇紫菜的單性生殖與遺傳純系分離. 高技術通訊, 17(2): 205—210
嚴興洪, 何亮華, 黃健等, 2008. 壇紫菜的細胞學觀察. 水產學報, 32(1): 131—137
張德瑞, 鄭寶福, 1960. 福建紫菜一新種: 壇紫菜. 植物學報, 9(1): 32—36
鄭寶福, 1981. 紫菜一新種——少精紫菜. 海洋與湖沼, 12(5): 447—453
鄭寶福, 1988. 新種紫菜——青島紫菜的描述. 海洋與湖沼, 19(5): 419—424
趙玲敏, 謝潮添, 陳昌生等, 2009. 5.8S rDNA-ITS區片段的序列分析在壇紫菜種質鑒定中的應用. 水產學報, 33(6): 940—948
章景榮, 王素娟, 1993. 紫菜一新種——福建紫菜的研究. 海洋與湖沼, 24(4): 356—359
曾呈奎, 張德瑞, 1978. 中國兩種新紫菜. 海洋與湖沼, 9(1): 76—83
潘國瑛, 王永川, 1982. 我國紫菜屬一新種——多枝紫菜. 海洋與湖沼, 13(6): 543—548
戴繼勛, 沈頌東, 1999. 紫菜的細胞遺傳學研究現狀及展望. 青島海洋大學學報, 29(4): 637—642
Kumar C A, Rao P S N, 2005. A new species of(Rhodophyta, Bangiales) from the Malvan coast of Maharashtra (India). Feddes Repertorium, 116(3—4): 222— 225
Kumar C A, Panikkar M V N, 1997. Indian species of(Rhodophyceae, Bangiales). Feddes Repertorium, 108(5—6): 419—423
López-Vivas J M, Pacheco-Ruiz I, Riosmena-Rodríguez R, 2011. Life history ofDawson (Bangiales, Rhodophyta) from the Gulf of California, México., 50(5): 520—529
Murray J A, 1881. Plants and Drugs Sind. Cambridge University Press, London, UK: 219
Noda H, Horiguchi Y, 1975. Studies on the flavor substances of “Nori”, the dried laver, 1. Dimethyl sulfide and dimethyl-β-propiothetin. Bulletin of the Japanese Society of Scientific Fisheries, 41(4): 481—486
Pereira R, Yarish C, 2010. The role ofin sustainable culture systems: physiology and applications. In:. Celluar Origin, Life in Extreme Habits and Astrobiology, 15: 339—354
Sahoo D, Baweja P, Kushwah N, 2006. Developmental studies in: A high-temperature resistant species from the Indian Coast. Journal of Applied Phycology, 18(3—5): 279—286
Sutherland J E, Lindstrom S C, Nelson WA, 2011. A new look at an ancient order: Generic revision of the Bangiales (Rhodophyta). Journal of Phycology,47(5): 1131—1151
Wittmann W, 1965. Acto-iron-heamatoxylin-chloral hydrate for chromosome staining. Stain Technology, 40: 161—164
Yarish C, Chopin T, Wilkes R, 1999. Domestication of nori for Northeast America: the Asian experience. Bulletin of the Aquaculture Association of Canada, (1): 11—17
Yoshida T, Notoya M, Kikuchi N, 1997. Catalogue of species ofin the world, with special reference to the type locality and bibliography. National History Research Special Issue, 3: 5—18
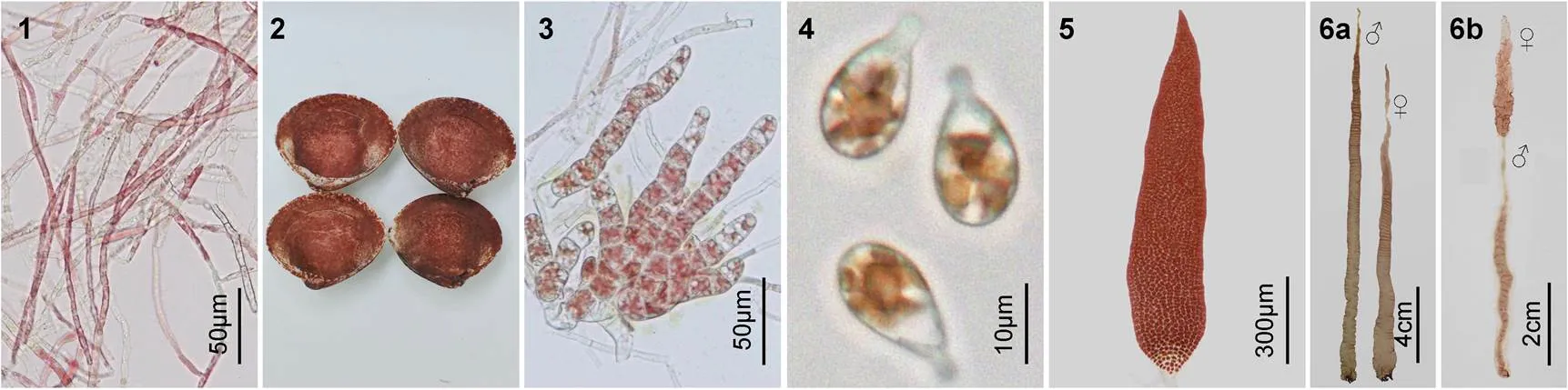
圖版Ⅰ生活史的不同階段
PlateⅠ Photomicrographs of different phases of life-cycle in
1. 自由絲狀體; 2. 貝殼絲狀體; 3. 膨大藻絲; 4. 殼孢子萌發體; 5. 小葉狀體; 6. 成熟葉狀體: 6a. 雌雄異體, 6b. 雌雄同體。

圖版Ⅱ的生殖細胞和繁殖方式
PlateⅡ Photomicrographs of spermatangiopore, carposporangium, and reproduction of
1, 2. 精子囊的表面觀和切面觀, 含精子囊64個, 分列式為♂A4B4C4; 3, 4. 果孢子囊表面觀及切面觀, 果孢子囊為16個或32個, 分列式為♀A2B2C4或♀A4B2C4; 5. 果孢子發育成絲狀體; 6. 雌性葉狀體邊緣的細胞游離出來進行單性生殖, 產生絲狀體。
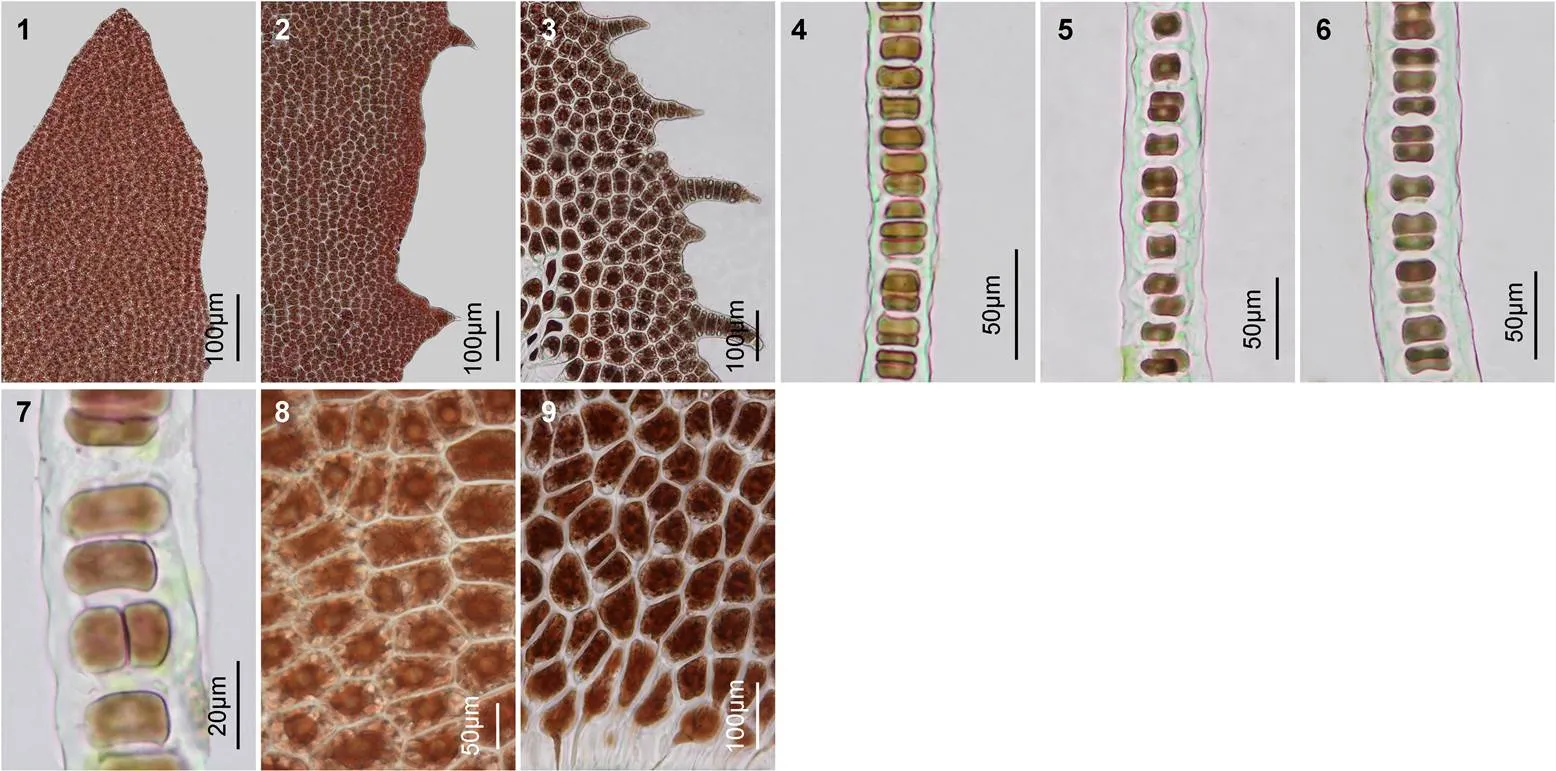
圖版Ⅲ葉狀體的邊緣刺、切面觀和細胞形態結構
PlateⅢ Photomicrographs of margin and spines, section and surface views of the tip, middle and base of blades in
1—3. 葉狀體的梢、中、基部的邊緣和刺狀突起; 4—6. 葉狀體梢、中、基部的橫切面觀; 7. 局部葉狀體含兩層細胞; 8. 部分細胞內含有兩個星狀色素體; 9. 葉狀體基部的假根絲細胞, 具有圓頭狀向下延伸的假根絲。
圖版Ⅳ不同生活史階段的染色體微觀照片
PlateⅣ Chromosome microphotographs in different phases of life-cycle of
1.絲狀體營養細胞含10條染色體, 2=10; 2. 膨大藻絲細胞含10條染色體, 2=10; 3. 葉狀體的體細胞含5條染色體,=5; 4. 精子囊細胞含5條染色體,=5。標尺均為5μm
INDIA-ENDEMIC(BANGIALES, RHODOPHYTA): BIOLOGY AND LIFE HISTRY
ZHANG Yan-Yan1, YAN Xing-Hong1
(College of Fisheries and Life science, Shanghai Ocean University, Shanghai 201306, China)
To sustain the germplasm resources of, biological characteristics and life history offrom India were studied. We found that the gametophytic blades of this species are membranous, lanceolate, showing wedge-shape and dark green in the base, but slightly reddish brown in the tip. The blades are most monostromatic, except for a little part consisting of two layers of cells. There are mostly one or sometimes two stellate chloroplasts in the vegetative cell of the blade. The spines in the margins of the middle are sparse but rich at the base of the blades, consisting of one or more than 10 cells in a spine, which become bigger and increases in the number when aged. Before maturation, the base thickness of the blade is (51.5±4.23), the tip (29.4±2.31) and the mean 41.2μm; the blades are 5-10cm in length and 0.5-1cm in width. However, they could grow up to (45±8.7) cm long at maturation. Most blades are dioecious and few monoecious. The sectors of male and female sexual cells usually distribute linearly in the monoecious blades after matured. A mature carposporangium contains 16 or 32 carpospores, showing cell division as ♀A2B2C4or ♀A4B2C4. A mature spermatangiophore contains 64 spermatangia, showing cell division as ♂A4B4C4. The chromosome counts of vegetative cells and spermatangiain the blades are 5 (=5), and the chromosome number in both vegetative cells and conchosporangial branch of conchocelis are 10 (2=10). One-hundred and sixty base-sequences in the 5.8S rDNA-ITS region of this species are the same to those of. These results indicate that the genetic relationship between this species andis very close. However, whether they are the same species needs more works in the future.
; life-cycle; conchocelis; gametophytic blade; chromosome
Q94
10.11693/hyhz20130400030
* 國家高科技研究發展計劃(863計劃)資助項目, 2012AA100811號; 國家自然科學基金資助項目, 31072208號; 農業部公益性專項, 200903030號; 國家海洋局公益專項, 201105008號, 201105023號。張巖巖, 碩士生, E-mail: zhangyy870227@126.com
嚴興洪, 教授, 博士生導師, E-mail: xhyan@shou.edu.cn
2013-04-19,
2013-05-19
