作物遺傳育種研究進展Ⅰ.作物馴化
2014-08-07 08:50:24
作物研究 2014年1期
關鍵詞:植物
(湖南農業大學,長沙 410128)
任何作物都經歷著由野生植物采收、栽培、馴化、育種的逐步發展過程。馴化是把野生植物培育成栽培植物的過程。
植物由野生狀態變為栽培狀態,所處的生態條件發生了顯著改變。生態條件的變化自動地導致選擇壓的顯著改變,在野生條件下重要的性狀,在新的環境里喪失了優勢,在新的環境里會自動選擇適應的新性狀。因此,在栽培條件下,人為有意識選擇和無意識選擇均在發揮作用。有意識選擇包括:(1)不落粒,喪失自然擴散機制;(2)種子均一,快速萌發;(3)繁殖體(種子)變大(大粒);(4)同時成熟;(5)喪失機械保護機制(如刺毛);(6)果實、種子顏色改變,導致營養價值提高,出苗和苗期長勢弱;(7)喪失苦味或有毒物質。無意識選擇(自動選擇)是沒有人為干擾的作用,主要表現在三個方面:(1)繁殖方式:種子繁殖還是營養繁殖;(2)收獲的器官:繁殖器官還是營養器官;(3)耕種收獲的影響(子粒作物形成馴化癥狀群)和園藝環境對果樹的影響[1]。
動植物馴化是人類第一次產業革命開始的標志。第一批植物馴化距今已有12 000年左右,但是這個過程從未中斷,至今仍不斷發現野生植物的可利用價值,被馴化為新作物[2,3]。了解作物的馴化過程和機制,不僅能揭示作物的進化過程,而且有助于加快新作物的馴化。由于家系QTL定位、自然群體關聯分析、基因組重測序和選擇簽名(selection signature)篩選、反向遺傳學和候選基因分析方法等長足進步[4],近十年有關作物馴化的研究成為了一個熱點,并取得了顯著進展。
1 作物馴化的模式和遺傳效應
作物馴化是在植物栽培之后發生的,是一個逐步的不斷試錯的過程,是一個漫長的漸進過程,可能經歷數百代。Meyer & Purugganan[5]把這個過程分為4個時期,即:馴化起始期(第1階段)、有利等位基因頻率提高期(第2階段)、適應當地新環境的栽培群體形成期(第3階段)和有計劃育種期(第4階段)。其中只有第1階段屬于馴化(domestication),這一過程大約持續了2 000年,使得作物在進化上不同于野生祖先種,而第2到第4階段屬于多樣化(diversification)階段,使得作物品種在產量、適應性和品質上得以顯著改良(表1)。但也有人認為這種分類是模糊的[6]。
不同作物的馴化可能經歷不同路徑。傳統認為作物是單次馴化的結果,野生種與馴化種有大的生殖隔離。但隨著考古研究的深入和分子數據的增加,對更多作物的進化史有了更多了解,發現作物馴化還存在其他模式。原先認為是單次馴化的作物如水稻、玉米、小麥、珍珠粟等作物在馴化過程中實際上導入了野生親緣種的基因;大麥、高粱等作物是由一個野生種在不同地方、不同時間點經過多次馴化而來;花生、草莓等作物是由種間雜交后營養繁殖馴化而來[5]。
人們在馴化作物時,一般是選擇當地原產植物,且只是利用個別或少部分野生植株進行馴化,因而造成遺傳多樣性降低(馴化瓶頸效應,圖1)和遺傳漂移發生,如玉米adh1和glb1兩個中性位點分別只保留玉蜀黍86%和60%的遺傳多樣性。再如,向日葵在馴化和改良時492個位點有36個位點成了選擇目標,并且像玉米一樣,這些基因明顯聚集在一塊。這也說明馴化只改變了作物基因組的某些熱點區域(genetic hotspot),在一定程度上是可以預測的。
在作物馴化過程中選擇了對人類有利的性狀。在這個選擇過程中,控制目標性狀的靶標基因經歷了選擇性清除(selective sweep),只保留有利等位基因,與靶標基因緊密連鎖的基因也可能經歷同樣選擇而保留下來,從而產生搭車效應(或叫選擇牽連效應,圖1)。如玉米10號染色體有1.1 Mb、15個以上基因由于選擇喪失了遺傳多樣性,與靶標基因的來源一致。
2 作物馴化的影響因素[7]
(1)交配方式。自交植物更易被馴化,在馴化時多樣性喪失最多。馴化因定向選擇和異交衰退增加自交率,使植物由異交進化為自交,在栽培禾谷類植物中自花授粉植物比例高于異花授粉植物。自交植物多樣性喪失,更容易被馴化。
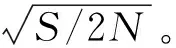
(3)是否出現遺傳瓶頸。在馴化時由于選擇和自交會導致多樣性喪失(遺傳瓶頸效應),馴化后由于基因突變或(通過花粉)基因交流(gene flow)、天然雜交又會產生或重新導入遺傳多樣性。由于突變表型每代可改變0.1%~1%。
(4)選擇強度。
(5)基因交流速率。
(6)遺傳結構(genetic architecture, 少數主效位點)。
(7)靶標基因間的連鎖關系。馴化時由于強烈選擇作用,使目的基因附近10~100 kb范圍受到影響。
作物馴化程度反映在表型分化、種植區域大小、栽培歷史、遺傳改造程度(如種間雜交、多倍化)和育種改良程度等方面。

表1 作物馴化(第1階段)和多樣化(第2到第4階段)過程經常發生的性狀變化[5]
3 作物馴化對性狀的影響
馴化使作物適應栽培環境,生長習性和植株形態等諸多方面發生顯著改變,如蜀黍馴化為玉米,短日照、多分枝變為日中性、單稈;一粒小麥、大麥和水稻等禾本科作物在馴化時先增加籽粒大小和改變形狀,再馴化不落粒性。刺菜薊、菊芋由同一種二倍體植物馴化而來,起源于地中海,栽培上菊芋采用營養繁殖,刺菜薊采用種子繁殖。同一種植物經同時對不同性狀進行選擇形成了2種作物:菊芋主要是大的花序,刺菜薊主要是肉質莖和葉柄。菊芋可能在西西里(Sicily)馴化,但刺菜薊起源于地中海西部,可能在西班牙和法國境內馴化。前者春季、秋季開花,后者只春季開花。植物在馴化過程中發生的變化,稱為馴化群性狀(表1)。
作物馴化的結果,一是使馴化植物喪失了在野外生存的能力,向逐漸適應栽培環境、符合人類需要的方向發展;二是使不同作物產生相同或相似的馴化性狀[6]。
4 作物馴化的分子機制
作物馴化后表型與野生祖先種發生了顯著改變,控制這種表型改變的靶標基因叫做馴化基因。馴化基因應滿足三個條件:(1)其功能已經鑒定并已知其控制的性狀;(2)有證據表明對其實施了正向選擇;(3)至少有一次突變在某次馴化的所有后代中完全或近乎完全固定下來。Meyer & Purugganan[5]統計了60個馴化有關基因,結果只有23個是馴化基因,另有5個既是馴化基因,又是多樣化基因,32個是多樣化基因。在這60個基因中37個編碼轉錄因子基因,14個編碼酶,6個編碼轉運蛋白和泛素連接酶,說明有些基因更可能成為馴化的靶標。轉錄因子基因(如AG基因)能整合上游基因、調控下游基因,一旦突變會導致多個性狀的變化(一因多效)。這類基因往往是馴化起始期(第1階段)的靶標基因[8],如水稻馴化之初改變水稻祖先的野草特征涉及的落粒性、種子休眠和株型等性狀的基因多是轉錄因子基因[9]。如果這種轉錄因子基因(如MYB基因)的表達具有組織特異性,受影響的性狀就會減少到最低[6]。編碼酶的基因只影響代謝途徑的一個反應,其突變只改變一個性狀,在不同作物中容易出現平行突變,如Waxy基因在許多禾谷類作物中發生了突變,形成糯性籽粒。
馴化基因絕大多數是核苷酸突變。在上述60個基因中,38個出現了移碼或提前終止,導致基因沒有功能,26個順式調控區突變,導致基因表達改變,且許多基因在幾個區域發生突變。但也發現染色體水平、基因水平的突變,如高粱SH1基因啟動子發生了序列刪除,玉米Sh1-5.1基因外顯子3后有一段未知序列(易位)、tb1基因順式調控區有轉座子插入、m19基因拷貝數不同,因此可以說作物馴化是建立在自然發生的不同水平、不同類型突變的基礎之上[4]。
5 作物馴化中心
人類在12 000年前的新石器時代即開始作物馴化。據估計有2 500種植物經歷過馴化,250種植物已完全馴化[5]。1940年,Vavilov根據作物遺傳多樣性鑒定出7個作物初生馴化中心,到21世紀又發現南美熱帶雨林、東南亞和北美東部等馴化中心,使世界上主要作物馴化中心增加到10個:中國、南亞、西南亞(即新月沃地)、地中海、埃塞俄比亞、大墨西哥(greater Mexico)、安梯斯山脈(the Andes)、北美東部、新幾內亞和亞馬孫盆地[10]。Purugganan & Fuller[11]將分子遺傳研究結果與考古證據相結合,認為作物馴化中心有13個,即:
1.北美東部 (Chenopodiumberlandieri,Ivaannua,Helianthusannuus, 距今4 500~4 000年);
2.中美洲(Cucurbitapepo, 距今10 000年;Zeamays, 距今9 000~7 000年);
2a.南美北部低地地區(Cucurbitamoschata,Ipomoeabatatas,Phaseolusvulgaris,林木, 距今9 000~8 000年);
3.安第斯山脈中部中海拔地區(Chenopodiumquinoa,Amaranthuscaudatus, 距今5 000年);
3a. 安第斯山脈中北部中、高海拔地區 (Solanumtuberosum,Oxalistuberosa,Chenopodiumpallidicaule, 距今8 000年);
3b. 亞馬孫流域南部低地地區 (Manihotesculenta,Arachishypogaea, 距今8 000年);
3c. 厄瓜多爾和秘魯西北部(Phaseoluslunatus,Canavaliaplagiosperma,Cucurbitaecuadorensis, 距今10 000年; 3c與3、3a和3b中心是否獨立存在疑問);
4. 西非撒哈拉沙漠以南地區(Pennisetumglaucum, 距今4 500年);
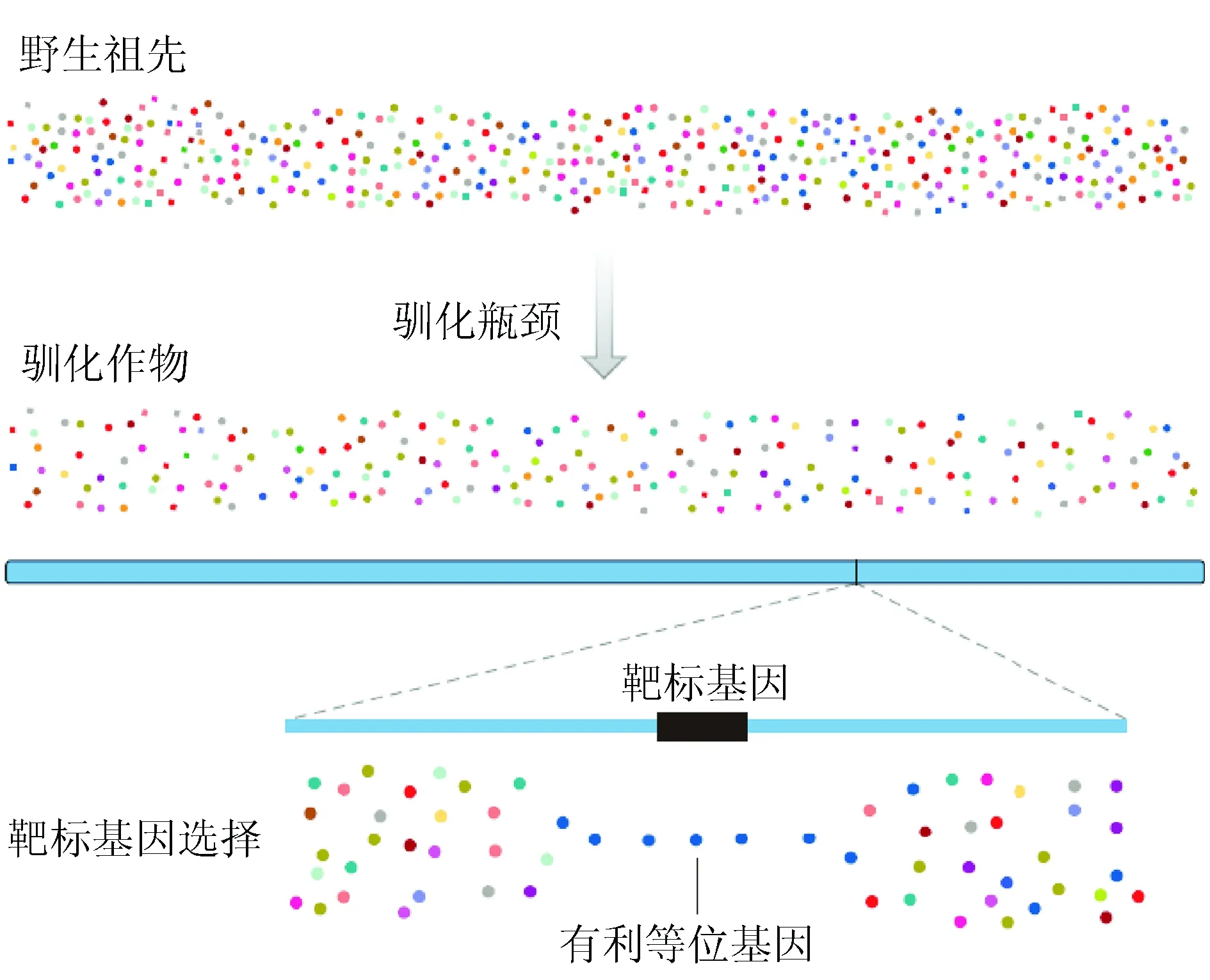
圖1 作物馴化的遺傳效應[4]注:不同顏色的點代表野生祖先和馴化作物群體中整個染色體上基因中性等位基因的多樣性程度。當野生祖先種中只有一部分植株成為馴化作物的先驅時,顯著發生遺傳漂移,預期導致全基因組的遺傳多樣性降低。相反,選擇只是差別性地降低控制受到選擇的性狀的特定基因遺傳多樣性降低。隨著有利等位基因達到較高頻率,靶標基因內及其附近存在的許多遺傳變異將從群體中消除,從而留下選擇的分子痕跡。
4a. 西非草原和林地地區(Vignaunguiculata, 距今3 700年;Digitariaexilis,Oryzaglaberrima, <3 000年);
4b. 西非熱帶雨林地區 (Dioscorearotundata,Elaeisguineensis, 很少記述);
5. 非洲蘇丹東部 (Sorghumbicolor, >4 000年?);
6. 東非高原 (Eragrostistef,Eleusinecoracana, 距今4 000年?) 及低地菜園區(Dioscoreacayenensis,Enseteventricosum, 很少記述);
7. 近東地區(Hordeumvulgare,Triticumspp.,Lensculinaris,Pisumsativum,Cicerarietinum,Viciafaba, 距今13 000~10 000年);
7a. 新月沃地東部(其余Hordeumvulgare);
8a. 印度古吉拉特邦(Panicumsumatrense,Vignamungo, 距今5 000年?);
8b. 印度河上游地區(Panicumsumatrense,Vignaradiata,Vignaaconitifolia, 距今5 000年);
8c. 恒河流域(Oryzasativasubsp.indica, 距今8 500~4 500年);
8d. 印度南部地區(Brachiariaramosa,Vignaradiata,Macrotylomauniflorum, 距今5 000~4 000年);
9. 東喜馬拉雅山脈和云貴高原(Fagopyrumesculentum, 距今5 000年?);
10. 中國北部 (Setariaitalica,Panicummiliaceum, 距今8 000年;Glycinemax, 距今4 500年?);
11. 日本北海道南部地區(Echinochloacrusgalli, 距今4 500年);
12. 中國長江流域(Oryzasativasubsp.japonica, 距今9 000~6 000年); 12a. 中國南部 (Colocasia,Coixlachryma-jobi, 很少記述, 距今4 500年?);
13. 新幾內亞和印度尼西亞蘇拉威斯以東地區(Colocasiaesculenta,Dioscoreaesculenta,Musaacuminata, 距今7 000年)。
[1] Zohary D. Unconscious selection and the evolution of domesticated plants[J]. Econ Bot, 2004, 58:5-10.
[2] Dempewolf H,Rieseberg LH,Cronk QC. Crop domestication in the Compositae: a family-wide trait assessment[J]. Genet Resour Crop Evol, 2008, 55:1141-1157.
[3] Shapter FM, Cross M, Ablett G, et al. High-throughput sequencing and mutagenesis to accelerate the domestication ofMicrolaenastipoidesas a new food crop[J]. PLoS ONE, 2013, (8): e82641.
[4] Olsen KM, Wendel JF. A buntiful hrvest: Genomic insights into crop domestication phenotypes[J]. Annual Review of Plant Biology, 2013,64:47-70.
[5] Meyer RS, Purugganan MD. Evolution of crop species: genetics of domestication and diversification[J]. Nat Rev Gent, 2013, 14:840-852.
[6] Lenser T, Theiβen G. Molecular mechanisms involved in convergent crop domestication[J]. Trends Plant Sci, 2013,18:704-714.
[7] Burger JC, Mark A, Chapman MA, et al. Molecular insights into the evolution of crop plants[J]. Amer J Bot, 2008, 95: 113-122.
[8] Doebley JF, Gaut BS, Smith BD. The molecular genetics of crop domestication[J]. Cell, 2006, 127:1309-1321.
[9] 區樹俊, 汪鴻儒, 儲成才. 亞洲栽培稻主要馴化性狀研究進展[J].遺傳,2012,34:1379-1389.
[10]Smith BD. Eastern North America as an independent center of plant domestication[J]. PNAS, 2006, 103:12223-12228.
[11]Purugganan MD, Fuller DQ. The nature of selection during plant domestication[J]. Nature, 2009, 457:843-848.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
