10株綠僵菌菌株分類地位的多基因系統進化分析
2014-08-10 12:29:50殷幼平王中康劉娟娟廖玉鳳
植物保護 2014年5期
王 萌, 殷幼平, 王中康, 劉娟娟, 廖玉鳳
(重慶大學生命科學學院基因工程研究中心,重慶 400030)
研究報告
10株綠僵菌菌株分類地位的多基因系統進化分析
王 萌, 殷幼平, 王中康*, 劉娟娟, 廖玉鳳
(重慶大學生命科學學院基因工程研究中心,重慶 400030)
為確定10株高效殺蟲綠僵菌菌株的分類地位,通過克隆、測序10株綠僵菌菌株的ef1-α、rpb1、rpb2、β-tubulin部分序列進行多基因系統進化分析,并結合形態特征確定10株綠僵菌菌株的分類地位。結果表明多基因分子鑒定在綠僵菌菌種的劃分上較單基因更為準確,結合多基因系統進化分析結果和形態學特征,將CQMa114、CQMa117、CQMa125、CQMa126、CQMa128歸為大孢綠僵菌,CQMa102歸為蝗綠僵菌,CQMa132、CQMa135歸為貴州綠僵菌,菌株CQMa138、CQMa140與其他綠僵菌親緣關系較遠,有可能為綠僵菌屬的新種或新的分類單位。
綠僵菌; 分類鑒定; 多基因系統進化分析; 形態學
綠僵菌屬(MetarhiziumSorokin)的系統分類地位為菌物界(Fungi),子囊菌門(Ascomycota),糞菌綱(Sordariomycetes),肉座菌目(Hypocreales),麥角菌科(Clavicipitaceae),是一個包括多種重要蟲生真菌的真菌類群[1]。據不完全統計,近年來世界各地有近百余種農作物、林木及衛生害蟲采用綠僵菌進行室內防效測試與評價[2-3]。從應用范圍看,綠僵菌已發展為僅次于白僵菌的殺蟲真菌屬種。殺蝗綠僵菌制劑已用于我國農牧區飛蝗和土蝗的大面積生物防治[4]。綠僵菌屬是麥角菌科的重要昆蟲病原真菌屬,由于具有較強的寄主專化性的大多數綠僵菌種類僅產生形態簡單的無性型,綠僵菌菌種的正確鑒別,對綠僵菌殺蟲制劑的研制和開發有著重要的指導意義。傳統的綠僵菌分類主要根據菌絲體、產孢結構和分生孢子進行分類鑒定。
進入20世紀60年代后,分子生物學方法進入真菌分類鑒定,其中應用最為廣泛的是利用保守的ITS rDNA 序列對真菌進行分類。但由于ITS序列提供的遺傳信息有限,Driver等根據ITS序列將綠僵菌屬劃分為多個變種,其中黃綠綠僵菌、金龜子綠僵菌分別定義為包含種下變種的復合種[5]。Bischoff等通過結合功能基因轉錄延長因子(ef1-α),RNA聚合酶Ⅰ(rpb1),RNA 聚合酶Ⅱ(rpb2)進行多基因系統進化分析,并結合形態學特征證明易碎綠僵菌不屬于黃綠綠僵菌類群[6]。這種多基因系統進化分析的有效性,為綠僵菌屬的分類鑒定提供了一個更加細化和可靠的方法[7]。Bischoff等結合功能基因ef1-α,rpb1,rpb2以及β-tubulin(微管蛋白基因)對綠僵菌屬進行系統進化分析的研究[7]。將Driver 命名的兩個變種上升到種的水平,分別為M.acridum和M.lepidiotae,同時還將前人鑒定的2個變種升到種的地位,重新命名為M.anisopliae,M.majus, 并且發現了2個新種:M.giohosurn和M.robertsii。相對于ITS單一靶標基因對綠僵菌屬的分類鑒別方法,多基因序列分析能夠更明確地確定綠僵菌屬種的分類地位,對其種下變種的細化分類具有明顯的優勢。
近年來本研究室經過廣泛的采集收集,從國內外采集了大量的殺蟲真菌菌株,其中一些種類在形態上差異很小,分類地位難以確定。本研究根據菌株的ef1-α、rpb1、rpb2、β-tubulin部分序列對10株經形態初步鑒定為綠僵菌的高效殺蟲菌株進行多基因系統進化分析,結合形態學特征以確定菌株的科學分類地位。
1 材料與方法
1.1 供試材料
供試菌株:實驗室保存經形態初步鑒定為綠僵菌的菌株10株,菌株信息見表1,菌株培養基:1/4 SDA培養基(蛋白棟2.5 g,葡萄糖10 g,酵母膏0.5 g,瓊脂15~20 g,蒸餾水1 000 mL)為菌株培養基。
1.2 試驗方法
1.2.1 形態學觀察
將各菌株分別接種于1/4 SDA培養基上,28 ℃培養6~8 d,觀察菌落形態、孢子形態、產孢結構等形態特性。
1.2.2 基因組DNA提取
將菌種接種于1/4 SDA液體培養基中,28 ℃振蕩培養5~7 d,收集菌絲。分別以Fermentas 真菌基因組DNA提取試劑盒提取各菌株的基因組DNA。
1.2.3 目的基因的擴增及測序
用于多基因系統進化分析的目的基因包括ef1-α、rpb1、rpb2、β-tubulin,引物序列參見文獻[7]。
PCR反應體系(50 μL)包括5 μL 10×PCR buffer,1 μL dNTP,1.5 μL MgCl2,正反向引物各1 μL,模板2 μL,0.2 μL PlatinumTaq酶(Invitrogen公司)。PCR反應程序為預變性95 ℃5 min,1個循環;95 ℃ 30 s,58 ℃ 30 s,72 ℃ 90 s,35個循環;72 ℃延伸10 min。擴增產物在1.5%瓊脂糖凝膠(含EB)中電泳30 min 后在紫外燈下切下凝膠,以Fermentas凝膠回收試劑盒回收擴增產物,回收產物經T4連接酶連接過夜,轉化trans5α感受態細胞,連接體系為5 μL solutionⅠ,4 μL DNA,1 μL pmd-19 載體。連接過夜后向裝有25 μL感受態細胞的1.5 mL 離心管中加入10 μL 連接產物,冰浴30 min,轉入42 ℃水浴 50 s ,再次冰浴2 min,后向轉化產物中加入800 μL 液體LB培養基,37 ℃培養1 h,將培養物3 500 r/min 離心3 min,棄上清液,留取下部約30 μL菌液用移液槍吹打混勻,將菌液涂平板,38 ℃培養12 h,挑取陽性克隆經PCR驗證后,送北京六合華大基因科技股份有限公司測序。
1.2.4 序列比對及系統進化分析
從GenBank中下載已登錄的綠僵菌ef1-α、rpb1、rpb2、β-tubulin序列信息,與本研究所測的菌株序列一起,用Clustal X 1.83軟件分別對基因序列進行比對分析,以球孢白僵菌為外群,用MEGA4.1 軟件構建菌株單基因和多基因系統進化樹,根據形態和系統進化分析結果確定各菌株的科學分類地位。
2 結果和分析
2.1 形態學特征
2.1.1 菌落形態
將菌株接種在1/4 SDA培養基上,28 ℃培養14 d后記錄菌落形態、產孢情況。觀察分生孢子和產孢細胞即分生孢子梗的特征。菌落形態特征見表1。
表1綠僵菌不同菌株菌落及孢子形態特征
Table1CharacteristicsofcoloniesandconidiaofcollectedMetarhiziumstrains

綠僵菌菌株StrainofMetarhizium菌落形態特征Charactersofmorphology菌落色澤Colorofcolony菌落形態Shapeofcolony基質色澤Colorofbasifixedcolony采集地Collectlocation年份YearCAMa102黃色Yellow茸狀Chopped黃色Yellow重慶Chongqing1990CQMa114橄欖綠Olivegreen絨毛狀Villiform淡黃Lightyellow河南Henan2005CQMa117橄欖綠Olivegreen絨毛狀Villiform淡黃Lightyellow廣西Guangxi2007CQMa125橄欖綠Olivegreen絨毛狀Villiform淡黃Lightyellow河南Henan2007CQMa126橄欖綠Olivegreen絨毛狀Villiform淡黃Lightyellow河南Henan2007CQMa128橄欖綠Olivegreen絨毛狀Villiform淡黃Lightyellow河南Henan2007CQMa132綠色Green茸狀Chopped黃褐色Tawny山東Shandong2009CQMa135墨綠色Blackandgreen茸狀Chopped黃褐色Tawny山東Shandong2009CQMa138綠色Green茸狀Chopped紅褐色Mahogany湖南Hunan2010CQMa140灰綠Greygreen絨毛狀Villiform黑色Black廣西Guangxi2011
2.1.2 顯微形態
將各菌株通過插片培養的方法接種于1/4 SDA培養基,6 d后取下玻片,在顯微鏡下觀察分生孢子和產孢結構(圖1、圖2),分生孢子大小及形狀見表2。其中菌株CQMa102的分生孢子呈卵形,大小與其他綠僵菌差異較大,分生孢子梗聚集為輪生體,CQMa114、CQMa117、CQMa125、CQMa126、CQMa128的分生孢子皆為長橢圓形,分生孢子大小無明顯差異,分生孢子梗緊密排列,呈帚狀分枝。CQMa132、CQMa135的分生孢子呈柱狀,大小7.5~11.8 μm,分生孢子梗聚集,每個分生孢子梗上有多個瓶梗。CQMa138、CQMa140分生孢子呈桿狀,分生孢子梗只有1~2個瓶梗。
表2綠僵菌菌株的分生孢子形態
Table2ConidialfeaturesofMetarhiziumspecies
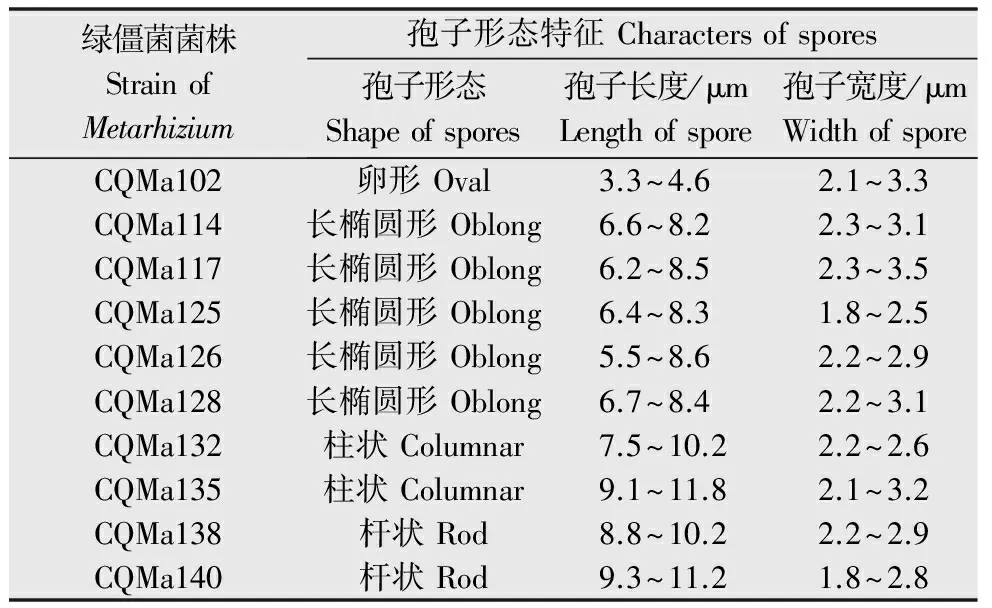
綠僵菌菌株StrainofMetarhizium孢子形態特征Charactersofspores孢子形態Shapeofspores孢子長度/μmLengthofspore孢子寬度/μmWidthofsporeCQMa102卵形Oval3.3~4.62.1~3.3CQMa114長橢圓形Oblong6.6~8.22.3~3.1CQMa117長橢圓形Oblong6.2~8.52.3~3.5CQMa125長橢圓形Oblong6.4~8.31.8~2.5CQMa126長橢圓形Oblong5.5~8.62.2~2.9CQMa128長橢圓形Oblong6.7~8.42.2~3.1CQMa132柱狀Columnar7.5~10.22.2~2.6CQMa135柱狀Columnar9.1~11.82.1~3.2CQMa138桿狀Rod8.8~10.22.2~2.9CQMa140桿狀Rod9.3~11.21.8~2.8

圖1 10株綠僵菌菌株的孢子形態
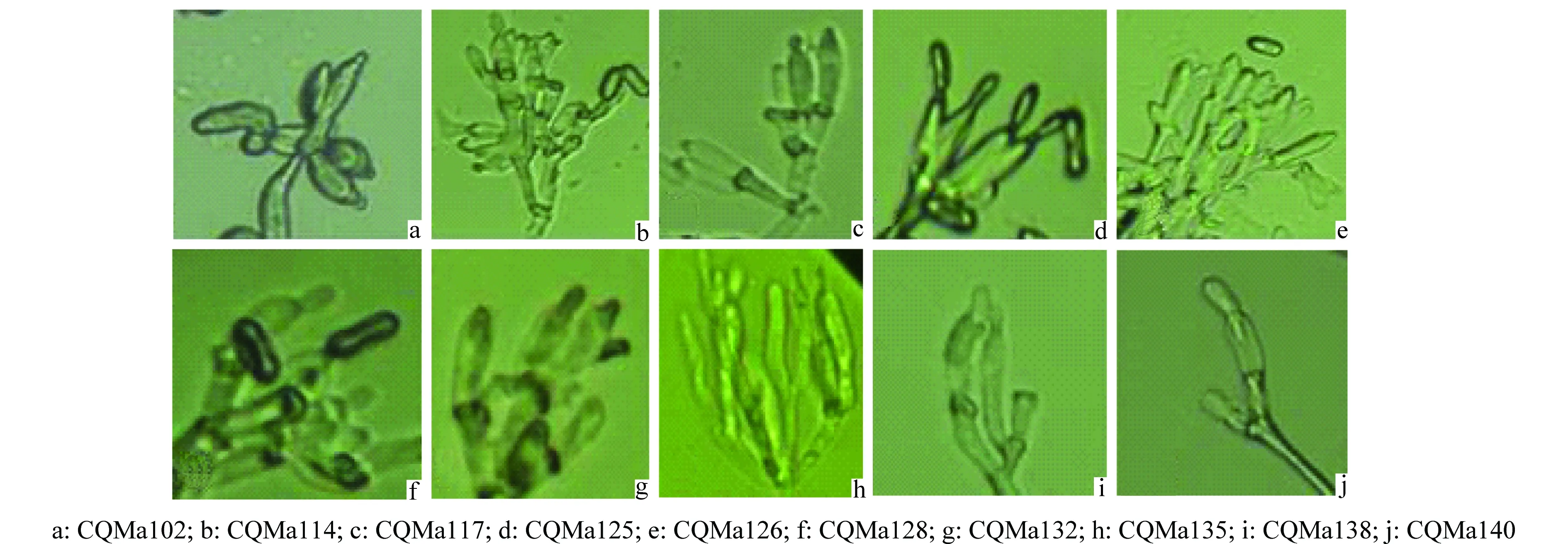
圖2 10株綠僵菌菌株的產孢結構形態
2.2 系統進化分析
2.2.1 單基因系統進化樹及系統進化分析
分別基于ef1-α、rpb1、rpb2、β-tubulin序列構建了單基因MP(maximum parsimony)系統進化樹,單基因系統進化樹結果較為一致的是綠僵菌屬分為4個類群,黃綠綠僵菌單獨為一個類群,獨立于金龜子綠僵菌類群之外。柱狀綠僵菌的分類也較為穩定,在4個單基因系統進化樹中都獨立為一個類群。球形綠僵菌和蝗綠僵菌及目的菌株CQMa102為一個類群,與其他金龜子綠僵菌親緣關系較遠。出現爭議的對其他形態上相近的綠僵菌菌株的親緣關系的劃分,對目的菌株的劃分結果也不一致。值得注意的是菌株CQMa138、CQMa140在4個單基因系統進化樹中都獨立于現有綠僵菌種的類群之外,與現有綠僵菌種的親緣關系較遠。
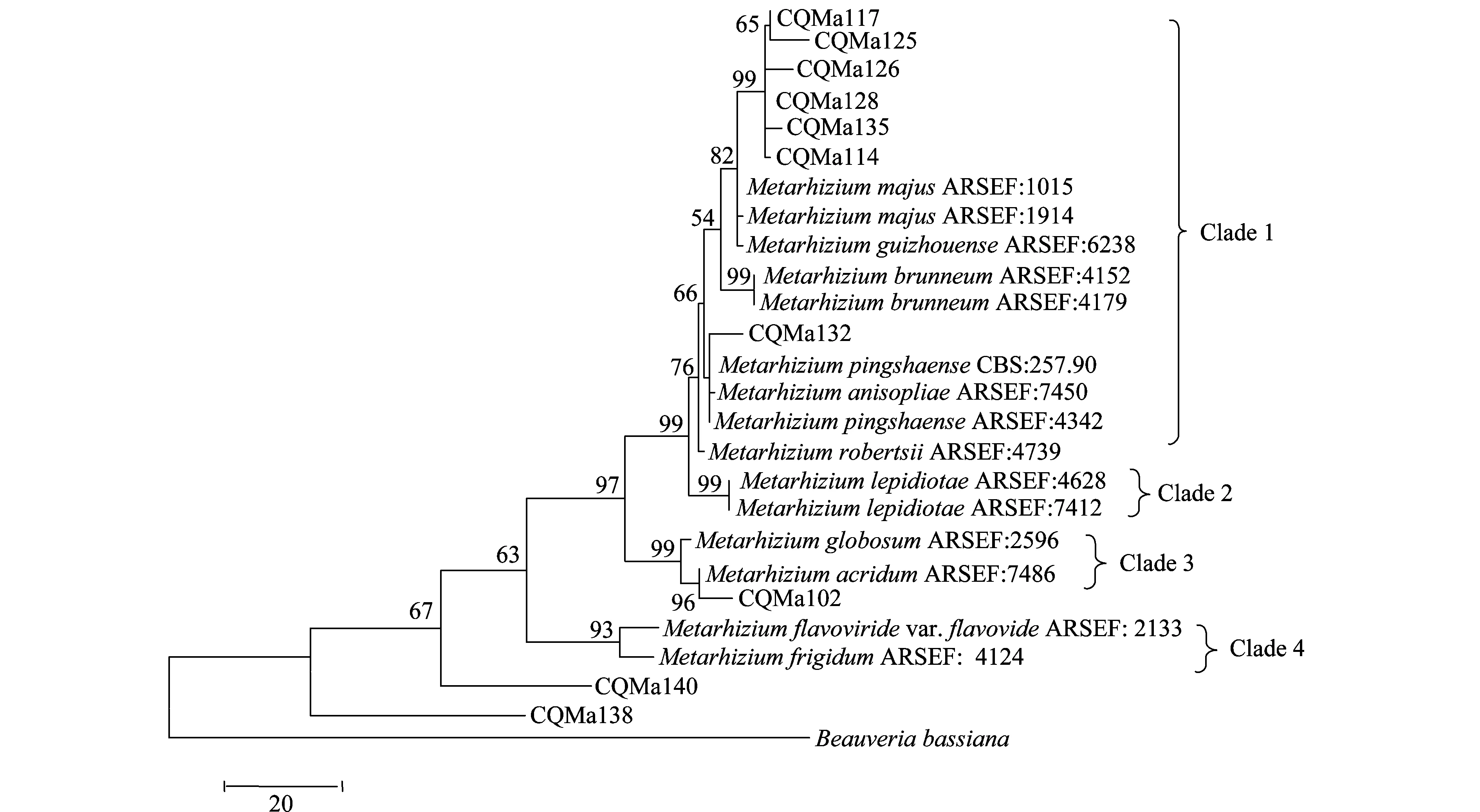
圖3 基于rpb1 序列的單基因MP系統進化樹

圖4 基于rpb2序列的單基因MP系統進化樹
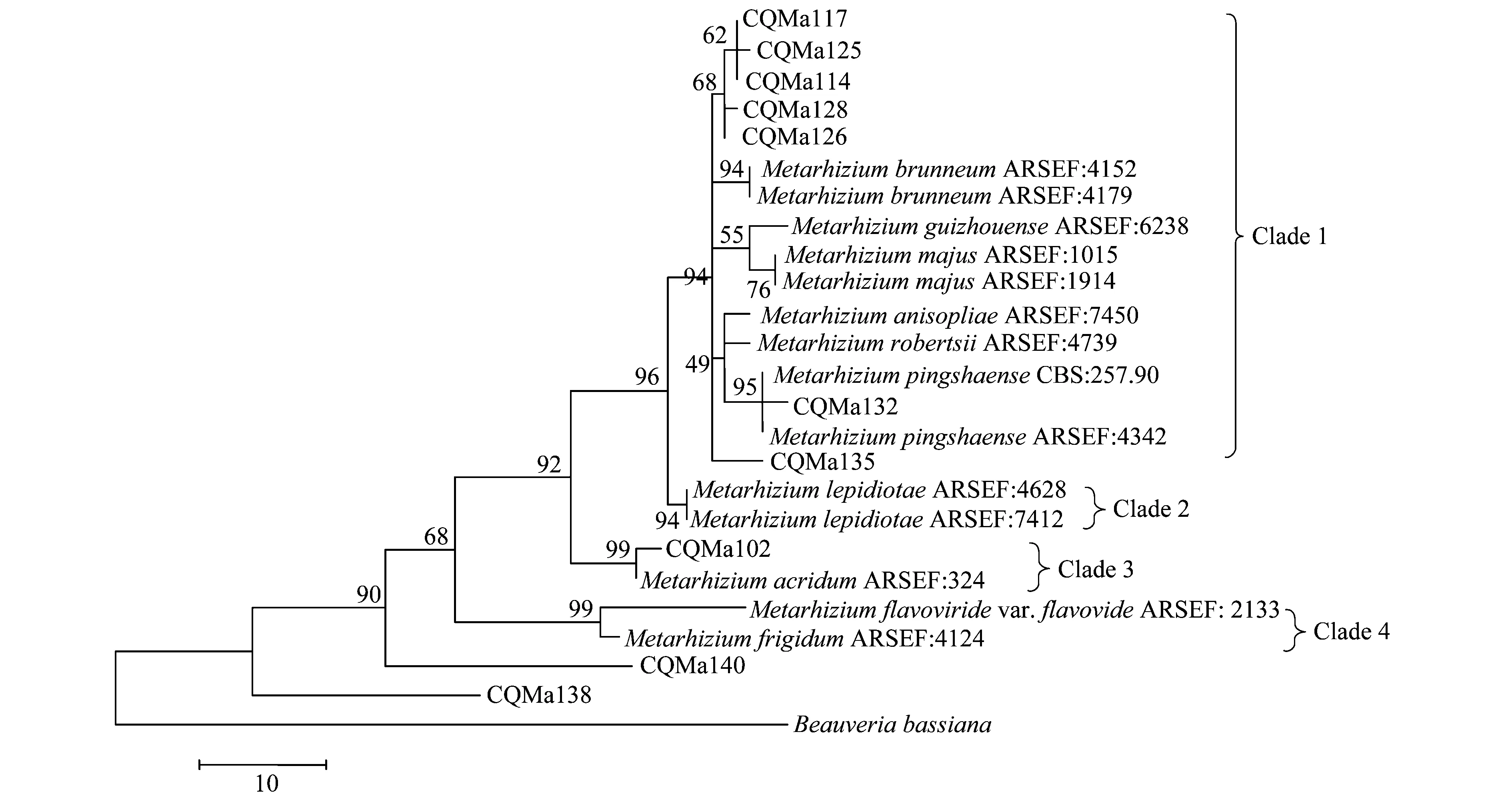
圖5 基于β-tubulin序列的單基因MP系統進化樹
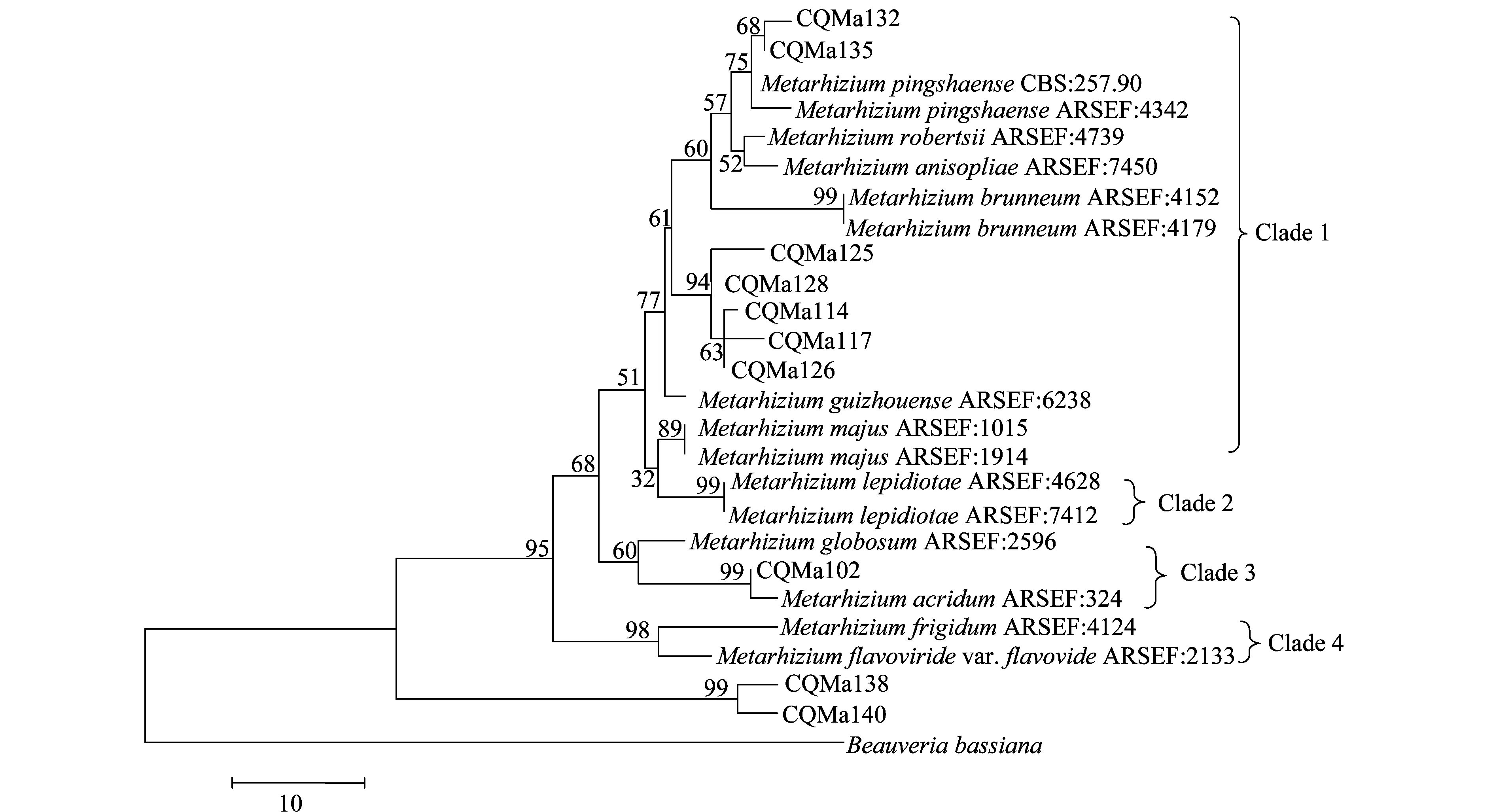
圖6 基于ef1-α 序列的單基因MP系統進化樹
2.2.2 多基因系統進化樹及系統進化分析
以MP(maximum parismony)及N-J(neighbor-joining)兩種方法構建了基于ef-α、rpb、rpb、β-tubulin序列的多基因系統進化樹(圖7、8),相比于單基因系統進化樹,其能正確反映各菌株的親緣關系,不存在同一個種的綠僵菌處在不同分支中的情況。兩種算法構建的多基因系統進化樹中各目的菌株的分類地位一致,各綠僵菌種之間的親緣關系也一致。菌株CQMa102與蝗綠僵菌處在一個分支,支持率分別為96和89,在單基因系統進化樹中與多基因系統進化樹中菌株CQMa102的分類地位一致。 CQMa114、CQMa117、CQMa125、CQMa126、CQMa128聚集在一起,處在大孢綠僵菌這一分支當中,支持率分別為97和92,在單基因系統進化樹中它們的分類地位則不一致,而兩種算法構建的多基因進化樹中他們的分類地位一致。CQMa135、CQMa132的分類地位單基因系統進化樹中不一致,兩種算法的多基因系統進化樹中菌株CQMa132、CQMa135的分類地位一致,與貴州綠僵菌處于一個分支中。從多基因系統進化樹中我們可以看到菌株CQMa138、CQMa140獨立于現有的綠僵菌種的類群之外,與它們的親緣關系較遠,在單基因系統進化樹中兩株綠僵菌也獨立于現有綠僵菌種的類群之外。有可能屬于新的分類單元,還需要深入對菌株的生物學形態、殺蟲譜以及多基因分子鑒定等方面開展研究,方能作出結論。
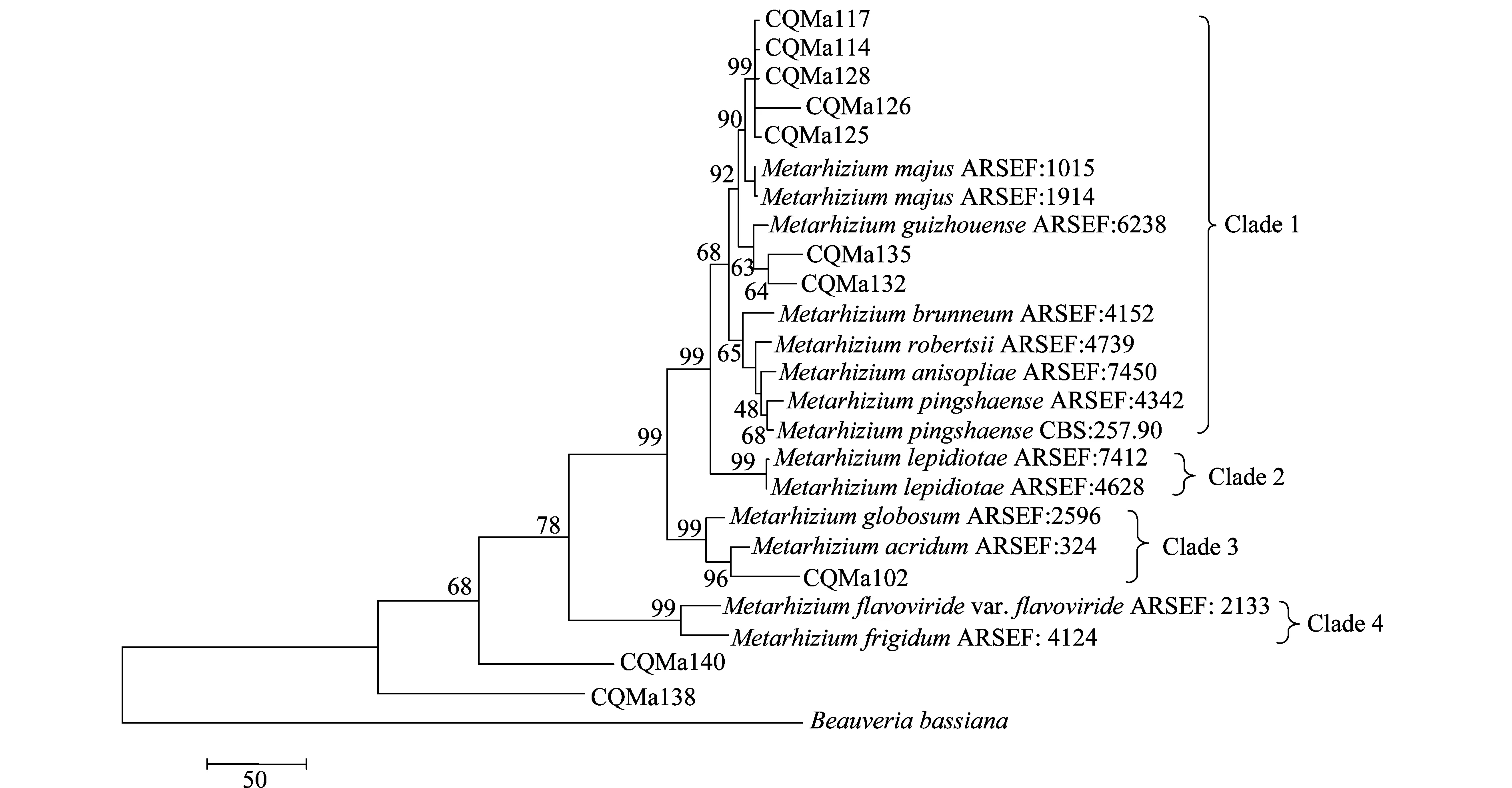
圖7 基于ef1-α、rpb1、rpb2、β-tubulin序列的MP多基因系統進化樹
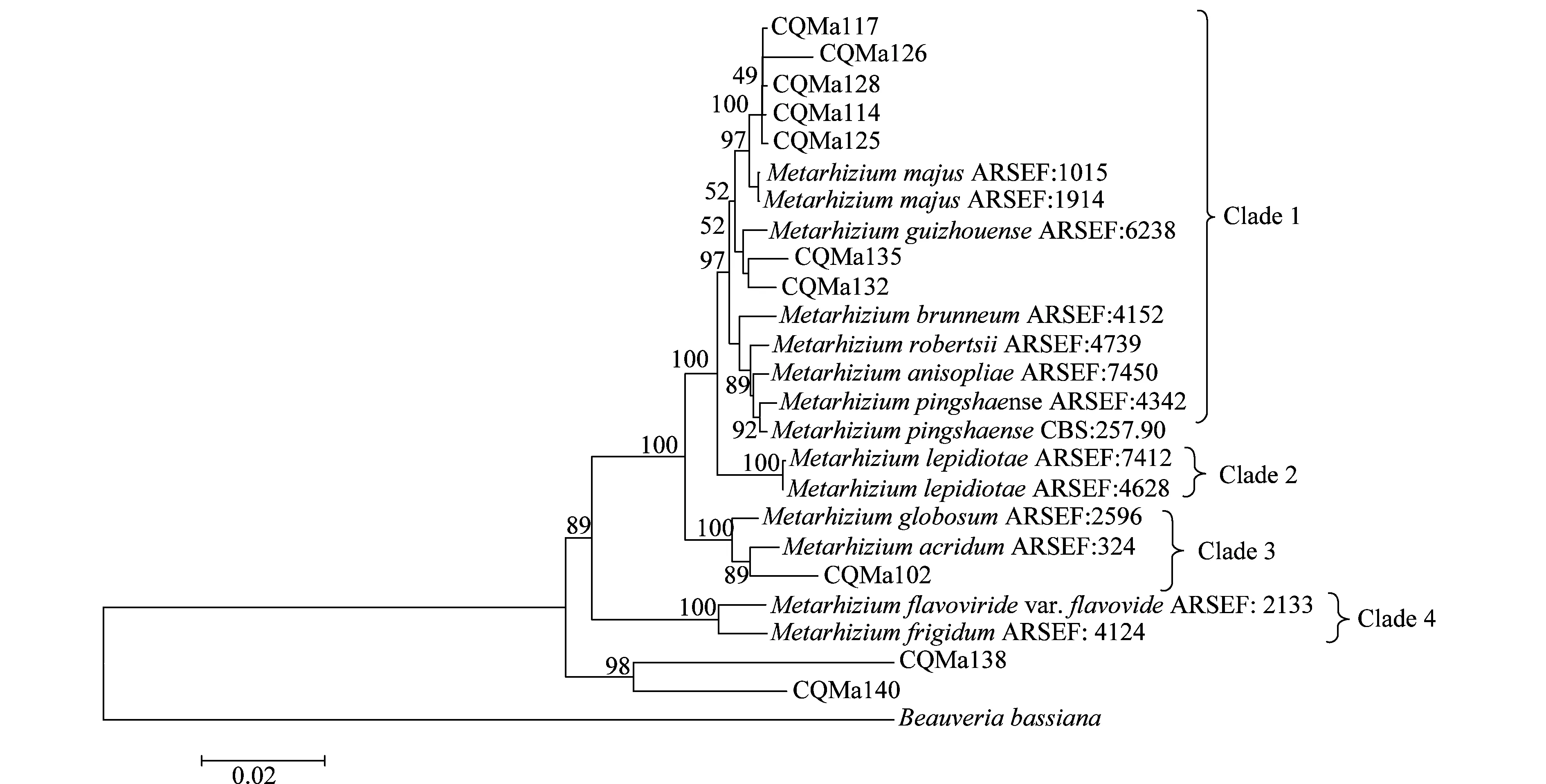
圖8 基于ef1-α、rpb1、rpb2、β-tubulin序列的N-J多基因系統進化樹
3 結論與討論
綠僵菌屬是一個包括多種重要蟲生真菌的真菌類群,具有很高的經濟和社會價值。綠僵菌具有較強的宿主專化性,因此對綠僵菌進行準確的分類鑒定為基于綠僵菌菌種的殺蟲劑研制和環境、生態的保護有著重要意義。綠僵菌屬的分類鑒定方法經歷了從傳統生物形態學的單一性狀到現代分子生物學的基因再到多基因位點分類的演變。由于菌落、瓶梗在不同培養條件下及在同一培養條件下都可能發生變化,且分生孢子大小存在重疊,因此不能完全依靠形態學特征作為分類的標準。隨著分子生物學的發展,含有遺傳信息的功能基因序列用于種類鑒定當中, ITS序列由于所含遺傳信息的局限性,不能區別復合種下形態學相近的物種。在Driver[5]的研究中,形態學明顯差異很大的大孢綠僵菌和金龜子綠僵菌在基于ITS序列的分析中處在一個分支當中,由于不能確定各個綠僵菌種的分類地位從而多賦予變種的分類地位。
在本研究中,基于4個不同基因序列的單基因系統進化樹中可以將分析的菌株分為4個大的類群,4個類群所包含的綠僵菌種基本穩定,但在不同的單基因系統進化樹中類群1中各菌株的親緣關系差異較大,這可能是單個基因所包含的遺傳信息有限,不能清晰地區分親緣關系較近的種,而結合幾個基因則能提供較充分的遺傳信息。其中菌株CQMa114、CQMa117、CQM125、CQMa126、CQMa128在單基因系統進化樹和多基因系統進化樹中都聚集在一起,與大孢綠僵菌親緣關系較近,形態學上分生孢子均為橄欖綠色,長橢圓形,因此鑒定為大孢綠僵菌。來源為河南的菌株CQMa114、CQM125、CQMa126、CQMa128雖寄主不同,但仍然具有很高的遺傳相似性。在Fegan[8]的研究中發現同一地域來源的菌株在RAPD圖譜上具有較大的相似率,說明同一地域來源菌株的遺傳差異較小。在基于rpb1、rpb2、β-tubulin序列的單基因系統進化樹中,貴州綠僵菌(M.guizhouense)均與大孢綠僵菌(M.majus)有很近的親緣關系,但在基于ef1-α的系統進化樹中,貴州綠僵菌卻單獨為一個分支。菌株CQMa132、CQMa135的來源和寄主相同,形態學上菌落均為茸狀,基質均為黃褐色,孢子形態為柱狀,在MP和N-J多基因系統進化樹中以及基于rpb2序列的單基因系統進化樹中均與貴州綠僵菌處在一個分支中,形態特征與貴州綠僵菌相符,因此鑒定為貴州綠僵菌,但在基于rpb1、β-tubulin序列的單基因系統進化樹中兩株綠僵菌卻處在不同的分支中,這說明不同基因的變異頻率有差異,因此單基因的系統進化分析和分類鑒定并不可靠。在Driver的研究中并沒有包括我國的貴州綠僵菌和平沙綠僵菌兩個種。
類群2為兩株柱狀綠僵菌(M.lepidiotae),類群3為蝗綠僵菌(M.acridum)、球孢綠僵菌(M.globosum)和菌株CQMa102,在單基因和多基因系統進化樹中2個類群的分類穩定。菌株CQMa102分離于自然感病的黃脊竹蝗僵蟲,分生孢子為卵形至梨形、黃色,分生孢子梗聚集呈輪生體,形態特征與蝗綠僵菌相似,加上可以侵染多種飛蝗和土蝗,因此鑒定為蝗綠僵菌。類群3在單基因系統進化樹中分類地位穩定且和多基因系統進化樹一致,從形態上看球孢綠僵菌和蝗綠僵菌與金龜子綠僵菌有較大的差異,這說明單基因在區分形態差異大的種時具有一定優勢,而類群1中各綠僵菌菌種的親緣關系在單基因中差異較大,這可能是單基因所包含的遺傳信息有限而不能清晰區分形態相近的綠僵菌種的親緣關系。
類群4為黃綠綠僵菌類群(M.flavoviride),在單基因和多基因系統進化樹中其分類地位穩定,與其他綠僵菌的親緣關系較遠,說明黃綠綠僵菌與金龜子綠僵菌在遺傳上有較大的差異,支持分為獨立的2個種的現狀;這一結果與Bridge[9]的研究一致。值得注意的是,菌株CQMa138、CQMa140在單基因和多基因系統進化樹中都獨立于現有綠僵菌種的類群之外,與其他綠僵菌種的親緣關系較遠,從形態上看CQMa138、CQMa140孢子均為桿狀,顏色分別為綠色和灰綠色,瓶梗數量較少,與其他綠僵菌種的形態有比較明顯的差異,結合系統進化分析結果和形態特征,初步推測CQMa138、CQMa140可能為綠僵菌屬的新種或新的分類單元,但在MP和N-J多基因系統進化中,兩個菌株的親緣關系又有比較明顯的差異,兩者的親緣關系還不明確,因此新種地位的確定還需更多的證據支撐。
[1]全宇, 劉永翔, 劉作易. 綠僵菌屬分類的研究進展[J]. 貴州農業科學, 2011, 39(10): 113-117.
[2]de Faria M R, Wraight S P. Mycoinsecticides and mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types[J]. Biological Control, 2007, 43:237-256.
[3]黃勃, 樊美珍, 李增智. 綠僵菌屬系統分類的研究進展(綜述)[J]. 安徽農業大學學報, 2002, 29(2): 169-172.
[4]Peng G X, Wang Z K, Yin Y P, et al. Field trials ofMetarhiziumanisopliaevar.acridum(Ascomycota: Hypocreales) against oriental migratory locusts,Locustamigratoriamanilensis(Meyen) in Northern China[J].Crop Protection,2008,27(9):1244-1250.
[5]Driver F, Milner R J, Trueman J W H. A taxonomic revision ofMetarhiziumbased on a phylogenetic analysis of rDNA sequence data[J]. Mycological Research, 2000, 104(2): 134-150.
[6]Bischoff J F, Rehner S A, Humber R A.Metarhiziumfrigidumsp. nov.: a cryptic species ofM.anisopliaeand a member of theM.flavoviridecomplex[J]. Mycologia, 2006, 98: 737-745.
[7]Bischoff J F, Rehner S A, Humber R A.A multilocus phylogeny of theMetarhiziumanisopliaelineage[J]. Mycologia, 2009, 101(4): 512-530.
[8]Fegan M, Manners J M, Maclean D J. Random amplified polymorphic DNA markers reveal a high degree of genetic diversity in the entomopathogenic fungusMetarhiziumanisopliaevar.anisopliae[J]. Journal of General Microbiology, 1993, 139: 2075-2081.
[9]Bridge P D, Williams M A J, Prior C. Morphological, biochemical and molecular characteristics ofMetarhiziumanisopliaeandM.flavoviride[J]. Journal of General Microbiology, 1993, 139: 1163-1165.
Multilocusphylogeneticanalysisofthetaxonomicstatusof10strainsofMetarhizium
Wang Meng, Yin Youping, Wang Zhongkang, Liu Juanjuan, Liao Yufeng
(SchoolofLifeSciences,ChongqingUniversity,ResearchCenterforGeneticEngineering,Chongqing400030,China)
In order to confirm the taxonomic status of 10 high-efficiency pesticide isolates ofMetarhizium, we employed a multilocus phylogenetic analysis approach by usingef1-α,rpb1,rpb2 andβ-tubulingene sequences and morphological characteristics to identify the taxonomic status of isolatedMetarhizium, the results proved that the multilocus phylogenetic analysis method was more accurate compared to single gene phylogenetic analysis. We identified CQMa114, CQM117, CQMa125, CQMa126 and CQMa128 asM.majus, CQMa102 asM.acridum, and CQMa132 and CQMa135 asM.guizhouense. The strains CQMa138 and CQMa140 might be new species ofMetarhizium.
Metarhizium; classification and determination; multilocus phylogenetic analysis; morphology
2013-09-16
:2014-05-24
公益性行業(農業)科研專項(201103002)
S 476.12
:ADOI:10.3969/j.issn.0529-1542.2014.05.003
* 通信作者 E-mail: w-zk@163.com
猜你喜歡
工業設計(2022年8期)2022-09-09 07:43:20
數學小靈通(1-2年級)(2021年4期)2021-06-09 06:25:56
大眾健康(2021年6期)2021-06-08 19:30:06
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
裝備制造技術(2019年12期)2019-12-25 03:06:46
中學生數理化·七年級數學人教版(2019年4期)2019-05-20 10:06:32
中國洗滌用品工業(2019年4期)2019-05-11 09:27:34
中學生數理化·七年級數學人教版(2018年6期)2018-06-26 08:36:06
初中生世界·七年級(2017年9期)2017-10-13 22:27:46
