甘肅省馬鈴薯束梗褐腐病病原鑒定及其生物學特性研究
2014-08-10 12:29:50陳泰祥陳秀蓉楊成德朱海波王涵琦
植物保護 2014年5期
關鍵詞:生長
陳泰祥, 陳秀蓉, 楊成德, 朱海波, 王涵琦, 卞 靜
(甘肅農業(yè)大學草業(yè)學院,草業(yè)生態(tài)系統(tǒng)教育部重點實驗室,甘肅省草業(yè)工程實驗室,中-美草地畜牧業(yè)可持續(xù)發(fā)展研究中心,蘭州 730070)
甘肅省馬鈴薯束梗褐腐病病原鑒定及其生物學特性研究
陳泰祥, 陳秀蓉*, 楊成德, 朱海波, 王涵琦, 卞 靜
(甘肅農業(yè)大學草業(yè)學院,草業(yè)生態(tài)系統(tǒng)教育部重點實驗室,甘肅省草業(yè)工程實驗室,中-美草地畜牧業(yè)可持續(xù)發(fā)展研究中心,蘭州 730070)
對甘肅省馬鈴薯(SolanumtuberosumL.)束梗褐腐病病原進行了分離鑒定和生物學特性研究。經形態(tài)特征觀察與rDNA-ITS序列分析,將病原菌鑒定為細基束梗霉[Doratomycesstemonitis(Pers.exFr.) F.J. Morton & G. Smith]。生物學特性研究結果表明,病菌菌絲生長最適溫度為25~30 ℃,最適pH為6, 病菌能利用多種碳源,但以蔗糖最好,氮源以甘氨酸最適,光照對菌絲生長沒有影響。分生孢子在5~40 ℃范圍內均能萌發(fā),最適25 ℃,最適pH為7,分生孢子萌發(fā)需液態(tài)水,濕度低于99%幾乎不萌發(fā),馬鈴薯汁液和葡萄糖液對孢子萌發(fā)有較好的促進作用。本研究為馬鈴薯束梗褐腐病的防治提供了理論依據。
馬鈴薯; 細基束梗霉; 病原分離鑒定; 生物學特性
馬鈴薯(SolanumtuberosumL.)是甘肅省最具優(yōu)勢的地方特色糧食作物,全省87個縣(市、區(qū))中有60多個縣種植馬鈴薯,特別是定西有著“中國馬鈴薯之鄉(xiāng)”的美稱[1]。近年來,隨著農業(yè)生產水平的提高,馬鈴薯種植規(guī)模不斷擴大,馬鈴薯儲藏期病害的發(fā)生和危害程度也愈加廣泛和嚴重。甘肅省已報道的馬鈴薯儲藏期病害有13種,主要有晚疫病、干腐病、炭疽病、絲核病、壞疽病、軟腐病、瘡痂病和病毒病等[2-6],病害發(fā)生造成大量爛薯,嚴重影響馬鈴薯產量和商品價值,所以有效地控制馬鈴薯儲藏期病害成為生產上亟待解決的問題。自2009年以來,甘肅省儲藏期馬鈴薯發(fā)生一種不常見病害,發(fā)病薯塊組織變褐腐爛,并形成扁平的不規(guī)則腔室,嚴重影響薯塊的品質。據調查,部分地區(qū)發(fā)病率高達20%~30%,而且病情逐年加重,發(fā)生面積不斷擴大,給生產上馬鈴薯的儲藏和供應帶來了較嚴重影響,已逐漸成為甘肅省馬鈴薯儲藏期的主要病害之一。關于此病害的研究,李金花[6]從采自張掖的病薯中分離到了該病菌,并對其病原進行了描述,認為是具柄細基束梗孢 (Cephalotrichumstemonitis),有文獻記載該病原菌能侵染儲藏期的馬鈴薯引起褐腐病,還能夠從水稻、玉米種子中分離到,并且曾從中華繡線梅和土壤中也分離到了該菌[7]。而該病害的病原生物學特性未見研究報道,因此,筆者對馬鈴薯褐腐病的病原以及生物學特性進行了較系統(tǒng)的研究,旨在為有效防治此病害提供依據。
1 材料和方法
1.1 材料
1.1.1 病薯來源
分別于2012年10月和2013年3月采自定西市安定區(qū)、張掖市肅南縣、山丹縣、民樂縣,武威市天祝縣等地的馬鈴薯儲藏庫中。采后置于封口塑料袋中,保持新鮮狀態(tài),帶回實驗室,觀察癥狀,描述并拍照。
1.1.2 供試儀器
NIKON光學顯微鏡,NIKON圖像分析系統(tǒng),PHS-3 c精密數顯酸度計,Bioneer水平凝膠電泳裝置,Bioneer PCR熱循環(huán)儀,Bioneer凝膠成像系統(tǒng),Biometra高速冷凍離心機等。
1.1.3 供試試劑
Rnase, Proteinase K,PCR Master Mix, Marker購于上海生工生物技術公司,通用引物ITS1(5′-TCC GTA GGT GAA CCT GCG G-3′)ITS4(5′-TCC TCC GCT TAT TGA TAT GC-3′)由上海生工生物技術公司合成,其他化學試劑均采用國產分析純。
1.1.4 供試培養(yǎng)基
PDA、Czapek培養(yǎng)基[8]。
1.2 方法
1.2.1 病原分離
采用組織分離法在PDA上進行病原菌的分離。
1.2.2 病原菌致病性測定
根據柯赫氏法則[8],用病菌分生孢子懸浮液分別以有傷(針刺傷)和無傷方式接種于健康的馬鈴薯薯塊上,并保濕。每處理3個重復,以接清水為對照。發(fā)病后從病健交界處再次分離病原菌。
1.2.3 病原菌鑒定
1.2.3.1病原菌形態(tài)學鑒定
將病菌接種于PDA平板,置25 ℃培養(yǎng)箱培養(yǎng),觀察菌落顏色、形狀、氣生菌絲疏密程度。在顯微鏡下觀察分生孢子、產孢細胞、孢梗束形態(tài)并顯微拍照,測量50個分生孢子的大小。根據病原形態(tài),參照相關文獻[7-9]進行病原鑒定,確定病原屬種。
1.2.3.2病原菌rDNA-ITS序列分析及系統(tǒng)發(fā)育地位
(1)DNA的提取
將病原菌接種于PDA培養(yǎng)液中,于25 ℃、150 r/min條件下振蕩培養(yǎng)4~5 d后收集菌絲,采用SDS和氯化芐結合法[10]提取基因組總DNA。
(2)rDNA-ITS擴增與序列測定
采用通用引物ITS1和ITS4擴增病菌rDNA-ITS片段。PCR反應體系為50 μL:即DreamTaqGreen PCR MasterMix(2×)25 μL,ITS1(10 mmol/L)1.0 μL,ITS4(10 mmol/L)1.0 μL,DNA模板1.0 μL,ddH2O 22 μL,以不加DNA模板而加等量ddH2O為陰性對照。根據預試驗結果優(yōu)化組合而得出以下PCR擴增反應程序:94 ℃預變性3 min,以下共30個循環(huán)(94 ℃變性30 s,55 ℃退火30 s,72 ℃延伸1 min),最后72 ℃延伸8 min。PCR產物經0.8%瓊脂糖凝膠電泳檢測后,送上海生工生物技術公司進行測序。
(3)序列比對和系統(tǒng)發(fā)育樹的構建
將病原菌rDNA-ITS序列在NCBI上用BALST進行同源性比較后,下載同源序列,用Clustal(version 1.81)進行多重序列匹配排列(multiple alignments)分析,形成一個多序列匹配排列陣,然后用Mega(version 4.0) 通過鄰接法(neighbor-joining)構建系統(tǒng)發(fā)育樹[11]。
1.2.4 病原菌生物學特性測定
1.2.4.1病原菌菌絲生長測定
(1)溫度對病原菌菌絲生長的影響
用直徑為0.5 cm的打孔器在菌落邊緣打取菌餅,接種于PDA平板中央,分別置于5、10、15、20、25、30、35 ℃和40 ℃的恒溫培養(yǎng)箱培養(yǎng),每處理3次重復,15 d后十字交叉法測量菌落直徑。
(2)pH對病原菌菌絲生長的影響
用HCl和NaOH將PDA的pH分別調成4、5、6、6.5、7、8、9、10、11,每處理3次重復,25 ℃恒溫培養(yǎng),接種、菌絲生長測量方法同1.2.4.1(1),(下同)。
(3)碳源對病原菌菌絲生長的影響
以Czapek培養(yǎng)基為基礎培養(yǎng)基,分別以乳糖、D-甘露糖、D-半乳糖、D-果糖、D-木糖、葡萄糖、麥芽糖、L-山梨糖、可溶性淀粉等量替換(添加量均為3%)蔗糖配制成含不同碳源的培養(yǎng)基。以不加碳源為對照,每處理3次重復 ,25 ℃恒溫培養(yǎng)15 d。
(4)氮源對病原菌菌絲生長的影響
以Czapek培養(yǎng)基為基礎培養(yǎng)基,分別以硝酸鉀、酵母膏、蛋白胨、硫酸銨、天門冬酰胺、甘氨酸、L-組氨酸、L-色氨酸、L-亮氨酸、L-精氨酸、DL-蘇氨酸、L-酪氨酸、牛肉膏等量替換(添加量均為0.2%)硝酸鈉配制成含不同氮源的培養(yǎng)基,以不加氮源為對照,每處理3次重復,25 ℃恒溫培養(yǎng)15 d。
(5)光照對病原菌菌絲生長的影響
將直徑為0.5 cm菌餅接種于PDA平板中央,分別置于24h/d光照、光暗自然交替、24 h/d黑暗條件下25 ℃恒溫培養(yǎng)15 d,每處理3次重復。
1.2.4.2病原菌分生孢子萌發(fā)的測定
(1)溫度對病原菌分生孢子萌發(fā)的影響
采用懸滴法,將適當濃度的孢子懸浮液滴在蓋玻片上,蓋至孢子萌發(fā)器上。分別置于5、10、15、20、25、30、35和40 ℃的恒溫條件下培養(yǎng),于18 h后鏡檢孢子萌發(fā)率。每處理3次重復,每次鏡檢不少于200個孢子(下同)。
(2)濕度對病原菌分生孢子萌發(fā)的影響
在干燥器內用硫酸法控制濕度[8],將孢子粉撒于載玻片上,置于相對濕度為99%、95%、 90%和85%條件下室溫(20~22 ℃)培養(yǎng)。以蒸餾水中孢子萌發(fā)率為對照,于18 h后挑取各處理孢子,無菌水制片,檢查萌發(fā)率。
(3)pH對病原菌分生孢子萌發(fā)的影響
用磷酸二氫鉀和磷酸氫二鈉緩沖液將孢子懸浮液pH配制為3、4、5、6、7、8、9、10、11。25 ℃恒溫培養(yǎng),于18 h后鏡檢孢子萌發(fā)率。
(4)營養(yǎng)液對病原菌分生孢子萌發(fā)的影響
以濃度為1∶5,1∶10,1∶20的馬鈴薯汁液、葡萄糖液和土壤浸漬液配制孢子懸浮液,置于25 ℃條件下恒溫培養(yǎng),于18 h后鏡檢孢子萌發(fā)率[12]。
1.2.5 數據處理
以上各試驗所得數據,采用IBM SPSS Statistics 19.0軟件進行差異顯著性分析(DMRT法)。
(四)在國內產能過剩的情況下,中國光伏“走出去”的步伐將進一步加快,以產融結合、廠商租賃等模式將產能向西方國家以及“一帶一路”沿線國家轉移,將成為中國未來光伏產業(yè)持續(xù)健康發(fā)展的現實選擇。
2 結果與分析
2.1 病害癥狀
根據調查,此病害主要發(fā)生在儲藏期的薯塊上,據觀察,從各地采集的病薯癥狀基本一致,多在芽眼處產生中小型灰褐色凹陷斑,斑上有開裂小孔,病菌向薯塊內部擴展,組織變褐腐爛,并形成扁平的不規(guī)則腔室。向外靠健康組織的部分變?yōu)楹稚颇舅ɑ壳皇乙贿厼檩^厚的黑色菌絲層,其上產生灰黑色菌絲,菌絲上產生直立的暗褐色粗壯束梗,梗的上部膨大成長橢圓體,基部較細,成叢產生,狀如毛刷,肉眼可見(圖1)。
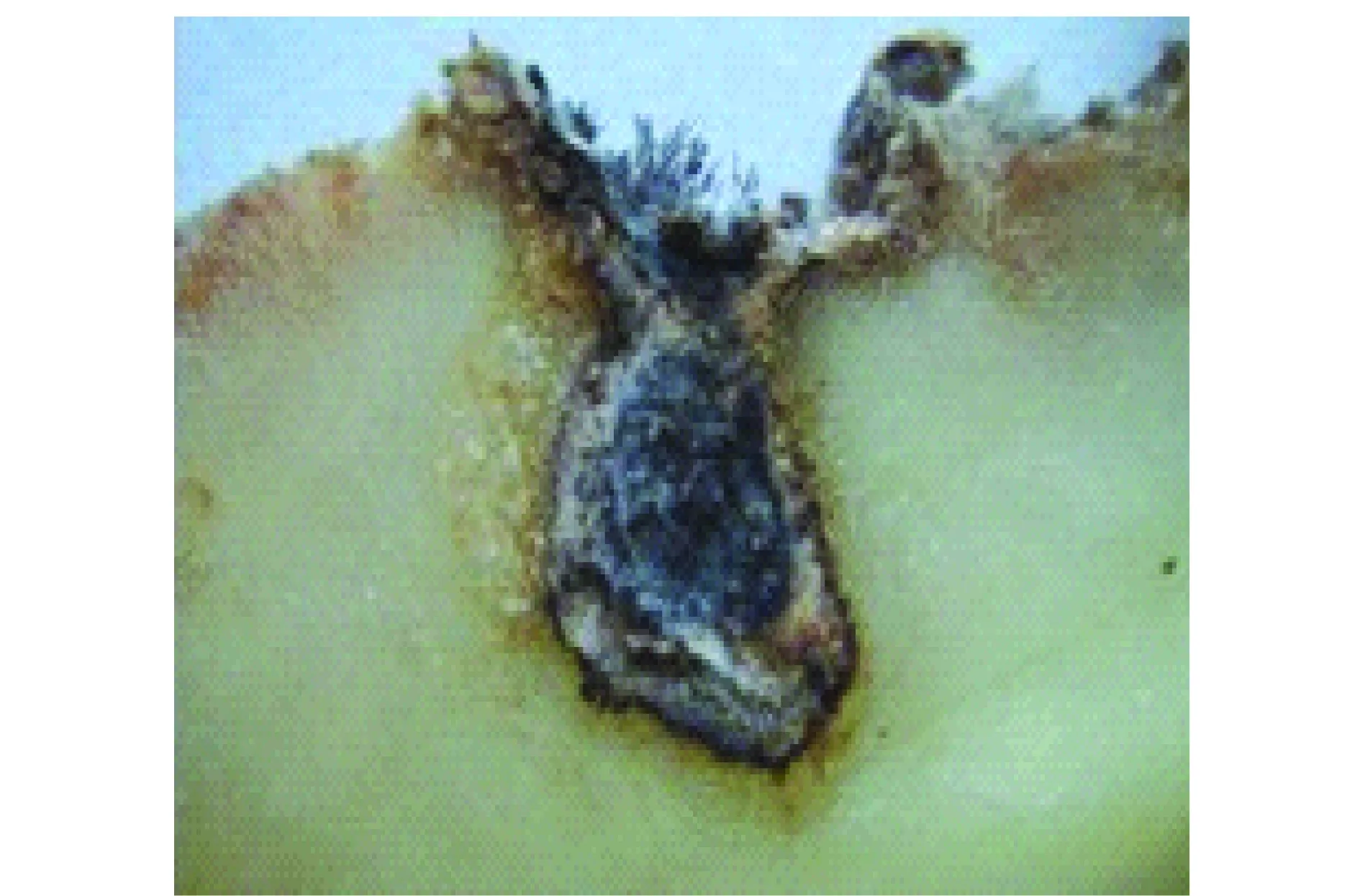
圖1 馬鈴薯褐腐病癥狀
2.2 致病性測定結果
接種15 d后接菌部位癥狀較明顯,與自然發(fā)病的癥狀相似,病部邊緣凹陷,菌絲體從邊緣生長而出,菌絲體褐色,絨狀,對照表面邊緣凹陷但沒有任何病狀;薯塊接菌部位內部褐色發(fā)黑,向內擴展(圖2a),對照無此癥狀(圖2b),再次分離,從接種的薯塊上獲得了與原接種菌相同的病原菌,證明該菌為馬鈴薯褐腐病的病原菌。
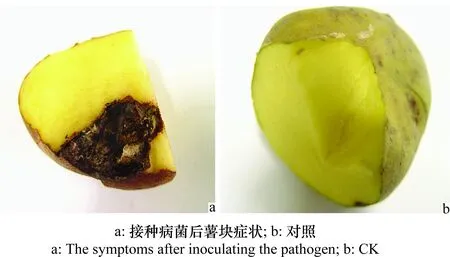
圖2 病原菌對馬鈴薯致病性測定
2.3 病原菌鑒定
2.3.1 病原菌培養(yǎng)特性及形態(tài)特征
據觀察,該菌在PDA上培養(yǎng)15 d后菌落背面灰褐色,呈細絨狀,菌落呈放射狀生長(圖3 b),菌落正面灰褐色,外圈有白色的絨狀生長圈,菌落表面絨狀,深淺不勻(圖3a)。
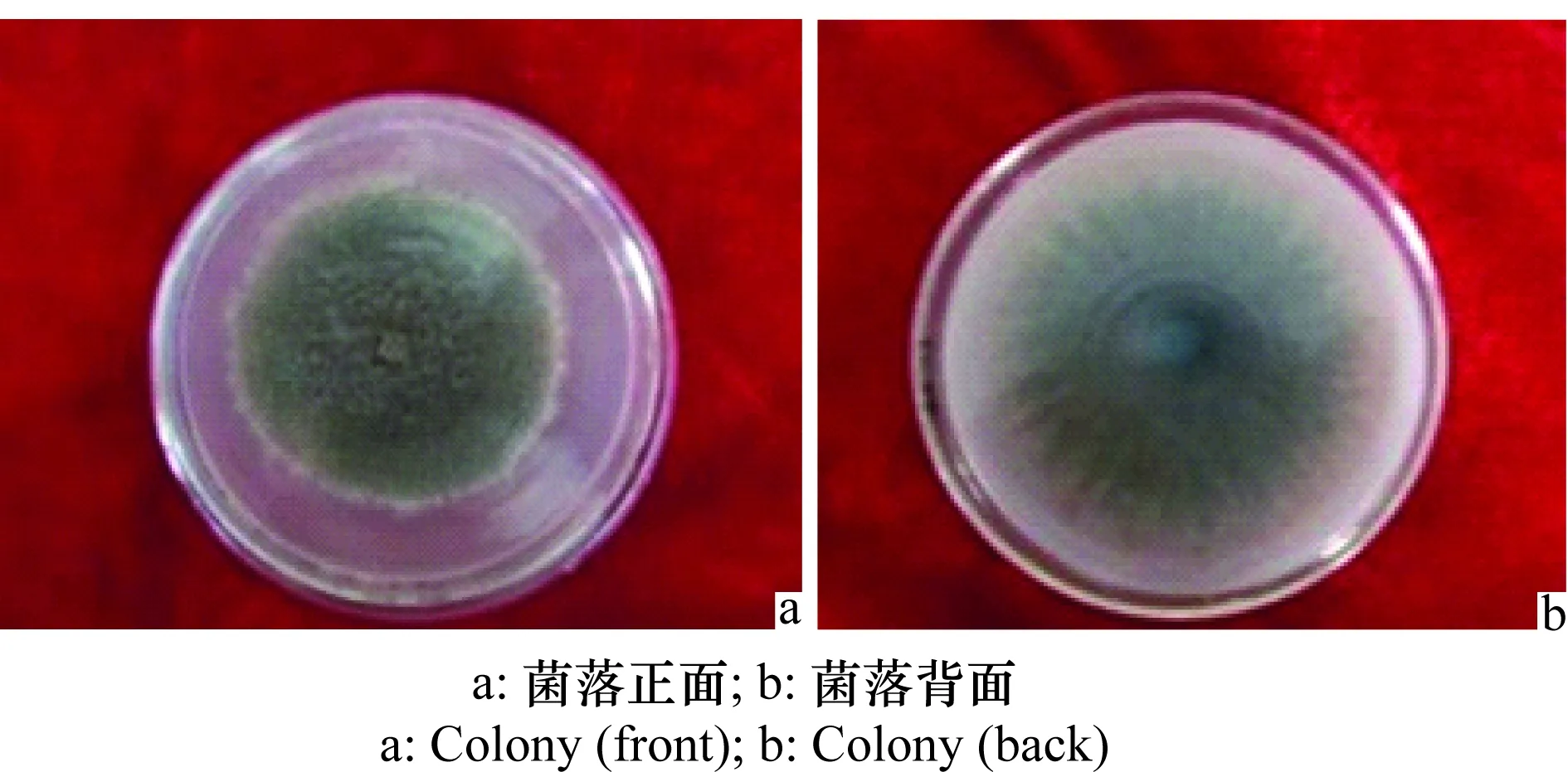
圖3 馬鈴薯褐腐病病菌菌落形態(tài)
病原菌菌絲體初無色,后變淡褐至褐色,有隔。分生孢子梗束狀簇生(見圖4 a,b),最多有7根;束梗基部有假根(棒狀體短于束梗)。梗的中上部有3~5個分枝,分枝頂部膨大呈棒狀或長橢圓體(圖4 d)。

圖4 馬鈴薯褐腐束梗霉(Doratomyces stemonitis)顯微形態(tài)
2.3.2 病菌rDNA-ITS序列分析
測序結果表明,該病菌(S1)序列長度為586 bp(圖5),經BLAST分析,與病菌測定序列同源性達97%~99%的菌株均為Doratomycesspp.下載相似性最高的序列,進一步用Clustal(version 1.81)和Mega(version 4.0) 通過鄰接法(neighbor-joining)構建系統(tǒng)發(fā)育樹。該病菌在系統(tǒng)發(fā)育樹中與Doratomycesstemonitis(登錄號FJ914696.1)聚在一起(圖6),說明其親緣關系最近,從分子水平上進一步鑒定該病原菌為Doratomycesstemonitis。
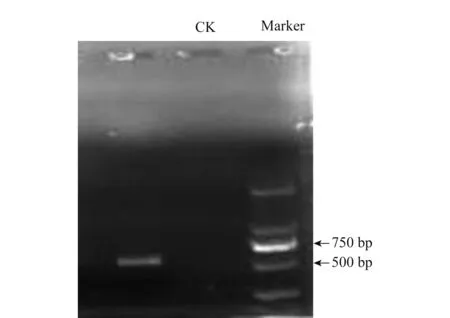
圖5 馬鈴薯褐腐病病菌rDNA-ITS PCR電泳圖

圖6 馬鈴薯褐腐病病菌系統(tǒng)發(fā)育樹
2.4 病原菌菌絲生長測定結果
2.4.1 溫度對病原菌菌絲生長的影響
測定結果(圖7)表明,在5~35 ℃范圍內均能生長,以25~30 ℃條件下生長最快,30 ℃菌落直徑達67 mm,低于5 ℃和超過35 ℃停止生長。表明該菌適宜生長溫度為25~30 ℃。

圖7 溫度對馬鈴薯褐腐病菌菌絲生長的影響
2.4.2 pH對病原菌菌絲生長的影響
不同pH對菌絲生長影響(圖8)有明顯差異,菌絲在4~11范圍內均能生長,最適pH為6。pH 4~5和pH 9~11菌絲雖能生長成正常菌落,但生長緩慢。此菌對酸度的適應范圍廣并偏好略帶酸性的培養(yǎng)條件,而馬鈴薯汁液呈中性偏酸性,可見該病菌侵染馬鈴薯后在適宜的溫濕度條件下可迅速蔓延。

圖8 pH對馬鈴薯褐腐病菌菌絲生長的影響
2.4.3 碳源對病原菌菌絲生長的影響
從測定結果(圖9)可見,該病菌對單糖、雙糖多糖等碳源都能利用,各供試碳源除L-山梨糖外菌落直徑均大于對照菌落直徑,且方差分析差異顯著, L-山梨糖抑制該病原菌的生長。可見該菌對碳源的需求不嚴格。

圖9 不同碳源對馬鈴薯褐腐病菌菌絲生長的影響
2.4.4 氮源對病原菌菌絲生長的影響
從測定結果(圖10)可見,病菌對氮源的利用表現明顯差異。其中甘氨酸、硫酸銨、天門冬酰胺最適合菌絲生長,在以硝酸鉀、酵母膏和DL-蘇氨酸為氮源的培養(yǎng)基上生長良好,而L-組氨酸、L-色氨酸和L-精氨酸等幾種氮源抑制菌絲的生長。
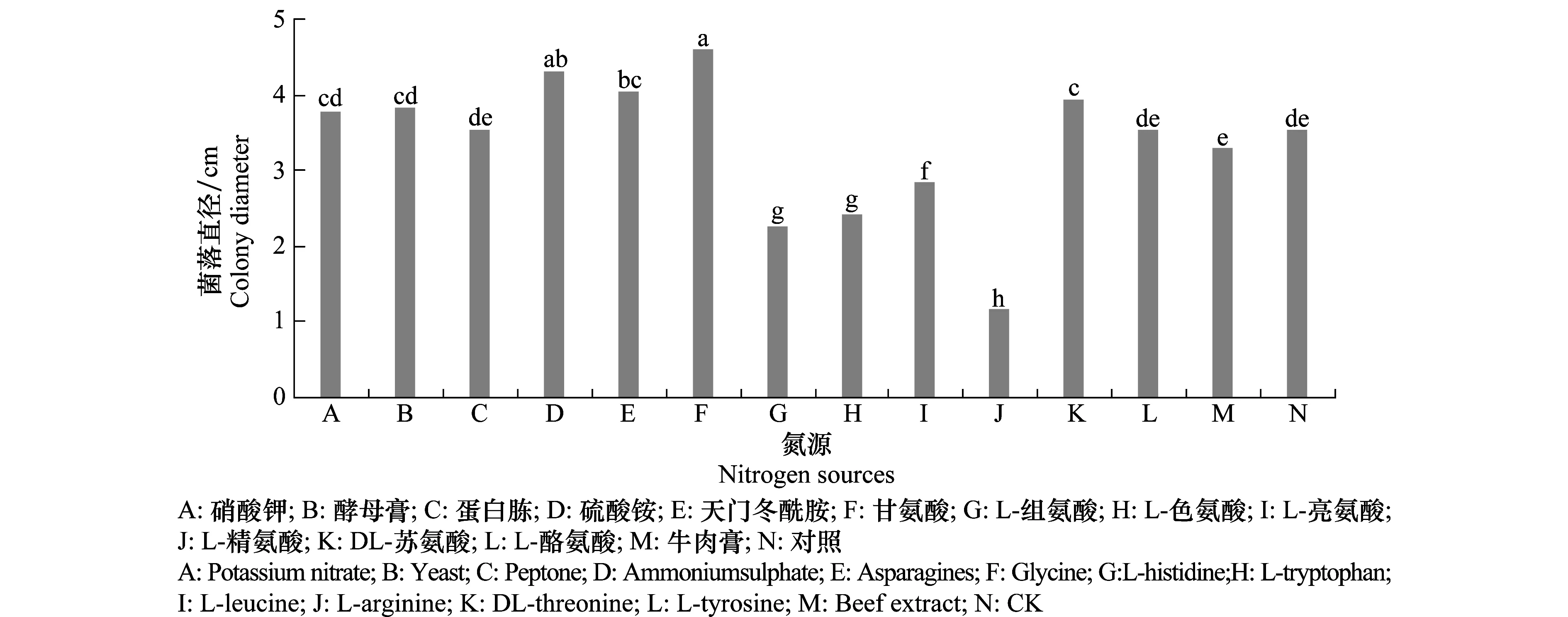
圖10 不同氮源對馬鈴薯褐腐病菌菌絲生長的影響
2.4.5 光照對病原菌菌絲生長的影響
測定結果(表1)表明,不同光照條件下病菌菌絲生長速度基本一致,菌落直徑均在63~70 mm之間,方差分析差異不顯著。說明該病原對光照不敏感,在光照和黑暗條件下均可正常生長。
表1光照對馬鈴薯褐腐病菌菌絲生長的影響
Table1EffectsofilluminationonmycelialgrowthofDoratomycesstemonitis

處理Treatment菌落直徑/mmAveragegrowthofcolony暗、光自然交替Dark,lightalternation(66.0±2.00)a24h/d黑暗24h/ddark(65.3±4.04)a24h/d光照24h/dlight(61.7±5.69)a
2.5 病原菌分生孢子萌發(fā)測定結果
2.5.1 溫度對病原菌分生孢子萌發(fā)的影響
試驗結果(圖11)表明,病菌分生孢子在5~40 ℃范圍內均能萌發(fā),20~30 ℃內萌發(fā)率保持在較高的水平。最適萌發(fā)溫度為25 ℃,18 h萌發(fā)率達90%以上,在5 ℃和40 ℃條件下有20%左右分生孢子可以萌發(fā),表明此菌對溫度的要求不嚴格,這種特性有利于其種群的生存和繁衍。
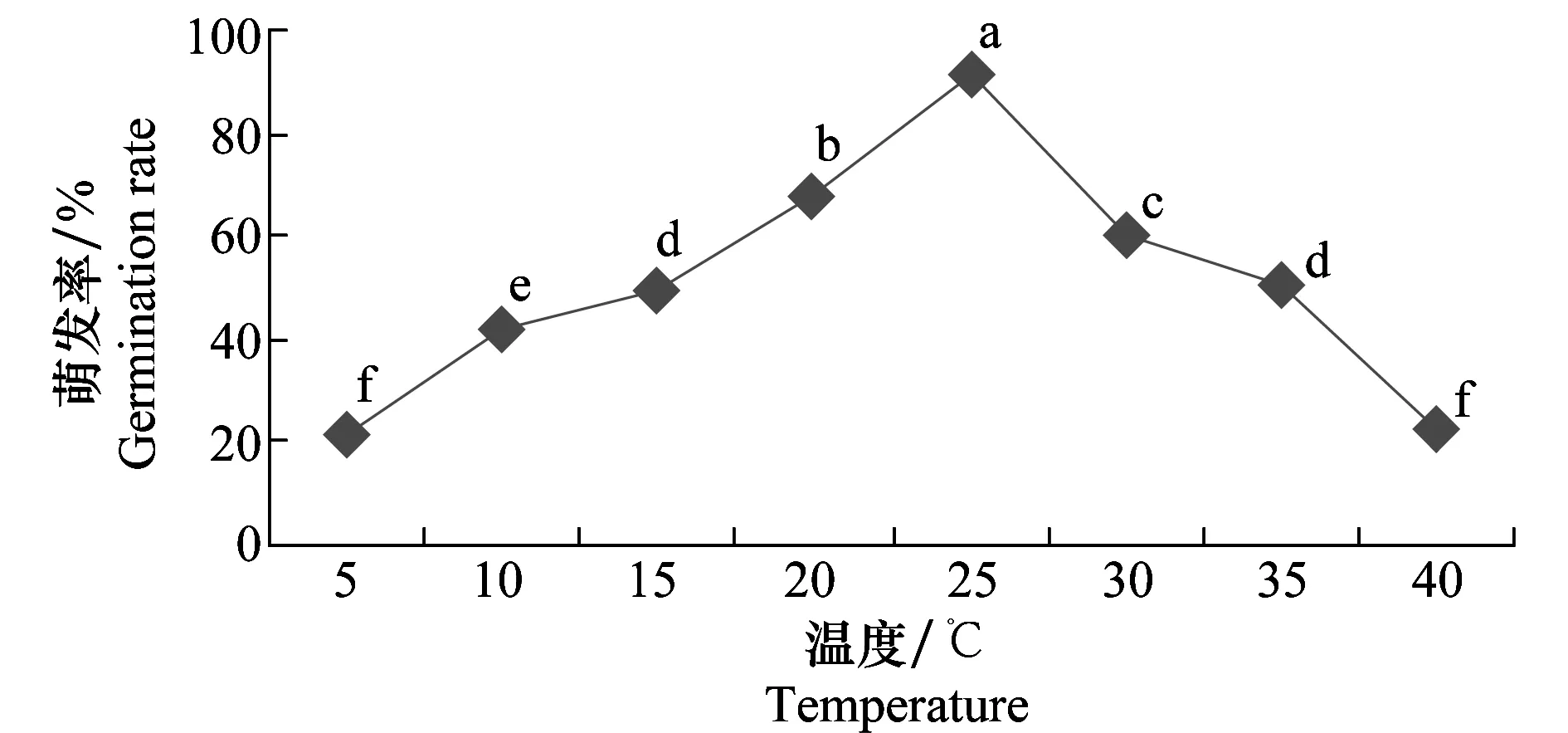
圖11 溫度對馬鈴薯褐腐病菌分生孢子萌發(fā)的影響
2.5.2 濕度對病原菌分生孢子萌發(fā)的影響
試驗結果表明:濕度99%時,18 h后僅有少數萌發(fā),萌發(fā)率不及8%,濕度99%以下的各處理萌發(fā)率為零,而水滴中18 h萌發(fā)率已達76%,表明該病菌分生孢子的萌發(fā)需要液態(tài)水。
2.5.3 pH對病原菌分生孢子萌發(fā)的影響
試驗結果(圖12)表明,病菌分生孢子在pH 4~11范圍內均能萌發(fā),pH 3不萌發(fā),最適萌發(fā)pH為7,pH 4、10、11雖能萌發(fā),但芽管較其他pH的短。可見孢子萌發(fā)對pH要求不嚴格。
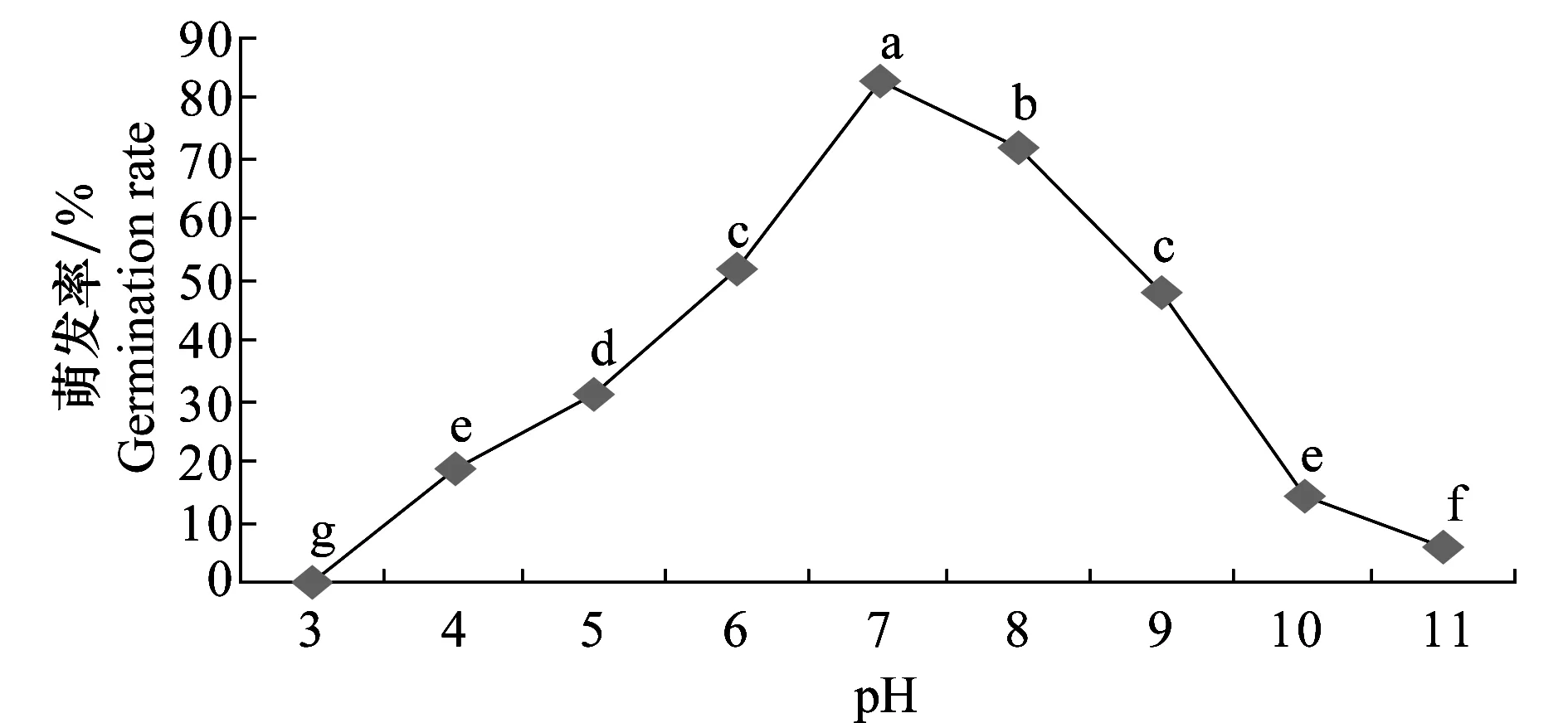
圖12 pH對馬鈴薯褐腐病菌分生孢子萌發(fā)的影響
2.5.4 營養(yǎng)液對病原菌分生孢子萌發(fā)的影響
分生孢子在葡萄糖液中的萌發(fā)最好(圖13),12 h萌發(fā)率已達72%以上,1∶5的葡萄糖液中高達92%,其次為馬鈴薯汁液各處理濃度,萌發(fā)率為51%~79%,顯著高于對照萌發(fā)率,說明該病菌侵染寄主后在適宜的溫濕度條件下可迅速蔓延,而土壤浸漬液各處理濃度萌發(fā)率和對照無顯著差異,表明土壤浸漬液對分生孢子的萌發(fā)沒有促進作用。
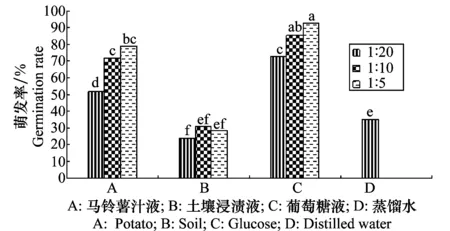
圖13 營養(yǎng)液對馬鈴薯褐腐病菌分生孢子萌發(fā)的影響
3 討論
通過形態(tài)學觀察結合分子生物學方法將甘肅省馬鈴薯褐腐病病原鑒定為馬鈴薯褐腐束梗霉[Doratomycesstemonitis(Pers.exFr.) F.J. Morton & G. Smith]。李金花于2007年報道[6]從采自甘肅張掖的病薯上分離到該菌,只對其形態(tài)進行了描述,未做致病性測定及生物學特性研究。白容霖于1999年報道此菌能侵染人參引起黑腐病[13],發(fā)病癥狀與侵染馬鈴薯癥狀相似:病斑呈橢圓形或不規(guī)則形凹陷, 黑褐色, 病斑邊緣明顯, 發(fā)病后期參根組織呈現黑朽狀,形成空腔。在濕度大的情況下, 病斑表面叢生紫黑色絨毛狀的病原菌的子實體。其描述的病原形態(tài)、大小均與本試驗的病原基本一致。以上報道的病菌是否與本研究的病原為同一種、是否存在生理分化現象,還有待進一步研究。還有文獻記載此菌有降解木聚糖、半纖維素以及纖維素的能力[14-15],可以從玉米、苜蓿的根際,甜菜、油菜、蠶豆的根以及多種植物的殘體上分離得到,可見該菌可以腐生形式存在于田間病殘體和土壤中。本研究從病原分離、致病性測定、病原鑒定及其生物學特性等方面進行了較系統(tǒng)的研究,是Doratomycesstemonitis引起馬鈴薯褐腐病在我國首次較系統(tǒng)的報道。
在早期文獻中,束梗孢目(Stilbellales)束梗孢科 (Stilbellaceae)細基束梗霉屬(Doratomyces)真菌被組合在Stysanus屬下,Hughes于1958年提出將屬名改為Cephalotrichum,后來 Morton 和 Smith于1963年建議將其名改為Doratomyces[13]。該屬內真菌的一個顯著特點是形成明顯的孢梗束,Doratomycesstemonitis(Pers.exFr.) F. J. Morton & G.Smith為該屬的模式種,是典型的無性型真菌。
生物學特性研究結果表明,低溫、干燥不利于該菌的生長,所以在低溫干燥的條件下貯藏馬鈴薯,可減少該病害的發(fā)生。此病原菌分離自儲藏期的馬鈴薯薯塊上,致病性測定表明傷口是病原侵染的主要途徑,而且在在采挖馬鈴薯薯塊過程中也發(fā)現部分表皮有傷口的薯塊也發(fā)生有此病害,所以農事操作時要盡量減少薯塊的機械損傷,以免造成傷口為病菌的侵染提供有利條件。
從碳源對菌絲生長的影響試驗結果來看,此病菌能利用多種碳源,僅有以L-山梨糖為碳源的培養(yǎng)基抑制該病菌的生長,而有研究報道[16]稱以L-山梨糖為唯一碳源時,必須添加有機氮菌株才能正常生長和產酶,L-山梨糖的作用可能是通過抑制了菌體蛋白或某些必需氨基酸、核苷酸的合成,從而影響了菌體生長和產酶的,并且這種影響發(fā)生在菌絲體指數生長期之前,而L-山梨糖對Doratomycesstemonitis的抑制機制是否與本報道一致,還有待于進一步研究。
[1]楊成德, 姜紅霞, 陳秀蓉, 等. 甘肅省馬鈴薯炭疽病的鑒定及室內藥劑篩選[J]. 植物保護,2012,38(6):127-133.
[2]魏周全, 宗世忠, 張廷義. 定西市馬鈴薯病害調查[J]. 中國馬鈴薯, 2005,19(2):124-126.
[3]文朝慧, 何蘇琴, 荊卓瓊. 甘肅省馬鈴薯壞疽病病原鑒定[J]. 植物保護,2012,38(5):40-45.
[4]費永祥, 張建朝, 邢會琴. 甘肅省馬鈴薯細菌性病害種類及1種新紀錄病害[J]. 河西學院學報,2010,26(2): 51-53.
[5]鄭軍慶, 趙多長, 韓曉榮, 等. 天水市馬鈴薯窖藏病害調查[J]. 中國蔬菜,2011(1): 33-34.
[6]李金花, 柴兆祥, 王蒂, 等. 甘肅馬鈴薯貯藏期真菌性病害病原菌的分離鑒定[J]. 蘭州大學學報, 2007, 42(3): 39-42.
[7]戴芳瀾. 中國真菌總匯[M]. 北京: 科學出版社, 1979.
[8]方中達. 植病研究方法[M]. 第3版. 北京: 中國農業(yè)出版社,1998.
[9]魏景超. 真菌鑒定手冊[M]. 上海: 上海科學技術出版社, 1979.
[10]張俊忠. 東祁連山高寒草地土壤真菌多樣性研究[D]. 蘭州:甘肅農業(yè)大學, 2010:35-36.
[11]張小媛, 何紅, 胡漢橋, 等. 紅海欖赤斑病病原鑒定及其生物學特性研究[J]. 植物病理學報, 2009, 39(6): 584-591.
[12]王艷, 陳秀蓉, 王引權, 等. 甘肅省當歸褐斑病菌Septoriasp.生物學特性及其營養(yǎng)利用研究[J]. 中藥材,2009,32(4):478-482.
[13]白容霖, 劉偉成. 人參黑腐病病原鑒定[J]. 特產研究, 1999(4):42-43.
[14]耿月華. 中國土壤暗色絲孢菌五個疑難屬的形態(tài)學和分子系統(tǒng)學分析[D]. 泰安: 山東農業(yè)大學, 2011:89.
[15]宋偉. 中國秦嶺太白山及周圍地區(qū)土壤中的暗色絲孢菌多樣性研究[D]. 泰安: 山東農業(yè)大學, 2008:106.
[16]郝學財.纖維素酶高產菌選育及發(fā)酵條件的研究[D]. 無錫: 江南大學,2006.
[17]孫俊, 劉志恒, 黃欣陽, 等. 辣椒褐斑病菌生物學特性研究[J]. 植物保護,2009,35(5):109-113.
IdentificationandbiologicalcharacteristicsofSolanumtuberosumbrownrotcausedbyDoratomycesstemonitis
Chen Taixiang, Chen Xiurong, Yang Chengde, Zhu Haibo, Wang Hanqi, Bian Jing
(PrataculturalCollegeofGansuAgriculturalUniversity;KeyLaboratoryofGrasslandEcosystem,MinistryofEducation,PrataculturalEngineeringLaboratoryofGansuProvince,Sino-U.S.CenterforGrazinglandEcosystemSustainability,Lanzhou730070,China)
The identification and biological characteristics of the pathogen isolated from brown rot ofSolanumtuberosumwere studied. The pathogen was identified asDoratomycesstemonitis(Pers.exFr.) F.J. Morton & G. Smith based on morphological characteristics as well as rDNA-ITS sequences. The optimum temperature for mycelial growth was 25-30 ℃ at pH 6. The pathogen could use monosaccharide, disaccharide and polysaccharide; the best carbon source was sucrose, and the best nitrogen source was glycine. Illumination had no obvious promotion effect on mycelial growth. The optimum temperature for sporangium sprout was 25 ℃ at pH 7. Spores could germinate only in water drop. The filtrate of potato and glucose solution had obvious promotion effect on spore germination. Soil solution had no effects on spore germination.
Solanumtuberosum;Doratomycesstemonitis; pathogen identification; biological characteristics
2013-11-01
:2014-02-16
甘肅省農牧廳項目
S 435.32
:ADOI:10.3969/j.issn.0529-1542.2014.05.005
* 通信作者 E-mail: chenxiurong@gasu.edu.cn
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
