萊州灣夏季大型底棲動物群落結構特征及其與歷史資料的比較
2014-08-14 01:21:22劉曉收趙瑞華爾路璐張志南
海洋通報 2014年3期
關鍵詞:環境
劉曉收,趙瑞,華爾,路璐,張志南
(中國海洋大學 海洋生命學院,山東 青島 266003)
萊州灣位于渤海南部,山東半島北部,海岸線長90多km,面積4 600多km2,不僅是山東省重要漁鹽生產基地,亦有豐富的石油和天然氣蘊藏。萊州灣受到黃河、小清河和山東半島沿岸十幾條入海河流的影響,是渤海漁業生物資源的重要產卵場和棲息地。近幾十年來,隨著沿岸經濟的迅速發展和涉海人類活動(捕撈、養殖、石油和天然氣開采等)的增加以及黃河徑流量的急劇減少所帶來的水層和沉積環境的改變,包括萊州灣在內的渤海生態系統的結構和功能正經歷著快速變化和退化的趨勢(金顯仕等,1998)。
底棲生物作為一類活動能力相對較弱的海洋生物類群,在受到人類活動的直接影響后,其生物種類及群落組成都將發生變化(韓慶喜等,2011)。渤海由于其重要的經濟價值和重污染的環境特征而一直倍受關注,建國以來已進行了多次包括大型底棲動物調查在內的綜合性生物生態調查(孫道元等,1991)。這些研究主要側重在黃河口水下三角洲及其鄰近海域、渤海中南部和渤海灣,鮮見針對萊州灣大型底棲動物的研究(周紅等,2010)。水質環境和生物多樣性綜合評價結果表明,萊州灣2002年5月和8月處于良好水平。值得指出的是,生物多樣性綜合評價僅采用了浮游植物生物多樣性、浮游動物生物多樣性和葉綠素a,沒有涉及底棲生物,而底棲生物在海洋生態系統健康評估中起重要作用。因此,評價結果存在一定的局限性(馬紹賽等,2006)。
本文通過研究萊州灣9個典型站位大型底棲動物的豐度、生物量和種類組成,分析大型底棲動物在萊州灣的群落結構及生物多樣性,并探討其與環境因子的相互關系,同時與歷史資料進行對比,以期了解萊州灣生態環境現狀和年際變化,為保護其生態資源實現可持續利用及生態系統健康評價提供科學依據。
1 材料和方法
1.1 研究海域和現場取樣
于2009年6月1-5日在渤海萊州灣海域9個站位進行了大型底棲動物的定量取樣,站位分布見圖1和表1。采用0.1m2的箱式采泥器,在取樣站位采集未受擾動的沉積物樣品,每個站位取3個平行樣,現場使用0.5mm孔徑的網篩分選大型底棲動物,將生物標本及殘渣全部轉移至樣品瓶,并用5%福爾馬林固定。同時在采樣點取一定量的表層沉積物,用于沉積物粒度、葉綠素等環境因子的分析。以上用于測定非生物環境因子的樣品立即放入-20℃低溫冰柜,冷凍保存。水層環境因子包括水深、表溫、表鹽、底溫、底鹽等,現場使用YSI600XLM-M水質分析儀測得。水體透明度使用透明度盤測定。
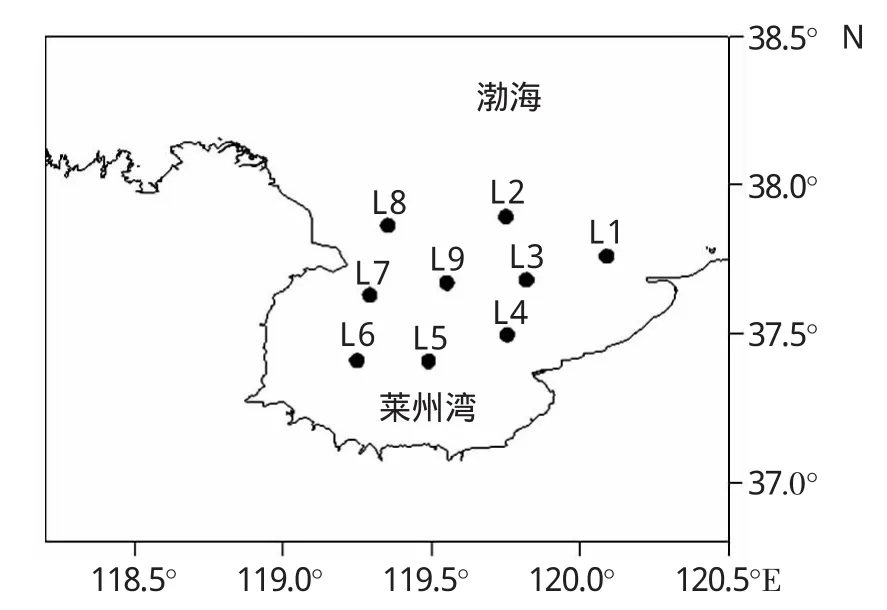
圖1 萊州灣大型底棲動物取樣站位圖
1.2 室內分析
1.2.1 沉積物環境因子的分析
沉積物環境因子包括粒度、有機質和葉綠素含量。沉積物粒度使用激光粒度分析儀測定。有機質含量參照海洋監測規范使用重鉻酸鉀-硫酸亞鐵還原法測定。葉綠素含量參照海洋調查規范和劉暉等使用熒光分光光度法測定(國家海洋局,1991;劉暉等,1998)。
1.2.2 生物樣品分析
用1‰的虎紅溶液對篩選所得的大型底棲動物標本和殘渣進行染色,靜置24 h之后,用普通解剖鏡進行分選鑒定并記數,其中殘體不計入豐度的統計;分類記數后的標本用濾紙吸干固定液,用感量為0.000 1 g的電子天平稱重,鑒定、稱重等均按 《海洋調查規范》 (GB/T 12763.6—2007)進行。
1.3 數據處理
對群落及環境數據的分析采用PRIMER 6.0軟件包中包含的一系列多元統計程序進行。群落結構分析基于種豐度矩陣,經雙平方根轉換、Bray-Curtis相似性計算進行組平均連接的等級聚合聚類(CLUSTER)和非度量多維標度排序(MDS),以此實現對群落的劃分(周紅等,2003)。
用PRIMER軟件得到的單變量有:群落種數(S)、豐富度(D)、香農-威納指數(Shannon-Wiener)(H′)和均勻性指數 (J)。群落結構與環境變量的關系采用BIOENV分析,找出單一環境變量與群落結構的相關關系及與群落結構形成最佳匹配(最大相關)的環境變量組合。采用ABC曲線,即豐度生物量比較曲線,分析大型底棲動物群落受污染或其它因素擾動的狀況(Warwick etal,1986)。
2 結果與討論
2.1 環境因子
2.1.1 水層環境因子
各站位水層環境因子見表1。本次調查的9個站位都位于水深較淺(3~16.8m)的海域,其中L7站最淺,離岸最近,水深只有3 m,L2站16.8 m,離岸最遠;透明度變化范圍為1~4.8m,L6站最小為1 m,L7站透明度也較小,而離岸最遠的L2站透明度最大。表溫為19.02℃~23.64℃,各站位差距不大,但底溫為15.81℃~23.04℃,各站位有較明顯變化,其變化趨勢與表溫有所不同;表鹽27.63~30.84,L7站最低;底鹽28.18~31.5,變化趨勢較表鹽也有所不同。
2.1.2 沉積物的粒度特征和沉積物類型
沉積物是通過物理的、化學的和生物沉積作用過程所形成的海底沉積物的總稱。沉積環境是底棲動物的棲息地,直接影響著底棲動物生物量的大小和群落的組成與分布。而沉積物粒度組成、分選狀況都是影響底棲動物的重要環境因子。本研究利用中值粒徑(Mdφ)、分選系數σi和四分位偏態Ski比較說明各站位沉積物粒度狀況。
由表2中值粒徑的變化來看,L4站明顯高于其他站位,而L4站的底質類型為砂,也與其他站位明顯不同,另外,L2、L5、L7站的底質類型分別為砂質粉砂、粉砂質砂,其中值粒徑值相近,略大于剩余幾個底質類型為粘土質粉砂的站位。由于底質類型的確定是由沉積物中粒級含量超過20%的粒級命名的,且含量多的在后,少的在前。可見粘土質、砂質含量越高,中值粒徑越小,即中值粒徑的值與粘土和粉砂(Y+T)的含量呈負相關。

表1 萊州灣各取樣站位的水層環境因子
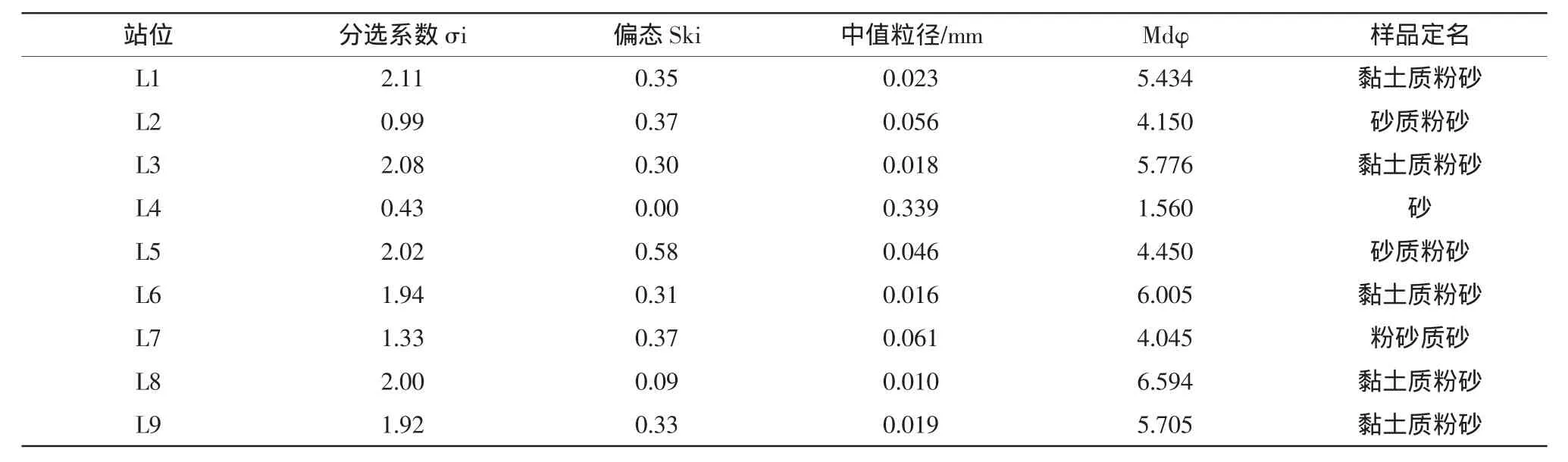
表2 萊州灣各取樣站位沉積物粒度特征和類型
2.1.3 萊州灣沉積物中有機質含量
有機質含量各站位差距明顯(圖2),其中L9站最高。與沉積物底質類型比較發現,有機質含量較高的站位如L1、L3、L6、L8、L9號站底質類型都為粘土質粉砂,最低的L4站底質類型為砂質,可見底質顆粒越細,有機質含量越高。

圖2 萊州灣取樣站位沉積物中有機質含量
2.1.4 萊州灣沉積物葉綠素a及脫鎂葉綠酸a含量
沉積物中葉綠素的含量關系到底棲動物的食物來源和底質環境的質量,脫鎂葉綠酸a(Pha)的含量預示著底質中死亡植物的量。
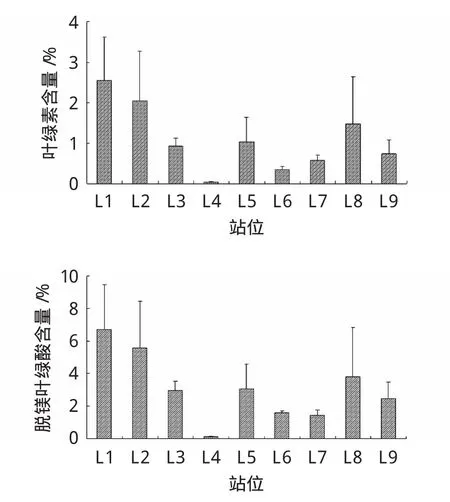
圖3 萊州灣取樣站位沉積物葉綠素a(Chla)及脫鎂葉綠酸a(Pha)含量
萊州灣沉積物中葉綠素a含量為0.04~2.55μg/g,脫鎂葉綠酸a含量為0.14~6.74μg/g,各站位差異較顯著(圖3)。Chla含量整體較低,Pha含量相對較高,二者變化趨勢基本相同,Chla含量 L1>L2>L8>L5>L3>L9>L7>L6>L4,而 Pha 含量 L1>L2>L8>L5>L3>L9>L6>L7>L4。分析影響沉積物中葉綠素a和脫鎂葉綠酸a含量高低的因素得知,其含量除了受海水中營養鹽高低影響外,海水的透明度和沉積物環境的穩定性也是重要的因素。L4站有機質含量最低,其葉綠素a和脫鎂葉綠酸a含量也是最低。L6、L7站海水透明度較低,因而這些站點沉積物中葉綠素a和脫鎂葉綠酸a含量普遍較低。由于各站位均位于近海,受河流徑流、養殖區等多方面因素綜合影響,其變化趨勢較為復雜。
2.2 大型底棲動物
2.2.1 類群組成

圖4 萊州灣大型底棲動物各類群豐度、生物量及種類比例圖
經鑒定,本研究共采集到大型底棲動物96種,包括節肢動物甲殼類、軟體動物、環節動物、棘皮動物、螠蟲等,各類群平均豐度、生物量及其所占比例見圖4。從豐度來看,大型底棲動物總平均豐度為1 902.21 ind./m2,優勢類群為軟體動物,其平均豐度1 241.85 ind./m2,占總平均豐度的65%;其次為節肢動物430.74 ind./m2,占總平均豐度的23%;環節動物203.33 ind./m2,占總平均豐度的11%;其他種類26.28 ind./m2,占1%。從生物量看,總平均生物量為8.30 g/m2,雖優勢類群仍是軟體動物(2.95 g/m2,占36%)及節肢動物甲殼類(2.08 g/m2,占25%),但紐蟲(0.77 g/m2,占9%)和棘皮類(0.95 g/m2,占11%)比重明顯增加,主要由于紐蟲和棘皮動物個體比較大。從物種數來看,節肢動物(40種,占42%)和軟體動物(32種,占33%)種數較多。另外,在L4站的3個平行樣中均發現了白氏文昌魚的存在,平均豐度16.67 ind./m2,平均生物量0.08 g/m2。
2.2.2 生物量和豐度的水平分布
萊州灣9個站位大型底棲動物平均豐度以L9站 (5 163.33±3 516.96 ind./m)2最大,L3站(4 406.67±1 700.36 ind./m)2次之,L8站 (266.67±45.09 ind./m)2最少(圖5)。有機質含量較低的幾個站位如L2、L4、L7站平均豐度也較低,有機質含量較高的L1、L3、L6、L9站中大型底棲動物平均豐度也較高,說明沉積物中有機質含量與底棲動物豐度密切相關,而L8站平均豐度可能受多方面因素的影響。此外,優勢種方面,L1、L2、L3、L8、L9站雙殼類占絕對優勢,如微形小海螂、紫殼阿文蛤、江戶明櫻蛤等;L4站,多毛類的奇異稚齒蟲、寡鰓齒吻沙蠶、小頭蟲占優勢;L6站甲殼類三葉針尾漣蟲、纖細長漣蟲、絨毛近方蟹占優勢;L7站甲殼類二齒半尖額漣蟲和小頭彈鉤蝦占優勢。
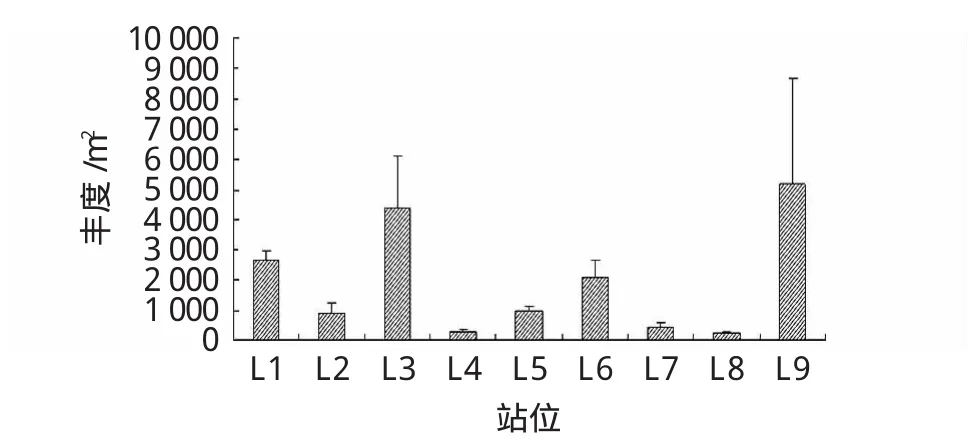
圖5 萊州灣取樣站位大型底棲動物平均豐度
平均生物量與平均豐度相比出現了變化(圖6),首先L3站18.31±20.76 g/m2為最大,其中貢獻最大的為異足倒顎蟹(34%)、紐蟲(21%)等體形較大的物種;另外變化較大的是豐度平均值只有 300±36.07ind./m2的 L4站,生物量平均值(16.52±0.89 g/m2)卻僅次于L3站,主要也是由于其某些樣品體型較大的緣故,如異足索沙蠶(占34%),紫蛤(占28%)等。L5站中對平均生物量貢獻最大的物種是棘刺錨參(59%),L6站(14.05±7.43 g/m2)中紅帶織紋螺(25%)、橄欖胡桃蛤(17%)、仿盲蟹(11%)貢獻最大。
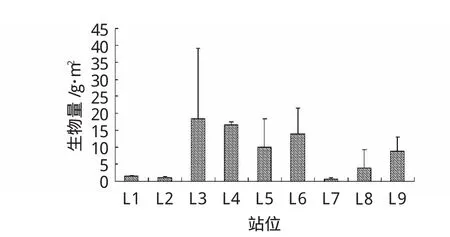
圖6 萊州灣取樣站位大型底棲動物平均生物量
2.3 群落結構分析
對各站位的大型底棲動物種類豐度進行平方根轉化,做出Bray-Curtis相似性矩陣,以此進行CLUSTER聚類和MDS標序(圖7)。在萊州灣所研究海域,大型底棲動物在30%的相似性水平上大致可劃分出3個組。

圖7 萊州灣大型底棲動物群落結構CLUSTER聚類和MDS標序圖
組1:為L2站和L8站,2個站相似性約45%。二者豐度較其他站位小,優勢種為微形小海螂、紫殼阿文蛤等小型雙殼動物,此群落生活在離海岸較遠的海域,水深13~16.8m,底溫15.81℃~16.87℃,底鹽30.86~30.33,底質類型為砂質粉砂和粘土質粉砂。
組2:L1、L3、L5、L9、L6站,5個站相似性約40%。這幾個站位物種豐度較大,優勢種有微形小海螂、紫殼阿文蛤、江戶明櫻蛤等雙殼類和三葉針尾漣蟲、纖細長漣蟲、絨毛近方蟹等甲殼類,此群落生活在灣內,水深10~15.5m,底溫16.65℃~21.42℃,底鹽28.5~31.5,底質類型主要有粘土質粉砂和砂質粉砂。
組3:L4、L7號站,相似性約30%。豐度小,優勢種有寡鰓齒吻沙蠶、奇異稚齒蟲等多毛類和二齒半尖額漣蟲、小頭彈鉤蝦等甲殼類,該群落生活在近岸海域,水深3~13m,底溫19.5℃~23.04℃,底鹽28.18~31.32,底質類型主要是砂和粉砂質砂。
2.4 群落結構的多樣性分析
2.4.1 種類組成
本次研究共采集到大型底棲動物96種,從豐富度指數來看,L1>L5>L6>L3>L9>L4>L8>L2>L7,L1站豐富度最高,為5.7;L7最低,為2.6。
2.4.2 多樣性指數的平面分布
多樣性指數(基于豐度數據)如圖8所示。
9個站位的香農-威納指數大小順序為L5>L6>L4>L1>L8>L7>L2>L3>L9,L9站的香農-威納指數最小,多樣性相對較低,主要由于L9站采集到了大量的紫殼阿文蛤,平均豐度為3 893 ind./m2,占本站位總平均豐度的75%。L5站雖豐富度指數次于L1站,但其均勻度指數較高,其優勢種江戶明櫻蛤、三葉針尾漣蟲、小莢蟶、口蝦蛄分別占21%、12%、9%、7%,沒有占絕對優勢的物種,故其多樣性指數為最高。此外,研究海域香農-威納指數為1.143~2.923,平均值為2.1,蔡立哲等結合群落結構的變化以及有機質等參數的分析(蔡立哲等,2002),將多樣性指數污染評價范圍分為5級,即無底棲動物為嚴重污染;H′值小于1,重度污染;H′值在1~2之間,中度污染;H′值在2~3之間,輕度污染;H′值大于3,清潔(蔡立哲等,2002)。據此,L2、L3、L7、L8、L9 站 H′值都在1~2之間,屬于中度污染;其他站位H′值都在2~3之間,屬輕度污染。

圖8 萊州灣取樣站位大型底棲動物多樣性指數
2.5 ABC曲線的分析
由圖9可知,L1、L2、L3、L7、L9站位的豐度k-優勢度曲線與生物量k-優勢度曲線出現交叉,說明其所在海域環境受到中等程度的擾動;L4、L6站豐度k-優勢度曲線和生物量k-優勢度曲線未重合,但生物量曲線起點不高,即優勢不是很明顯,且與豐度曲線距離較近,顯示出一種大型底棲生物群落傾向于受到中等程度的污染的狀況;L5、L8站生物量k-優勢度曲線始終位于豐度k-優勢度曲線之上,且優勢度明顯,表明大型底棲動物群落尚未受到干擾。而按照之前H′值判斷污染的方法中,L2、L3、L7、L8、L9站屬于中度污染;其他站位H′值都屬輕度,所以判斷某海域是否污染需要綜合判斷各方面的因素。
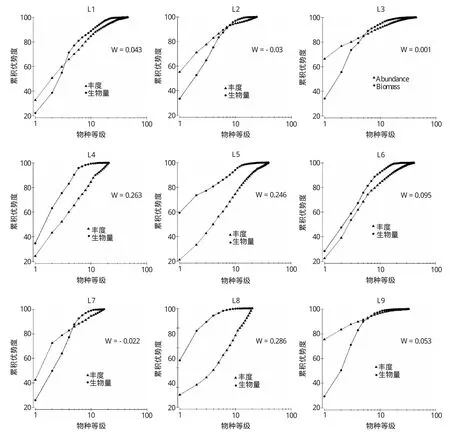
圖9 萊州灣各取樣站位的ABC曲線(▲:豐度k-優勢度曲線,●:生物量k-優勢度曲線)
2.6 環境因子與豐度生物量的關系
對各站的豐度和生物量數據進行平均處理,經平方根轉化,作出Bray-Curtis相似性矩陣,與對應的環境參數進行BIOENV分析,找出與豐度和生物量最為匹配的環境因子(表3、4)。
表3分析表明,與萊州灣大型底棲動物豐度最匹配的環境因子組合為水深(m)、底溫℃、中值粒徑mm、有機質(%)、葉綠素含量(μg/g)5個因素,其相關系數最大,為0.766。除水層環境因子外,大型底棲動物豐度主要受沉積物類型及沉積物有機質和葉綠素含量的綜合影響,豐度較高的L9、L3、L6站,都是位于粘土-粉砂含量較高的、有機質含量較高的海域。L4站則位于粘土-粉砂含量很低的砂質底質中,其有機質和葉綠素a含量都比較低,故其豐度也比較小。
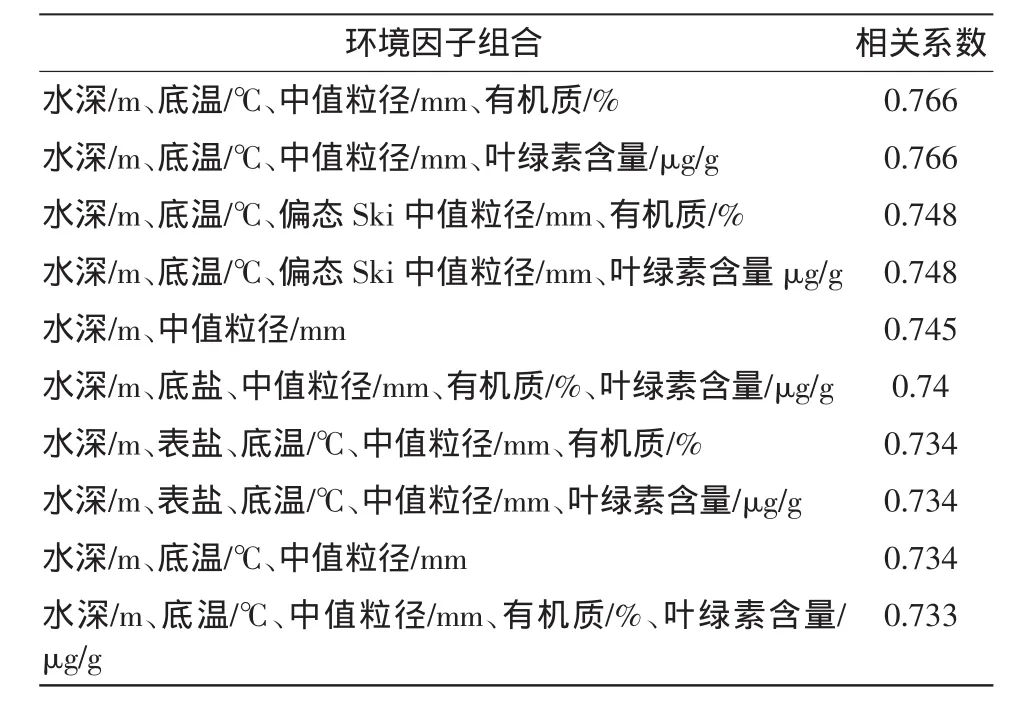
表3 豐度與環境因子匹配關系
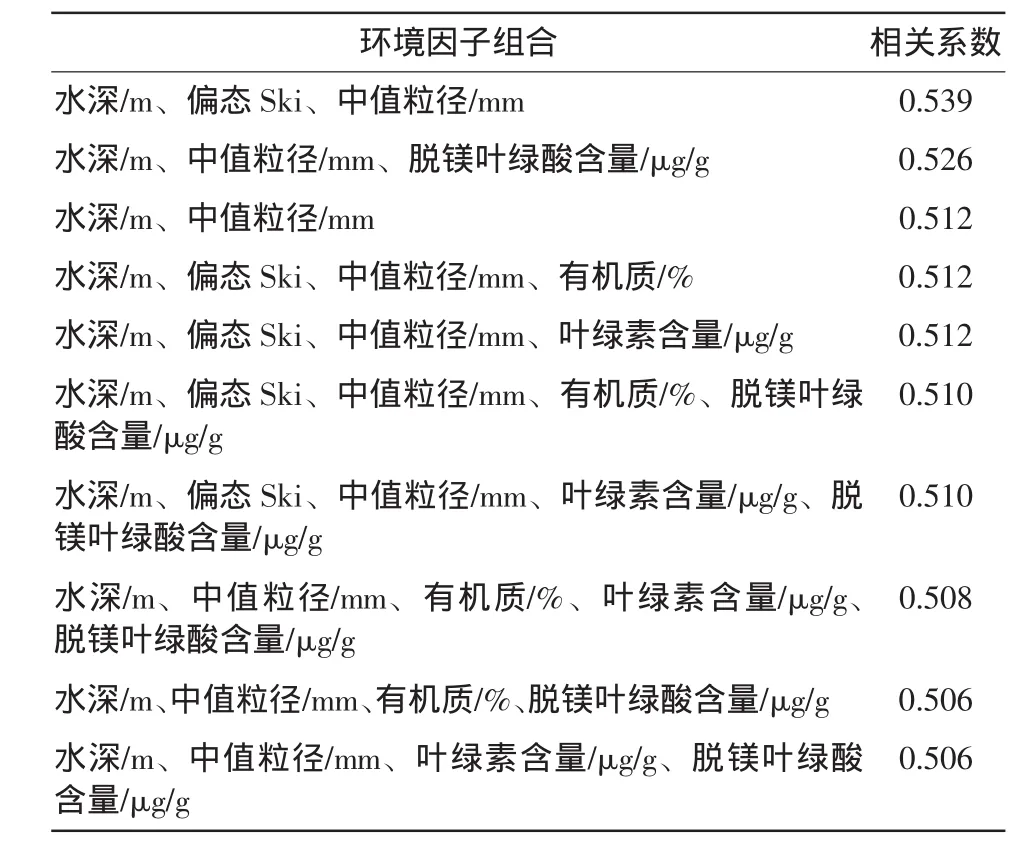
表4 生物量與環境因子匹配關系
表4分析表明與萊州灣大型底棲動物生物量最匹配的環境因子組合為水深(m)、偏態Ski、中值粒徑mm,相關系數為0.539。可見大型底棲動物的生物量與底質類型有密切聯系,同樣的水深情況下,中值粒徑越小,偏態系數越低,則生物量越小,如L4站和L8站,同樣位于13m深處,L8中值粒徑(0.01mm)與偏態系數(0.09)都比較小,L4站雖偏態系數(0)較小,但中值粒徑(0.339)較大,故L8站生物量較低。
2.7 與歷史數據的對比
通過與20世紀80年代中美黃河口聯合調查及90年代渤海生態系統動力學與生物資源可持續利用國家自然基金重大項目及2006年秋季渤海萊州灣大型底棲動物取樣調查所獲得數據進行對比發現(表5)。研究海域沉積物有機質含量在90年代明顯高于80年代、2006年和本研究的數據;沉積物粒度方面,本研究的中值粒徑值低于歷史數據,說明沉積物粉砂粘土含量升高,雖然萊州灣屬于遠離黃河口的低沉積速率區(張志南等,1990a),但長期以來仍有大量泥沙沉積。大型底棲動物的豐度與80年代、90年代及2006年相比均有所上升,而生物量和種數減少,生物多樣性下降,說明近年來人類活動的加劇致使某些物種在該海域消失,而某些機會種(特別是小型多毛類)得以大量繁衍(周紅等,2010)。
除了豐度與生物多樣性以外,萊州灣大型底棲動物的群落結構也發生了明顯變化,這種變化主要體現在優勢種的小型化趨勢,即小個體的多毛類、雙殼類和甲殼類取代大個體的棘皮動物和軟體動物(周紅等,2010)。1980年代萊州灣的生物量很高,穴居型的雙殼類和棘皮動物在數量和生物量上均占明顯優勢(張志南等,1990b),形成一個以凸殼肌蛤-心形海膽為優勢種的群落(孫道元等,1989;孫道元等,1991)。到1990年代在萊州灣豐度和生物量很高的心形海膽(Echinocarium cordatum)和凸殼肌蛤 (Musculista senhousia),被較小的紫殼阿文蛤(Alveniusojianus)和銀白齒緣殼蛞蝓 (Yokoyamaia argentata)取代 (韓潔等,2001),而20世紀以后,除了紫殼阿文蛤繼續占優勢地位外,更小的種類小亮櫻蛤、微型小海螂等相繼成為優勢種。1990年代以后,對生物量貢獻很大的大型種類已在萊州灣失去優勢,可以解釋灣內生物量的下降(表5);而周紅等(2010)對萊州灣2006年的研究與本研究類似,都是體型較小的多毛類、雙殼類和甲殼動物占優勢。
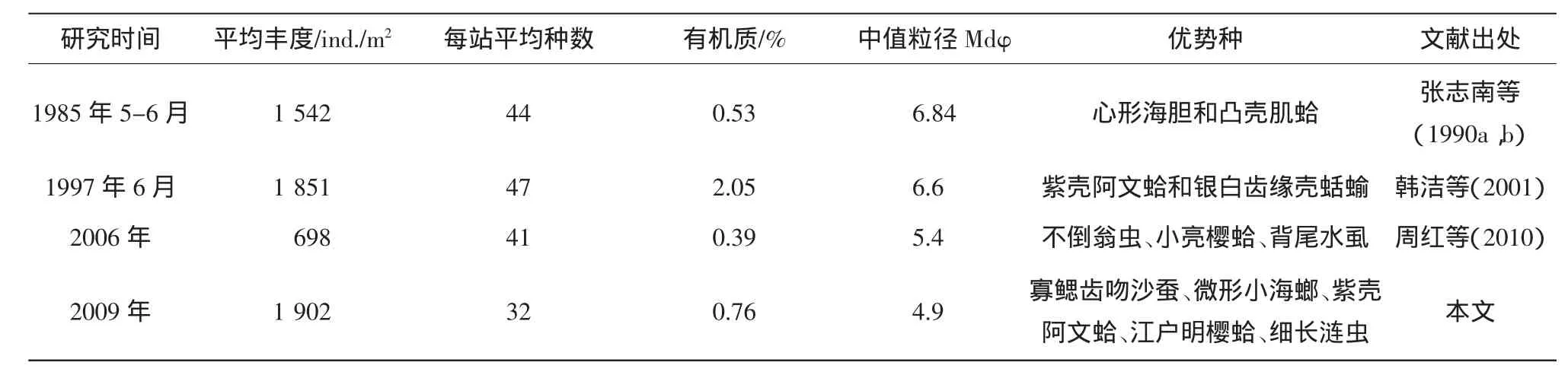
表5 萊州灣大型底棲動物及環境因子歷史數據對比
3 結論
(1)豐度和生物量
本次調查共鑒定出大型底棲動物96種,類群包括軟體動物、節肢動物甲殼類、環節動物、棘皮動物、脊索動物、扁形動物、帚形動物、螠蟲動物共8類,大型底棲動物平均豐度為1 902.21 ind./m2,平均生物量為8.30 g/m2。其中軟體動物在豐度上占絕對優勢,占65%,主要是采集到了大量的紫殼阿文蛤的緣故,而生物量方面,軟體動物所占比例仍居第一位,但比例下降到了36%,體形較大的棘皮動物和紐蟲所占比例分別上升到了12%和9%。L9站豐度(5163.33 ind./m2)最大,L3站(4 406.67 ind./m2)次之,L8站(266.67 ind./m2)最少。生物量方面L3站18.31 g/m2居首,L4站(16.52 g/m2)次之,L7站最小。
(2)群落結構
CLUSTER聚類和MDS標序圖顯示,依據豐度所研究海域共也劃分為3個組,一個是離海岸較遠水深較深的的L2和L8站,其相似性達到了45%;另一個是L1、L3、L5、L9、L6站,相似度為40%,比較而言此群落平均豐度較大;最后一個是水深最淺的L4、L7站,相似度約30%。
(3)與環境因子的關系
本文所研究海域底質類型包括粘土質粉砂、粉砂質砂、砂質粉砂、砂4種,沉積物中(T+Y)含量與中值粒徑呈負相關,而與有機質含量成正相關,即顆粒粒級越細,沉積物中有機質含量越高,也越有利于底棲生物生存。葉綠素a與脫鎂葉綠酸a也是影響大型底棲生物生態結構的重要因素,其含量除了受海水中營養鹽高低影響外,也受海水的透明度和沉積物環境的穩定性的影響。經BIOENV分析,與大型底棲動物豐度匹配最佳的環境因子組合是水深、底溫、中值粒徑、有機質和葉綠素含量,與生物量匹配最佳的環境因子組合是水深、偏態、中值粒徑。
(4)生物多樣性
通過多樣性指數中的單個變量分析和ABC曲線分析,L5站受到的擾動最小,H′值最大,即生物多樣性最高;豐度最大的L9由于污染等原因,紫殼阿文蛤過于集中,多樣性最低。此外本海域L1、L2、L3、L7、L9站所處位置受到中等程度的污染;L4、L6站有受到中度污染的趨勢;L5、L8站擾動較小,環境還沒有受到很明顯的污染。另外,在L4站發現白氏文昌魚的存在。萊州灣東部海區文昌魚的出現,說明在相似的海況底質條件下,輔以適當保護措施,其群體在我國近海得到生存繁衍是可能的(高天翔等,2000)。
(5)與歷史數據的對比
近二十年來,萊州灣海域大型底棲動物的豐度有所增加,但生物量和物種數目有所減少,生物多樣性下降。造成該變化的主要原因是優勢種的小型化趨勢,即小型多毛類、雙殼類和甲殼類取代大個體的棘皮動物和軟體動物。
Warwick,R M,1986.A new method for detecting Pollution effects on marine macrobenthic communities.Marine Biology,92:557-562.
Zhou H,Zhang ZN,Liu X S,etal,2007.Changes in the shelf macrobenthic community over large temporal and spatial scales in the Bohai Sea,China.Journal of Marine Systems,67:312-321.
蔡立哲,馬麗,高陽,等,2002.海洋底棲動物多樣性指數污染程度評價標準的分析.廈門大學學報(自然科學版),41(5):641-646.
高天翔,張宏義,姜衛蔚,等,2000.萊州灣東部水域出現的文昌魚幼體的初步調查.海洋湖沼通報,3:20-23.
國家海洋局,1991.海洋監測規范.北京:海洋出版社.
韓潔,張志南,于子山,2001.渤海大型底棲動物豐度和生物量的研究.青島海洋大學學報,31(6):889-896.
韓慶喜,李寶泉,韓秋影,等,2011.漁業捕撈對威海港附近海域底上大型底棲群落結構影響的初步研究.海洋通報,30(2):121-126.
金顯仕,唐啟升,1998.渤海漁業資源結構、數量分布及其變化.中國水產科學,5(3):18-24.
劉暉,吳以平,高尚德,等,1998.蝦病暴發前蝦池中一些環境因子的變化.青島海洋大學學報,28(3):377-382.
馬紹賽,趙俊,陳碧鵑,等,2006.萊州灣漁業水域生態環境質量分析與綜合評價研究.海洋水產研究,27(5):13-16.
孫道元,劉銀城,1991.渤海底棲動物種類組成和數量分布.黃渤海海洋,9(1):42-49.
張志南,谷峰,于子山,1990a.黃河口水下三角洲海洋線蟲空間分布的研究.海洋與湖沼,21(1):11-19。
張志南,李永貴,圖立紅,等,1989.黃河口水下三角洲及其鄰近水域小型底棲動物的初步研究.海洋與湖沼,20(3):197-208.
張志南,圖立紅,于子山,1990b.黃河口及鄰近水域大型底棲動物的初步研究(I)生物量.青島海洋大學學報,20(1):37-45.
張志南,圖立紅,于子山,1990c.黃河口及鄰近水域大型底棲動物的初步研究(II)與沉積環境的關系.青島海洋大學學報,20(2):43-52.
周紅,華爾,張志南,2010.秋季萊州灣及鄰近海域大型底棲動物群落結構的研究.中國海洋大學學報,40(8):80-87.
周紅,張志南,2003.大型多元統計軟件PRIMER的方法原理及其在底棲群落生態學中的應用.中國海洋大學學報,33(1):58-64.
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
瘋狂英語·新策略(2019年9期)2019-10-17 01:51:34
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
濰坊學院學報(2017年2期)2017-04-20 08:44:31
中國環境監察(2016年5期)2016-10-24 05:25:52
中國商論(2016年33期)2016-03-01 01:59:38
