準噶爾荒漠6種類短命植物生物量分配與異速生長關系
2014-11-07 01:46:32陶冶張元明
草業學報 2014年2期
陶冶,張元明
(1.中國科學院干旱區生物地理與生物資源重點實驗室 中國科學院新疆生態與地理研究所,新疆 烏魯木齊 830011;2.中國科學院大學,北京 100049)
類短命植物(ephemeroid plant)是指利用早春季或夏初相對充足的水分在短時間里迅速完成生活史,地上部分當年死亡、而地下部分為多年生的一類特殊植物類型[1-2]。類短命植物與短命植物(ephemeral plant)一道成為準噶爾荒漠春季與初夏維持地表穩定、物種多樣性及生態系統穩定性的重要貢獻者[1,3]。準噶爾荒漠是短命和類短命植物在中亞干旱區分布的最東端,因此,這類植物也是我國荒漠植物區系中重要而獨特的組成部分[1-2]。
生物量是植物的基本生物學特征和功能性狀之一,是物質和能量積累的基本體現[4-5]。生物量分配指植物生長發育過程中所同化的資源用于根、莖、葉、花、果等各器官的比例,也就是各器官累積的資源在總資源中所占的比例,是植物生殖與生存平衡的結果。生物量分配主要受植物的遺傳特性、環境條件和個體大小的影響[6-7],其分配模式在不同生活型植物間差異較大[8]。目前關于荒漠地區短命植物、一年生草本、多年生草本植物等都有較多的研究[8-10],但關于類短命植物這一特殊生活型植物生物量分配的研究仍相當匱乏[11],對其生物量分配的特點和規律仍知之甚少。
異速生長關系,即相關生長關系(allometric relationship),指生物體的某些生物學特征(如個體形態指標、器官生物量等)與個體大小(重量)之間的冪函數關系。這種規律在生物界普遍存在,與尺度無關[12-16]。異速生長關系通過對數化大大減弱了單純使用生物量學特征的絕對量或比例系數所帶來的波動性,能揭示植物器官生物量分配特征間的與尺度無關的內在規律[17-18]。目前,異速生長關系在刻畫生物體構件結構與功能特征上得到了大量應用,其中以植物體構件(器官)生物量間的相關生長關系研究最為常見[17]。通過代謝理論和分型網絡模型的理論預測均表明,無論有次生組織還是無次生組織的植物,其地上、地下生物量間均為等速生長關系(冪指數為1)[12-16]。
類短命植物是一個生活型的概念,其包含不同科、屬的不同物種[1-2]。那么同為短命植物類群,不同物種是否具有一致或相似的生物量分配比例?它們的器官生物量間是否具有一致的相關生長關系?為解決上述科學問題,我們選擇了準噶爾荒漠常見的6種類短命植物:百合科(Liliaceae)的小山蒜(Alliumpallasii)、棱葉韭(Alliumcaeruleum)、粗柄獨尾草(Eremurusinderiensis)、黑鱗頂冰花(Gageanigra)、伊犁郁金香(Tulipailiensis),以及菊科(Asteraceae)的細葉鴉蔥(Scorzonerapusilla)作為研究對象。這6個物種既有不同科的植物,又有同科不同屬的和同屬不同種的植物。通過對其生物量分配和異速生長關系的對比研究,期望找出類短命植物生物量分配的一些特點及器官生物量分配的協同變異關系,為深入了解該類生活型植物的生存策略與生態功能提供科學依據,并為不同生活型植物構件功能特征的對比研究提供參考。
1 材料與方法
1.1 研究區概況
準噶爾荒漠位于新疆北部,其中心為我國最大的固定和半固定沙漠——古爾班通古特沙漠(44°11′~46°50′ N,84°31′~91°20′ E,4.88×104km2)。該沙漠年均蒸發量>2000 mm,而年均降水量普遍不超過150 mm,沙漠腹地僅有70~100 mm,且主要集中于冬春季;年均溫6~10℃,極端高溫為40℃以上,≥10℃的年積溫可達3000~3500℃,空氣相對濕度平均50%~60%,5-8月通常<45%[19-20]。準噶爾荒漠是我國唯一一個受北冰洋氣流影響的荒漠區,其物種組成和豐富度與我國其他荒漠地區差異明顯[1]。梭梭(Haloxylonammodendron)、白梭梭(Haloxylonpersicum)、心葉駝絨藜(Ceratocarpusewersmanniana)、假木賊(Anabasis)等灌木是準噶爾荒漠植物群落的主要建群種。該地區具有豐富的短命和類短命植物,是早春荒漠生態系統的重要組成部分;同時,準噶爾荒漠也是該類群植物在中亞干旱區分布的最東端[2]。
1.2 植物樣品采集與處理
所有植物樣品均于2010年5月底類短命植物開花末期采集,6種植物均處于相同物候期。粗柄獨尾草、黑鱗頂冰花、細葉鴉蔥和小山蒜均采自古爾班通古特沙漠南部地區,伊犁郁金香采自烏魯木齊雅瑪里克山(海拔960 m),棱葉韭采自石河子將軍山(海拔580 m)。3個采集地點均屬于準噶爾荒漠,其中古爾班通古特沙漠位于準噶爾盆地中央,土壤類型為風沙土;烏魯木齊雅瑪里克山和石河子將軍山均屬于準噶爾盆地南緣低山丘陵荒漠地貌,土壤類型為棕鈣土。
野外調查時,設置2個粗柄獨尾草樣地,大小為15 m×15 m;4個小山蒜樣地,大小為5 m×5 m;其余物種均設置2個樣地,大小為10 m×10 m。利用全株挖掘法獲取樣地內全部供試植物生物量。在獲取地下生物量前,將每株植物地上部分齊地面剪除待用。植物根系挖掘的廣度和深度因種而異,以植株為圓心,粗柄獨尾草挖掘半徑為0.6~0.8 m,深度約為0.7 m;黑鱗頂冰花為0.1和0.2 m;細葉鴉蔥為0.3和0.6 m;小山蒜為0.1和0.2 m;伊犁郁金香為0.2和0.4 m。粗壯主根直接收獲,斷裂的細根利用1~2 mm土篩篩選。將每株植物的地上和地下兩部分分裝在做好標記的信封內,帶回實驗室后置于75℃烘箱內烘48 h,以保證樣品完全烘干。將每株植物的地下根系、地上葉片及繁殖器官單獨稱重、記錄。所用天平精度為0.0001 g。
1.3 生物量及其分配比例計算
百合科植物屬于單子葉植物,沒有明顯的莖,因此,將5種百合科類短命植物個體統一分為根、葉和繁殖器官(包括花和花序軸) 3個部分。分別計算各組分(地上生物量AGB、地下生物量BGB、總生物量TB和葉片生物量LB)的生物量大小,并計算根冠比(root to shoot mass ratio, R/S)、葉生物量比(leaf to total plant mass ratio, L/T)和繁殖分配(reproduction allocation ratio, RAR)。各生物量指標均采用平均值±標準誤(mean±SE)的形式給出。
1.4 數據統計分析
對6個物種的AGB、LB、繁殖器官生物量(RB)、BGB、TB、R/S、L/T及RAR進行One-way ANOVA分析。利用Levene’s test檢驗方差齊性與否,方差齊性時使用Tukey’s HSD test進行多重比較,方差不齊時則使用T2 Tamhane’s test進行多重比較[21]。R/S、RAR隨個體大小(地上生物量)的變化趨勢通過冪函數來判斷。常規數據分析和作圖在Excel 2003中完成,One-way ANOVA分析在SPSS 19.0中實現。
1.5 異速生長分析
異速生長關系是生物量分配和器官功能關系研究的重要手段和指標[17-18,22-23]。異速生長關系可以表示為Y=β·Xα,其中,Y是某種生物學特征或者功能,β是標準化常數,X指個體大小,α是異速生長指數。α=1為等速關系,即因變量和自變量呈均勻或等比例生長;α≠1為異速關系,即因變量和自變量間為不均勻或不等比例生長。確定異速生長參數時,須將冪函數轉換為logY=logβ+α·logX的形式,利用降主軸回歸(reduced major axis regression, RMA, 即Model Type II)方法計算回歸模型的指數、95%置信區間(95% CI)及決定系數(R2)[17,22]。普通最小二乘回歸(ordinary least squares regression, OLS, 即Model Type I)多用于預測2個變量之間的變化趨勢,當需要了解2個變量之間的功能關系時,RMA回歸是最合適的方法,而且該方法也是異速生長分析的標準方法[12-17]。對數化之后的α即為冪函數對數化之后線性回歸的斜率,logβ為線性回歸的截距。
不同物種間(不同群體間)的異速生長關系主要有圖1所示的4種情形[24-25]。一般地,除計算和比較異速生長指數α(即同斜率檢驗,Shift A檢驗)和截距(logβ)外,還需要對α進行等速生長檢驗(即與1.0進行比較)。對若干物種間的α進行多重比較時,若α相同(α不同即出現了Shift A),則需要進一步比較它們的截距是否相同(即截距漂移檢驗,Shift B檢驗),同時對α相同的這些物種間進行共軸漂移檢驗(Shift C檢驗),即判斷它們是否具有同一的線性擬合軸。此時,如果截距不同但共軸,則屬于Shift B情形;如果截距相同但發生了共軸漂移,則就屬于Shift C的情形;如果截距不同軸也不同,則屬于Shift D的情形;如果截距相同也同軸,那就表明這些物種的這對指標間具有完全相同的相關生長關系[24-25]。
上述異速生長指數(α)和截距(logβ)的計算、等速生長檢驗、截距漂移檢驗、共軸漂移檢驗及α的方差分析均使用SMARTR軟件完成[26]。

圖1 不同群體間異速生長關系幾種可能的變化情形[24-25]Fig.1 Schematic illustrating possible differences of allometric relationships among different plant groups[24-25]
2 結果與分析
2.1 生物量及其分配特點
準噶爾荒漠6種類短命植物單株生物量差異明顯(表1)。AGB、BGB、LB、RB和TB均以粗柄獨尾草為最大,黑鱗頂冰花最小。以TB為例,粗柄獨尾草(21.144 g/株)是黑鱗頂冰花(0.094 g/株)的近225倍。TB大于1 g/株的有3種(分別是粗柄獨尾草、細葉鴉蔥和棱葉韭),另外3種在0.094~0.702 g/株之間。小山蒜和伊犁郁金香的TB最為接近(分別為0.628和0.702 g/株),細葉鴉蔥和棱葉韭之間也無顯著差異(分別為1.290和1.855 g/株),四者的生物量均介于粗柄獨尾草和黑鱗頂冰花之間。
細葉鴉蔥具有最大的R/S (3.066±0.223),其次是粗柄獨尾草(1.441±0.077),黑鱗頂冰花(0.842±0.035)和小山蒜(0.873±0.028)的R/S基本相等,棱葉韭(0.355±0.019)的R/S最小(圖2)。6種類短命植物的L/T以黑鱗頂冰花為最大(0.289±0.006),其次是粗柄獨尾草(0.166±0.008)、細葉鴉蔥(0.151±0.007)和伊犁郁金香(0.171±0.006),最小的是同為蔥屬的小山蒜和棱葉韭,僅為(0.105±0.004)和(0.092±0.004)。繁殖分配比例(RAR)最大的是棱葉韭(0.649±0.011),其次為伊犁郁金香(0.496±0.012)和小山蒜(0.433±0.008),最小的是細葉鴉蔥(0.111±0.013),而粗柄獨尾草(0.251±0.011)和黑鱗頂冰花(0.261±0.007)的RAR幾近相等。從繁殖器官生物量與葉片生物量之比(RB/LB)也可以看出,棱葉韭(7.396±0.398)、小山蒜(4.394±0.242)和伊犁郁金香(3.046±0.161)仍較大,細葉鴉蔥(0.766±0.105)仍為最小。

表1 6種類短命植物地上生物量、葉片生物量、繁殖器官生物量、地下生物量及全株生物量Table 1 The aboveground biomass (AGB), leaf biomass (LB), reproduction organ biomass (RB), belowground biomass (BGB) and total plant biomass (TB) of six ephemeroid species
同列不同字母表示差異顯著(P<0.05)。Different letters in the same row indicate significant difference (P<0.05).
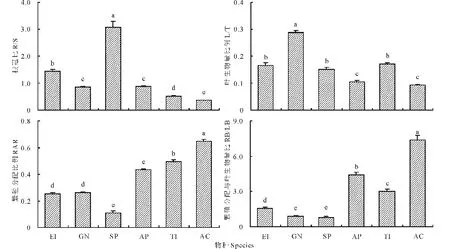
圖2 6種類短命植物根冠比、葉生物量比、繁殖分配比例及繁殖生物量與葉片生物量之比Fig.2 The root to shoot mass ratio (R/S), leaf to total plant mass ratio (L/T), reproduction allocation ratio (RAR) and reproduction organ biomass to leaf biomass ratio (RB/LB) of six ephemeroid species 物種EI、GN、SP、AP、TI、AC分別代表粗柄獨尾草、黑鱗頂冰花、細葉鴉蔥、小山蒜、伊犁郁金香和棱葉韭,不同字母表示差異顯著(P<0.05)。The species EI, GN, SP, AP, TI and AC mean E. inderiensis, G. nigra, S. pusilla, A. pallasii, T. iliensis and A. caeruleum; different letters indicate significant difference (P<0.05). 下同 The same below.
總體上,6種類短命植物用于有性繁殖和光合作用的生物量分配比例是不一致的,其中棱葉韭、小山蒜和伊犁郁金香的繁殖分配比例最高(即葉片生物量分配比例低),細葉鴉蔥最低。因此,盡管同為類短命植物,不同科、不同屬及同屬不同種之間的生物量大小及器官分配比例仍差異明顯,沒有一致的規律可循。
2.2 生物量分配與個體大小的關系
隨著植株個體大小的增加,6個物種R/S均呈現下降趨勢(圖3),除伊犁郁金香外,其他5個物種的R/S下降趨勢均達到了顯著(P<0.05)或極顯著水平(P<0.01),其中以細葉鴉蔥最為明顯(冪指數為-0.496,R2=0.6498)。這種下降趨勢在個體較小時最為強烈,當個體較大時,R/S的下降速度趨緩。因此,即使是同一種植物,由于個體大小的不同,同化產物分配到地上和地下的絕對量也是不同的;但同一種內,體型較大的植物具有相對穩定的地上和地下生物量分配比例。
RAR也隨著植株個體大小的增大發生變化(圖4),但變化趨勢基本與R/S相反。粗柄獨尾草、黑鱗頂冰花、細葉鴉蔥和小山蒜4種植物的RAR隨植株個體的增大均呈顯著(P<0.05)或極顯著(P<0.01)增加趨勢,其中以細葉鴉蔥的增加趨勢最為明顯(冪指數為0.8286,R2=0.7834),表明四者的繁殖分配與個體大小之間存在顯著的異速生長關系(冪指數<1)。伊犁郁金香和棱葉韭的RAR隨個體的增大沒有顯著變化(P>0.05),說明二者用于有性繁殖的生物量分配比例是基本恒定的(冪指數趨于0),與個體大小無關。
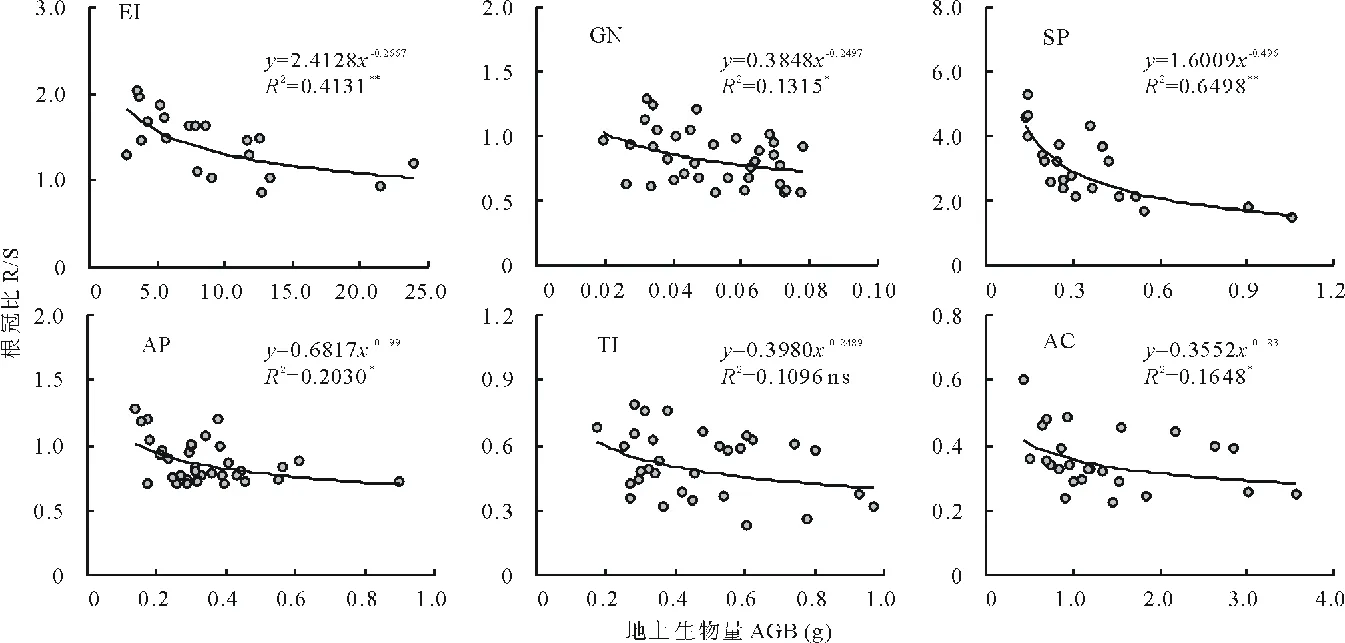
圖3 6種類短命植物根冠比隨個體大小(地上生物量)的變化趨勢Fig.3 The change trends of root to shoot mass ratio (R/S) along with plant size (aboveground biomass, AGB) of six ephemeroid species*: P<0.05, **: P<0.01, ns: P>0.05. 下同 The same below.
2.3 構件生物量間的相關生長關系
6個物種AGB-BGB間、AGB-LB間以及LB-BGB間均具有極顯著(P<0.01)的相關生長關系(圖5,表2)。等速生長檢驗表明,粗柄獨尾草和細葉鴉蔥的AGB-BGB間、細葉鴉蔥的AGB-LB間,以及小山蒜和棱葉韭的LB-BGB間的異速生長指數(α)顯著<1.0,也即屬于因變量分配速率小于自變量的異速生長關系。盡管如此,6種植物AGB-BGB間及LB-BGB間的α差異并不顯著,且各具有一個共同的α:AGB-BGB間共同的α為0.8764,LB-BGB間共同的α為0.8585,二者均顯著小于1.0,也屬于異速生長關系(表2)。6種植物AGB-LB之間的α差異明顯,且沒有共同的α;其α最大的是小山蒜、伊犁郁金香和棱葉韭,其次是粗柄獨尾草和黑鱗頂冰花,最小的是細葉鴉蔥(僅為0.6700),而且也僅有細葉鴉蔥是異速生長關系。
總體來看,不同科、不同屬以及同屬不同種之間,器官生物量AGB-BGB間以及LB-BGB間都具有相同的α,與物種種類無關。而對AGB-LB而言,菊科的細葉鴉蔥與百合科的5個物種之間的α均差異顯著,表明AGB-LB間的異速生長關系在科間是不同的;同屬的小山蒜和棱葉韭的α無顯著差異,即AGB-LB間的異速生長關系(僅指α)在屬內是相同的;而不同屬之間AGB-LB間的相關生長關系既有相同的類群(組1內相同,組2內相同;組1:小山蒜、伊犁郁金香和棱葉韭,組2:粗柄獨尾草和黑鱗頂冰花),也有不同的類群(組1和組2之間不同)。因此,6種植物器官生物量間的相關生長關系(僅指α)也不盡一致,但大部分表現出等速生長關系和與系統分類無關的特點。
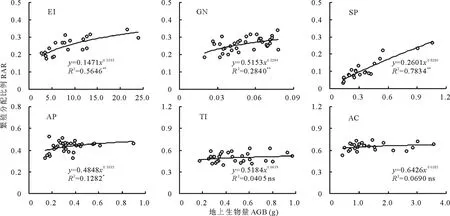
圖4 6種類短命植物繁殖分配比例隨個體大小(地上生物量)的變化趨勢Fig.4 The change trends of reproduction allocation ratio (RAR) along with plant size (aboveground biomass, AGB) of six ephemeroid species
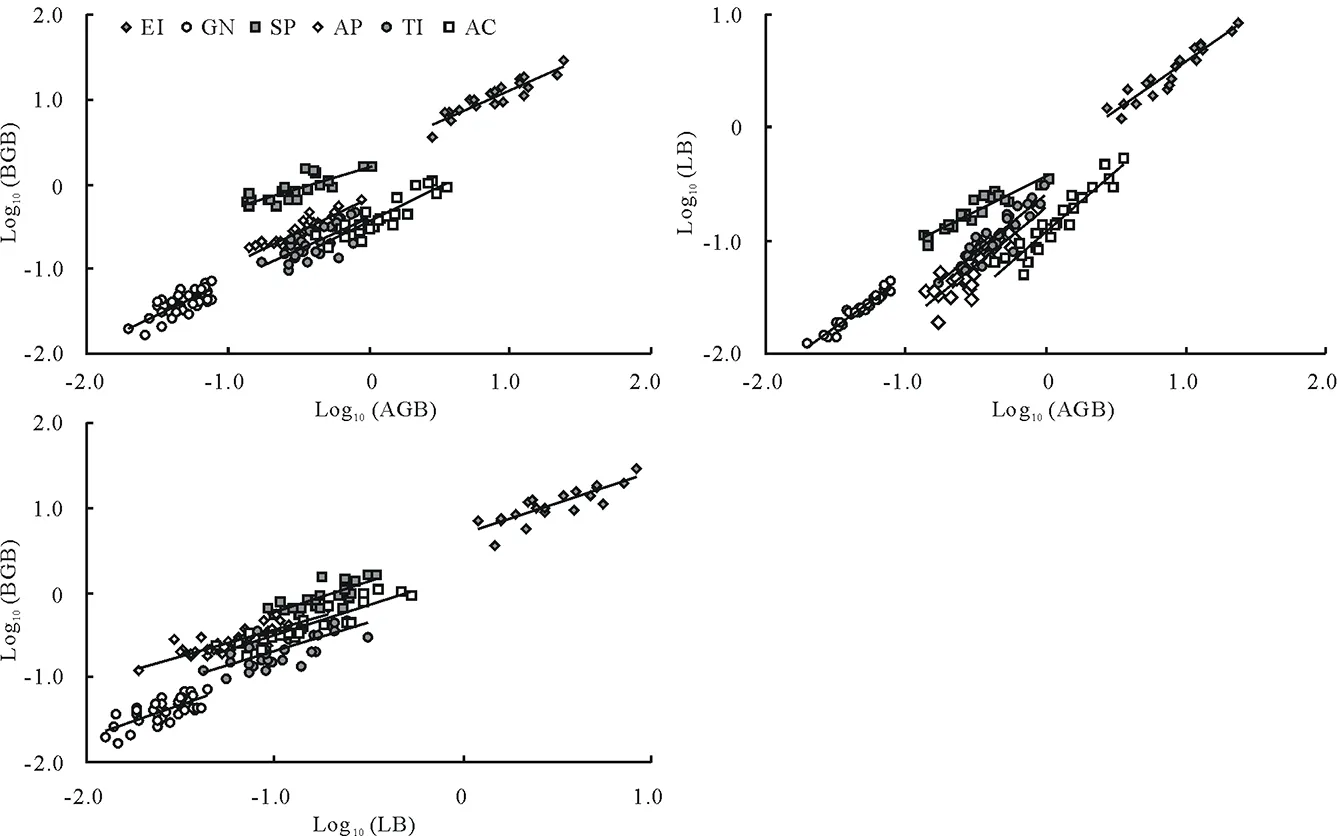
圖5 雙對數(以10為底)坐標下6種類短命植物地上(AGB)與地下(BGB)、地上與葉片(LB)及葉片與地下生物量間的RMA線性擬合關系Fig.5 Log10-Log10 linear regression (RMA) plots of aboveground biomass (AGB) and belowground biomass (BGB), AGB and leaf biomass (LB), and LB and BGB of six ephemeroid species
由于6個物種AGB-BGB間,以及LB-BGB間各自具有共同α(即未發生Shift A) (表2),故而進一步檢測它們是否具有共同的截距(Shift B檢驗)和線性擬合軸(Shift C檢驗)。結果表明(表3),AGB-BGB間粗柄獨尾草和細葉鴉蔥、黑鱗頂冰花和伊犁郁金香的截距沒有差異,但發生了共軸漂移(Shift C);LB-BGB間的粗柄獨尾草和細葉鴉蔥、粗柄獨尾草和小山蒜則發生了截距漂移(Shift B);其他所有種對間均同時發生了截距漂移和共軸漂移(Shift D)。
相關生長指數(α)一列的不同字母表示6個物種間α差異顯著(P<0.05)。 Different letters indicate significant difference (P<0.05) of six allometric scaling exponents (α).
表3 6種類短命植物3對生物量指標共45個種對異速生長關系類型Table 3 Allometric relationships of 45 species pairs related with three biomass pairs of six ephemeroid species
物種代號1~6分別是EI、GN、SP、AP、TI、AC,同圖2。The species number 1-6 are EI, GN, SP, AP, TI and AC, the same with Fig.2.
6個物種ABG-LB間沒有共同α,有11個種對的α差異顯著(發生Shift A) (表3),但有2組 (組1:小山蒜、伊犁郁金香和棱葉韭,組2:粗柄獨尾草和黑鱗頂冰花)共4對植物的α間沒有顯著差異(表2)。對這4個種對進行進一步的異速生長檢驗表明,粗柄獨尾草和黑鱗頂冰花具有相同的截距但發生了共軸漂移(Shift C),另外3對既發生了截距漂移又出現了擬合軸的漂移(Shift D)。因此,LB-ABG間并沒有Shift B的情形出現。
3 討論
3.1 不同生活型植物生物量分配特征
長期生活在相同環境下的不同物種,往往會形成相似的生物學特征,這就是趨同適應[27]。類短命植物便是由不同物種組成的具有相似生物學特性的一類功能群(functional group),它表現為地上短生、地下多年生的特點,是草本植物對干旱荒漠氣候和不均勻的季節性降水的適應。植物生物量分配是指植物將有限資源分配到不同器官及結構功能上,從而實現對外界環境的更好適應[28]。對干旱區的草本植物而言,高的根生物量分配比例也不是荒漠化演替中植被的普遍特征,其比值的高低變化和生活型或溫度的關系也許更強于和干旱程度的關系[29]。
研究表明,一次結實的草本植物繁殖分配一般占其凈同化能量的20%~40%,多次結實植物在20%以下[30]。對準噶爾荒漠幾種胡盧巴屬短命植物生物量分配的研究表明,其繁殖分配一般在50%以上,其中網脈胡盧巴(Trigonellacancellata)可達62.3%[10]。大部分短命植物的根冠比在0.1以下,地下生物量分配比例很小[9]。對科爾沁沙地70種草本植物生物量分配研究表明,無論一年生、二年生還是多年生草本,絕大部分的物種繁殖分配比例<20%、葉片生物量比例<30%,而大部分多年生草本植物表現出更大的根冠比[8]。可見,不同生活型草本植物間的生物量分配有較大差異。
艾沙江等[11]對天山北麓特有類短命植物新疆郁金香(Tulipasinkiangensis)生物量分配的研究表明,在果實成熟期新疆郁金香地下鱗莖生物量占全株的50%左右(也即根冠比約為1),繁殖分配比例可達35%以上,而葉片生物量比例不到15%。可見,新疆郁金香具有較高的有性繁殖分配比例和根系生物量分配比例,前者為土壤提供大量的種子庫來源,保證了該物種有性繁殖的繼續,而后者則為植株存儲足夠的有機物質以保證來年植株的再次萌發。
本研究表明,6種類短命植物根冠比在0.355~3.066之間,葉片生物量比例在9.2%~28.9%之間,繁殖分配比例在11.1%~64.9%之間。可見,類短命植物根的生物量在全株生物量中占有很大的比例。不同物種間的有性繁殖和光合作用的生物量分配也有明顯差異。菊科的細葉鴉蔥與百合科的粗柄獨尾草和伊犁郁金香具有相似的葉片生物量比例,且同為蔥屬的小山蒜和棱葉韭的葉片生物量比例也相近(但最小)。棱葉韭、伊犁郁金香和小山蒜的繁殖器官生物量比例較大,細葉鴉蔥最小。因此,這種生物量分配比例的差異表明不同科、不同屬及同屬不同種之間的生物量大小及器官分配比例沒有一致的規律可循。盡管如此,與荒漠一年生草本[8]和短命植物[7]相比,類短命植物地下生物量比例較大,體現了類短命植物地上短生、地下多年生的特點。
同一物種不同生境間的生物量分配一般會發生明顯改變,主要歸因于植物對環境脅迫的適應[25,31],體現了植物的可塑性(plasticity)。而不同物種間生物量分配的異同可能主要體現了系統分類地位或遺傳發育的影響,如本研究發現同為蔥屬的小山蒜和棱葉韭具有相同的葉片生物量比例和相近的繁殖器官生物量比例,而屬間及科間不同物種生物量分配的相似性很低。因而,屬一級分類單元內的物種間可能具有較相似的生物量分配特征,而屬間及科間則會有較大差異。
3.2 植物生物量分配與個體大小的關系
個體大小顯著影響植物生物量向營養器官和繁殖器官的分配比例,二者之間存在“此消彼長”的權衡關系[11]。換句話說,生物量分配存在顯著的個體發育漂變(ontogenetic drift)[25]。本研究中,6種短命植物中有5種植物的根冠比隨個體的增大而顯著減小,表明個體較大的植株其根系生物量分配比例下降而地上部分生物量分配比例增加。增加的地上生物量分配既可能用于營養器官,也有可能用于繁殖器官,這主要取決于外界環境和資源狀況[32]。當外界環境較優越時,植物會更多地將同化產物分配給營養器官,而偏輕于繁殖器官;反之,當外界環境較為惡劣時,植物會增大繁殖投入,以確保產出足夠的種子。
綜合以往研究,植物花期繁殖體重量確實隨植株個體的增大而顯著增加,但是繁殖分配與個體大小間則沒有表現出固定的模式。5種毛茛科(Ranunculaceae)植物[33]、3種風毛菊屬(Saussurea)植物[34]、黃帚橐吾(Ligulariavirgaurea)[35]等均表現為個體越大繁殖分配越低,即個體大小與繁殖分配存在異速關系,說明這些物種的繁殖分配受個體大小制約。小半灌木油蒿(Artemisiaordosica)的繁殖分配則呈現出在一定范圍內隨個體增大而上升,然后又隨個體繼續增大而下降的趨勢[36]。青藏高原東緣高寒草甸5種一年生龍膽屬(Gentiana)植物繁殖分配與個體大小間無固定的相關模式,體現了不同物種對相似環境壓力的適應對策不同[37]。垂穗披堿草(Elymusnutans)的繁殖分配比例與個體大小間沒有明顯的相關性,其繁殖分配比例表現為一個相對固定的值[38]。類似地,4 種胡盧巴屬(Trigonella)短命植物的繁殖器官生物量也不隨個體大小而發生變化,表明其生殖生物量是不具大小依賴性的,營養生長與生殖生長的增加是成比率的[10]。
本文對6種類短命植物的研究表明,有4種植物(粗柄獨尾草、黑鱗頂冰花、細葉鴉蔥和小山蒜)的繁殖分配隨個體的增加而顯著增加,但屬于明顯的異速生長關系(冪指數<1),也即繁殖分配與個體大小的生長是不等比例的;另外2個物種(伊犁郁金香和棱葉韭)的繁殖分配沒有顯著變化。這表明不同種類的類短命植物器官生物量分配與個體大小間無一致的變化規律,但多表現為顯著的異速增加的趨勢。由此可見,本研究的結果與毛茛科植物[33]、風毛菊屬植物[34]及黃帚橐吾[35]等生境相對優越的植物的研究結果相反,也不同于同地區的短命植物的結果[10],這是否是類短命植物的一個共同特點?
3.3 類短命植物構件生物量間的相關生長關系
植物體生物學特征間的相關生長關系已通過分形分配網絡模型、代謝理論等多種方法得到有力證明,并得到大量實測數據的驗證[12-18,23]。相關生長關系與尺度無關,其通過對數化除去了個體大小(尺度)對生物量分配規律的影響,揭示了不同物種間生物量分配的基本速率問題。相關生長關系理論認為在全球尺度范圍內,自然條件下種內和種間個體水平生物量具有相同的分配速率——由一個世界范圍的數據庫(包括生長于不同生境的草本與樹木,包括雙子葉植物、單子葉植物和裸子植物)得到[12-16]。相關指數始終為1.0體現了恒定的資源分配速率,而變化的相關常數logβ則反映了不同物種器官生物量絕對量的差異,在自然條件下這是由物種自身屬性所決定的。
因此,僅從異速生長指數上看,6種植物器官生物量間大部分表現為等速生長關系,這符合理論預測[12-16];而且AGB-BGB間及LB-BGB還各自具有共同的異速生長指數,這是物種間趨同適應的表現;但6個物種間沒有出現完全一致的相關生長關系,這可能是由系統分類地位決定的遺傳特性的差異導致的。
致謝:中國科學院新疆生態與地理研究所博士研究生周曉兵、程軍回、吳林參與了野外取樣,新疆農業大學碩士研究生張樂、王韻、齊會楠幫助測定部分植物的生物量,邱東幫助測定、錄入和校對數據,在此一并感謝。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
鐵道通信信號(2020年9期)2020-02-06 09:15:22
數學大王·趣味邏輯(2019年5期)2019-06-13 20:27:43
小學科學(學生版)(2019年5期)2019-05-21 01:00:18
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
經濟技術協作信息(2018年30期)2018-11-22 06:20:24
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
