角果藜的結實格局及地下散布單位的入土機制
2014-11-07 01:46:48王喜勇高蕊周曉青魏巖
草業學報 2014年2期
關鍵詞:植物
王喜勇,高蕊,周曉青,魏巖
(1.新疆農業大學草業與環境科學學院 新疆草地資源與生態重點實驗室,新疆 烏魯木齊 830052; 2.中國科學院新疆生態與地理研究所,新疆 烏魯木齊 830011;3.遼寧丹東市青山保護局,遼寧 丹東 118000; 4.陜西寶雞石油中學生物組,陜西 寶雞 721001)
植物物候是植物為了適應氣候條件的節律性變化而形成的與此相應的植物發育節律[1],是植物在進化過程中遺傳下來的為生存不斷適應環境的結果[2]。植物在不同物候期的物候特點,集中體現了植物體在不同環境條件下所采用的適應策略。
在異質性環境中,種子多型性被認為是對時空異質環境的一種適應[3]。地上地下結果性(amphicarpy) 是種子多型性中的特殊一種,是指同一植株產生不同形態的地上果實(aerial fruit)和地下果實(subterranean fruit)的現象[4-5]。其中,地下結果性又分為兩類,即植物體地下開花地下結果和基部開花地下結果[6]。關于基部開花地下結果植物地下果實形成機制的研究已較為深入,Barker[6]提出了主動地下結果性和被動地下結果性。主動地下結果性指植物體通過一定的生長方式或向性方式形成的小梗或小柄的主動方式使果實入土;而被動地下結果性指植物體借助風或水的作用將覆蓋植物體或果實的土壤沉積下來的被動方式使果實入土。地上地下結果性現象作為植物對生態環境的一種獨特適應方式,在研究植物的生態適應機制、生活史對策進化方面具有重要價值。
角果藜(Ceratocarpusarenarius)隸屬藜科角果藜屬,為一年生草本植物,廣泛分布于亞洲中部的荒漠和沙漠中[7]。在我國僅生長在準噶爾盆地,并能在準噶爾盆地的局部地區形成層片或群聚[8]。角果藜是首次在藜科中發現的具有地上地下結果性的物種[9]。那么,角果藜的地下散布單位是如何入土的?其入土的時空特點如何?本文通過對角果藜的物候、地上地下結實格局及地下散布單位入土方式的研究,揭示地下散布單位的入土機制,探討角果藜適應荒漠環境的生活史特征,對于深入了解荒漠植物的進化規律及其對特定環境的生態適應機制具有重要的意義。
1 材料與方法
1.1 研究材料與研究地概況
角果藜為藜科角果藜屬一年生草本植物,3月初萌發,花期4-8月,花單性,雌雄同株,具有地上地下結果特性。角果藜的果實為胞果,由雌花發育形成。雌花無花被具2枚苞片,受精后,2苞片完全愈合將果實包被。本文將果實及包被其外面的苞片稱為散布單位(dispersal units)。地上散布單位倒三角形,地下散布單位倒卵形[9]。
研究區位于新疆準噶爾盆地南緣低山礫質荒漠的自然種群(43°48′ N, 87°40′ E, 750~850 m),土壤為細礫質灰棕荒漠土,植被類型為小半灌木荒漠小蓬群系,天然植被以小蓬(Nanophytonerinaceum)、散枝豬毛菜(Salsolabrachiata)、紫翅豬毛菜(Salsolaaffinis)、角果藜為主,蓋度20%~30%。該地區屬中溫帶荒漠區,春秋季較濕潤,夏季干旱,熱量充足,冬季漫長而寒冷。≥5℃初日溫度出現在3月下旬,≥10℃初日溫度出現在4月下旬;高溫出現在7月,平均最高氣溫為29.7℃[10]。
1.2 研究方法
1.2.1物候觀測 2007—2008連續2年對角果藜自然種群的物候進行了觀察。物候參照宛敏渭與劉秀珍[11]的物候觀測法,在萌動期至開花期每隔2 d觀察1次,果實形成期至成熟期每隔7 d觀察1次。
1.2.2分支方式與結實格局的觀察 在果熟期觀察植株的分支方式、散布單位在植株上的空間位置,并用AutoCAD軟件繪制角果藜的分枝方式和散布單位空間分布圖。
1.2.3地下果實入土方式的測定 于2007年4月下旬-10月下旬每隔30 d隨機采集20株角果藜,測量地下散布單位頂端及其著生基部離地距離(20個重復,取平均值)。同時,在Motic B5型生物顯微鏡下測量地下散布單位與其主莖間的角度(20個重復,取平均值),采用SPSS 11.0軟件對夾角與生殖生長天數間進行相關性分析,確定地下散布單位的入土方式。
2 結果與分析
2.1 物候特征
角果藜的種子萌發早,于3月中上旬冰雪未完全融化時就開始萌發;3月中上旬子葉出現到4月中上旬枯落,子葉期持續時間長(2007年29 d;2008年28 d);完全營養生長期較短,從萌發到現蕾(2007年36 d,2008年29 d),僅占整個生活周期的約1/5,之后,營養生長和生殖生長并存;生殖時間早,萌發后約30 d現蕾,生殖期持續時間相對較長(2007年179 d,2008年173 d),約占整個生活周期的4/5以上(表1,圖1)。

圖1 角果藜的物候圖(2007,2008年)Fig.1 The phenological chart of C. arenarius in 2007 and 2008
角果藜開花后,苞片迅速愈合并增大形成倒三角形或倒卵形的散布結構,而果實則“停滯”發育,至8月下旬果實開始膨大,9月末以“集中、快速”的模式發育成熟。
角果藜2007,2008年的物候見表1。
2.2 角果藜的分枝方式及結實格局
觀察發現,角果藜為三歧聚傘狀分枝方式,枝頂形成聚頭狀雄花序后,停止發育,其下部葉腋依次形成3個分枝,植物體共有5~6級分枝形成球狀。同時,主莖第1輪雄蕊著生的節膨大并形成離層。角果藜地上散布單位為倒三角形,多數,單生于葉腋,呈螺旋上升狀排列于每個分枝上。地下散布單位為倒卵形,2個,位于主莖第1節處,僅為第1對真葉葉腋處的1對雌花受粉后發育形成(圖2)。

表1 角果藜的物候特點(2007,2008年)Table 1 Phenological characteristics of C. arenarius in 2007 and 2008
a,萌動 Germination;b,展葉 Seedling;c,子葉敗落 Cotyledon decline;d,現蕾 Flower bud appearing;e, 始花 First flower appearing; f,盛花 Full blooming;g,末花 Flowering termination;h,地下果形成Underground fruit appearing;i,地上果形成Aboveground fruit appearing;j,果熟 Fruit maturing;k,生活周期Life cycle.

圖2 角果藜結實格局Fig.2 Branching mode and fruit-set patterns of C. arenarius
2.3 地下散布單位的入土方式
地下散布單位是由第1節對生葉腋處的2朵雌花發育而來。4月下旬,2朵雌花地上開花授粉,雌花外2枚苞片于5月中上旬完全愈合形成散布單位,散布單位于6月中下旬入土繼續發育形成地下散布單位。從圖3和表2可以看出,地下散布單位與角果藜主莖間的夾角隨時間變化逐漸增大,由初形成時的(32.05±0.32)°(圖3-1)達到生殖生長135 d后(134.01±0.60)°(圖3-3)的最大值。相關性分析顯示,地下散布單位與主莖間夾角與生殖生長天數之間呈顯著正相關關系(Person相關系數為0.920,P<0.01);表3顯示,隨地下散布單位的翻轉與增大,其頂端離地表距離逐漸減小,在6月末、7月初地下散布單位頂端進入土壤。同時,在風沙及雨水帶來的土表覆蓋物掩埋作用下,地下散布單位完全進入土壤。因此,地下散布單位在主動性的“翻轉”和被動性埋藏的雙重作用下,順利完成入土。

圖3 角果藜地下散布單位翻轉過程Fig.3 Reversing process of subterranean dispersal units of C. arenarius

項目Item生殖生長時間 Time of procreation growth 15 d (4月 April)30 d (5月 May)45 d (5月 May)60 d (6月 June)75 d (6月 June)90 d (7月 July)夾角 Angle (°)32.05±0.3254.62±1.04106.24±2.02133.17±0.45134.01±0.60133.68±0.88
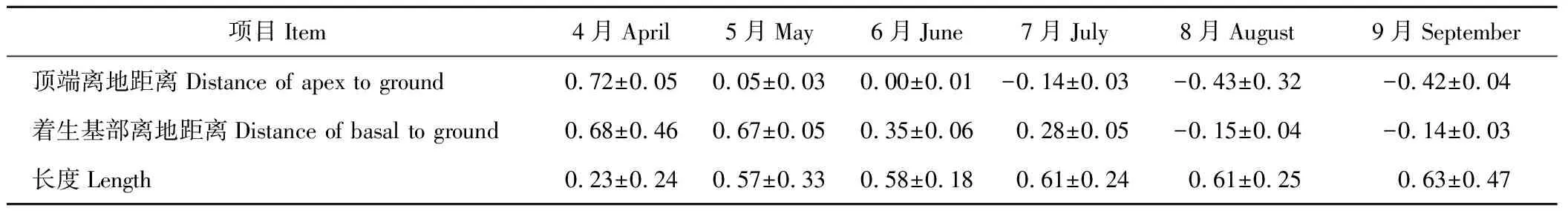
表3 地下散布單位著生基部、頂端離地距離及其長度Table 3 The distance of apex and basal to ground and length of subterranean dispersal units for C. arenarius (mean±SE) cm
3 討論
不同的物候特征對植物的生殖成功有重要影響[12-14]。在新疆準噶爾盆地3-4月的早春,氣溫逐漸升高,≥5℃初日溫度出現在3月下旬,≥10℃初日溫度出現在4月下旬,實踐表明5~6℃是植物發育的基礎溫度[15-16],同時積雪融化,淺層土壤形成充足的水分,整個生活史內早春是土壤含水量最高的時期,這些條件對植物的生長發育極為有利。角果藜利用有利的環境條件,提早萌動從而保證較高的資源利用效率。子葉是種子植物胚的組成部分之一,為貯藏養料或幼苗時期進行同化作用的器官,所以子葉在種子萌發成幼苗的初期,其作用十分重要。角果藜的子葉持續長約30 d,不但將其儲藏的養料最大程度的提供給幼苗,同時綠色子葉的光合產物也是幼苗早期主要的能量來源,保證幼苗的成功生長。角果藜完全營養生長時間短,為有性生殖積累基本能量;生殖時間早,在幼態下進行生殖,營養生長和生殖生長同時進行,持續時間較長,避免了夏季干旱高溫來臨,植物尚未結實而導致種群滅絕。繁殖時間是由遺傳決定的,以適應不同的環境壓力[17-18],如有較大的死亡危險,推遲種子產生就可能導致繁殖的徹底失敗。6月夏季來臨,溫度逐漸升高,土壤含水量下降,環境條件對植物的生長發育極為不利,短命植物大多果實已成熟并完成了其生活史[19]。而非短命植物角果藜仍處于營養生長和生殖生長并存期,通過觀察發現,角果藜開花后,花外苞片愈合,但包被其內的果實處于“停滯發育”期,直至8月下旬惡劣環境逐漸好轉時果實才以“集中、快速”的模式發育成熟。這種繁殖策略,既能使植物體實現最大的繁殖產量,又能使其逃避準噶爾荒漠夏季干熱氣候對其種子繁殖所帶來得高死亡風險,最終達到生殖成功。
研究表明種子的異型性符合兩面下注(bet-hedging) 策略,主要表現在通過產生不同休眠程度的種子和不同空間擴散程度的種子,在時間和空間上分散風險,從而適應環境時空異質性[20]。在數量上,角果藜地上散布單位多數,地下散布單位僅為2個,植物通過感受環境脅迫,產生更多的容易擴散的種子,使植物的后代能夠逃離不利的環境,尋找比較適宜的環境[21];在休眠程度上,地上散布單位比地下散布單位的休眠程度淺,易于萌發,而休眠程度較深的地下散布單位則可以形成持久地下種子庫;在擴散方式上,地上散布單位隨母體“風滾”散布拓展后代分布范圍,地下散布單位保留在母體附近不易擴散[9];在形成機制上,角果藜形成散布單位的花具有專一性,即只有第1對真葉葉腋處的1對雌花受粉后發育成地下果實,而其他雌花(兩種雌花無形態差異)受粉后均形成地上果實,這種專一性使植物體開花后首先確保地下散布單位的形成;在散布單位的分布格局上,地下散布單位近地表形成利于入土并被沙土埋藏,并且保留在母體附近進行原位萌發,對地下果實的投資為種群在原分布區的繁衍提供了可能,使其能夠在惡劣的荒漠環境達到最大的繁殖成功。地上散布單位著生于三歧聚傘狀分枝形成的球型植株上,利于其風滾散布時的散布效率。角果藜的地上地下兩種散布單位在數量、萌發程度、擴散方式[9]、形成機制、結實格局上的差異性都充分體現了這種兩面下注的適應策略。
在基部開花地下結果植物地下果實入土機制中,地下果實被動性的入土行為主要是借助風沙、雨水等外力因素來實現,主動性入土機制主要是通過特化結構來實現的[6]。豆科中的3個種(Trifoliumsuberraneum,Voandzeiasubterranean,Derstingiellageocarpa)是通過某些向性生長方式形成小梗或小柄的方式完成地下果實的主動入土[22]。單子葉植物(Narcissus,Eucomispallidiflora,Eucomisautumnalis)和雙子葉植物Catanancheluten(Compositae)通過收縮根使莖基部的果實入土[23-25]。Wilson 和 Honey[26]通過解剖實驗研究發現收縮根通過收縮區中內部皮層薄壁組織細胞的縱向縮短和垂直擴展,同時還有包括皮層外部細胞、外皮層和表皮在內的外部組織越過收縮區表面被推入巨大的褶皺中進行收縮。本文研究結果表明,角果藜是通過主動性和被動性的雙重作用使地下散布單位入土的。一方面,不僅可借助風沙等外力因素來埋藏地下散布單位。同時,果實主動性的動態“翻轉”模式是更為節省且有效的入土策略。作為荒漠植物,生存環境極其惡劣,而特化組織的形成需要更多的能量分配。因此角果藜選擇這種特殊的主動入土模式即保證了果實的順利入土,又節省了能量分配,是其對荒漠環境長期適應的結果。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
