Kv1.5對大鼠低氧高二氧化碳性PASMCs增殖、凋亡的影響及與MAPK通路的關系*
2014-11-08 02:28:06馬迎春鄭夢曉黃林靜王園園王萬鐵
中國病理生理雜志 2014年9期
馬迎春, 鄭夢曉, 黃林靜, 王園園, 應 磊,2, 王萬鐵,2△
研究表明,慢性缺氧可降低肺動脈平滑肌細胞(pulmonary artery smooth muscle cells,PASMCs)胞膜上電壓依賴性鉀離子通道(voltage-dependent K+channel,Kv)的活性,同時還可直接調控 Kv通道基因的變化進而引起PASMCs的收縮和增殖;除此之外,胞質中充足的鉀離子還可以抑制PASMCs的凋亡[1]。PASMCs上 Kv通道功能失調,尤其是Kv1.5的表達下調及功能障礙,已成為肺動脈高壓的基本特征之一[2-3]。絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)信號通路是各種信號通路的匯集點,對其它信號通路有重要調控作用,在應激、創(chuàng)傷、炎癥和休克等疾病的發(fā)生過程中起一定的作用。該通路的激活可能參與肺動脈高壓的發(fā)生且研究發(fā)現[4]。因此有關低氧和MAPK信號通路之間的關系也漸漸成為研究的熱點。而實際情況中低氧一般都伴隨有高二氧化碳的潴留,故在實驗中我們選擇復制低氧高二氧化碳模型。本實驗將采用CCK-8法檢測細胞活性,Western blotting檢測鉀離子通道 Kv1.5、增殖細胞核抗原(proliferating cell nuclear antigen,PCNA)及Bax蛋白的表達水平,以此來研究低氧高二氧化碳條件下 Kv1.5對離體大鼠PASMCs增殖、凋亡的影響,同時探討這些變化與MAPK通路之間的關系。
材料和方法
1 動物和主要試劑
SPF級健康雄性 Sprague-Dawley(SD)大鼠24只,體重(220±20)g,由溫州醫(yī)科大學實驗動物中心提供[SCXK(浙)2010-0044號]。
SB203580、U0126、茴香霉素(anisomycin)、Kv1.5兔抗鼠多克隆抗體、Ⅰ型膠原酶、木瓜蛋白酶和牛血清白蛋白(Sigma);胎牛血清、DMEM高糖培養(yǎng)基和胰酶/EDTA(Gibco);小鼠 α-平滑肌肌動蛋白(αsmooth muscle actin,α-SMA)單克隆抗體和辣根過氧化物酶(horseradish peroxidase,HRP)標記的羊抗兔Ⅱ抗(Abcam);FITC標記的羊抗小鼠IgGⅡ抗(武漢博士德公司);HRP標記的小鼠單克隆β-actin(上海康成公司);PCNA兔抗鼠多克隆抗體(Bioworld);Bax兔抗鼠多克隆抗體(Santa Cruz);Cell Counting Kit-8(CCK-8)試劑盒(同仁化學);二硫蘇糖醇(Merck)。
2 大鼠PASMCs的培養(yǎng)鑒定及實驗分組
常規(guī)麻醉大鼠后用75%乙醇消毒,于超凈臺內迅速取出心肺組織,在解剖顯微鏡下分離獲得3~4級肺動脈(直徑300~700 μm),輕微剝除外膜及去除內皮細胞后將平滑肌剪成1 mm×1 mm×1 mm大小的組織塊。加入消化酶于37℃消化15 min,吸出消化酶加入含血清的DMEM終止消化,玻璃滴管將其輕輕吹打成細胞懸液,靜止3 min,待未被消化的組織塊沉淀后吸出細胞懸液于另一離心管中。余組織塊繼續(xù)加入消化酶37℃消化5 min后加入培養(yǎng)液終止消化,吹打成細胞懸液,如此反復2~3次即可將組織塊完全消化成單個PASMCs。收集PASMCs種植于培養(yǎng)瓶中,加入含20%胎牛血清的高糖培養(yǎng)基,置于37℃、5%CO2培養(yǎng)箱中培養(yǎng)。2~3 d后細胞已經貼壁并長成長梭形,并呈“峰-谷”狀生長,如培養(yǎng)液呈黃色可給予換液,1周左右區(qū)域細胞已融合生長,即可傳代。細胞傳至3代后,按傳代方法消化,吹打細胞,然后使細胞在第1瓶中貼壁15 min,光鏡下可見部分細胞貼壁;后將第1瓶中培養(yǎng)基移入第2瓶中,使細胞再次貼壁15 min,光鏡下又可見大部分細胞貼壁;最后再將第2瓶中培養(yǎng)基移入第3瓶中,再使細胞貼壁,后將3瓶均補齊培養(yǎng)基。繼續(xù)培養(yǎng)1~2 d后,光鏡下觀察,第1瓶中以較大長梭形細胞為主,為成纖維細胞;第2瓶為混合細胞;第3瓶中為較純的梭形平滑肌細胞;取第3瓶細胞繼續(xù)培養(yǎng)。也可重復純化幾次;獲得較純的平滑肌細胞。體外培養(yǎng)的大鼠肺動脈平滑肌細胞用激光共聚焦免疫熒光染色法鑒定,具體方法參考本室黎關龍等[5]先前的報道。
選第3~5代生長良好的對數生長期PASMCs,將其消化制成細胞懸液,按2×108cells/L的密度接種于10 cm培養(yǎng)皿及按1×108cells/L的密度接種于96孔板(100 μL),加入含10%胎牛血清的DMEM高糖培養(yǎng)基,37℃、5%CO2培養(yǎng)箱中進行培養(yǎng),待融合成單層細胞(80% ~90%)時換成無血清培養(yǎng)基饑餓24 h使細胞周期同步化然后行分組。(1)正常(normal,N)組:5%CO2、21%O2、37 ℃;(2)低氧高二氧化碳(hypoxia+hypercapnia,HH)組:6%CO2、4%O2、37℃;(3)DMSO 對照(hypoxia+hypercapnia+DMSO,HD)組:0.05%DMSO、6%CO2、4%O2、37 ℃;(4)U0126(hypoxia+hypercapnia+U0126,HU)組:10 μmol/L U0126、6%CO2、4%O2、37℃;(5)SB203580(hypoxia+hypercapnia+SB203580,HS)組:10 μmol/L SB203580、6%CO2、4%O2、37℃;(6)茴香霉素(hypoxia+hypercapnia+anisomycin,HA)組:10 μmol/L 茴香霉素、6%CO2、4%O2、37℃。以上每組5皿細胞,96孔板每組設6個復孔,培養(yǎng)72 h。
3 方法
3.1 CCK-8法測定各組PASMCs的活性 造模結束后用PBS洗滌2遍,吸干水后每孔加入110 μL的培養(yǎng)基(100 μL DMEM+10 μL CCK-8 溶液,避光操作),用錫箔紙包好后放入培養(yǎng)箱中孵育合適時間后,用酶標儀在450 nm處測其吸光度(A)值(可選取0.5 h、1 h、2 h 和4 h 幾個時點檢測),A 值表示細胞相對活性。
3.2 Western blotting 法檢測各組 PASMCs Kv1.5、PCNA和Bax蛋白的表達 提取細胞總蛋白并用BCA法測定蛋白濃度,用10%聚丙烯酰胺凝膠電泳(SDS-PAGE)分離蛋白后轉移到PVDF膜上,5%脫脂奶粉室溫封閉1 h。取稀釋的兔抗鼠Kv1.5多克隆抗體(1∶400)、兔抗鼠 PCNA多克隆抗體(1∶1 000)、兔抗鼠 Bax多克隆抗體(1∶1 000)或內參照β-actin(1∶10 000),4℃靜置孵育過夜。次日用TBST洗膜后加HRP標記的羊抗兔Ⅱ抗(1∶40 000),室溫孵育1 h。洗膜后采用ECL化學發(fā)光法獲得目的條帶。Quantity One凝膠軟件分析系統分析Kv1.5、PCNA、Bax及 β-actin 灰度值,以目的蛋白Kv1.5、PCNA、Bax條帶灰度值和 β-actin條帶灰度值的比值代表Kv1.5、PCNA和Bax蛋白表達的相對含量。
4 統計學處理
采用SPSS 17.0軟件分析,數據用均數±標準差(mean±SD)表示。多組樣本均數比較進行方差齊性檢驗,組間比較采用單因素方差分析(One-way ANOVA),方差齊性者兩兩比較采用LSD法,方差不齊者進行Dunnet’s T3檢驗。以P<0.05為差異有統計學意義。
結 果
1 激光共聚焦免疫熒光法鑒定PASMCs
原代培養(yǎng)的PASMCs呈典型的“峰-谷”狀生長。鏡下顯示細胞呈梭形,具有長短不等的數個突起,彼此融合,邊界不清,折光性好,細胞核位于細胞中央,呈卵圓形,見圖1。培養(yǎng)至第3~5代的細胞經小鼠α-SMA單克隆抗體孵育、FITC標記的羊抗小鼠Ⅱ抗孵育、PI染色后在激光共聚焦顯微鏡下觀察,約99%呈陽性反應,即高倍鏡下可見胞漿內大量發(fā)綠色熒光、與細胞長軸平行的纖維細絲,此為α-SMA;核卵圓形居中,呈紅色,見圖2。

Figure 1.Primary culture of rat PASMCs(×100).圖1 大鼠原代培養(yǎng)肺動脈平滑肌細胞
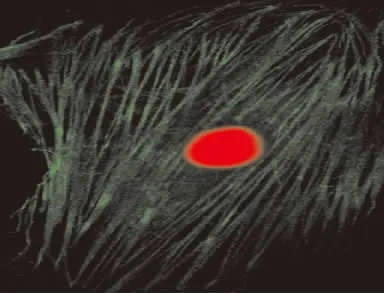
Figure 2.Immunofluorescence identification of rat PASMCs(×600).圖2 激光共聚焦免疫熒光法鑒定平滑肌細胞
2 各組PASMCs的活性變化
與N組相比,HH組、HD組細胞活性明顯增加(P<0.01),HH組和 HD組間無顯著差異(P>0.05);較之HD組,HU組、HS組及HA組細胞活性均明顯降低,且以HA組細胞活性降低最顯著(P<0.05 和 P <0.01),見圖3。

Figure 3. Changes of cell viability in all groups.N:normal group;HH:hypoxia+hypercapnia group;HD:hypoxia+hypercapnia+DMSO incubation group;HU:hypoxia+hypercapnia +U0126 incubation group;HS:hypoxia+hypercapnia+SB203580 incubation group;HA:hypoxia+hypercapnia+nisomycin.Mean±SD.n=6.** P < 0.01 vs N group;#P < 0.05,##P <0.01 vs HD group.圖3 各組細胞活性比較
3 各組PASMCs上相關蛋白的表達
3.1 各組PASMCs上Kv1.5蛋白的表達 與N組相比,HH組和HD組Kv1.5蛋白表達均明顯降低(P<0.01),HH 組和 HD 組間無差異(P >0.05);較之HD組,HU組、HS組及HA組Kv1.5蛋白表達明顯上升(P<0.01),且以HA組上升最明顯,見圖4。

Figure 4.Change of Kv1.5 protein expression in the PASMCs with different treatments.Mean ± SD.n=6.**P <0.01 vs N group;##P <0.01 vs HD group.圖4 各組Kv1.5蛋白的表達
3.2 各組PASMCs上PCNA蛋白的表達 與N組相比,HH組和HD組PCNA蛋白表達均明顯上調(P<0.01),HH組和HD組間無顯著差異(P>0.05);較之HD組,HU組、HS組及HA組PCNA蛋白表達明顯均明顯下調(P<0.01),且以HA組下降最明顯,見圖5。
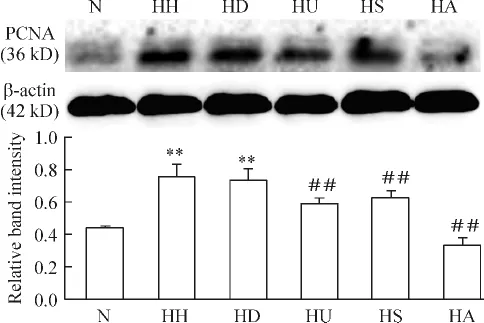
Figure 5.Change of PCNA protein expression in the PASMCs with different treatments.Mean ± SD.n=6.**P <0.01 vs N group;##P <0.01 vs HD group.圖5 各組PCNA蛋白的表達
3.3 各組PASMCs上Bax蛋白的表達 與N組相比,HH組和HD組Bax蛋白表達均明顯下調(P<0.01),HH組和HD組間無顯著差異(P>0.05);較之HD組,HU組、HS組及HA組Bax蛋白表達均明顯上調(P<0.01),且以HA組升高最明顯,見圖6。
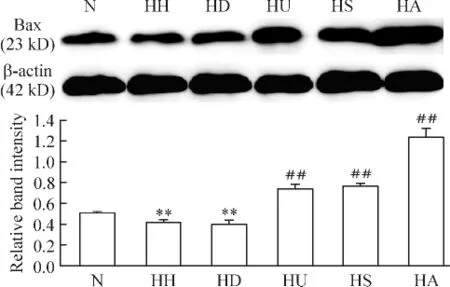
Figure 6.Change of Bax protein expression in the PASMCs with different treatments.Mean ± SD.n=6.**P < 0.01 vs N group;##P <0.01 vs HD group.圖6 各組Bax蛋白的表達
討 論
有研究表明,肺血管重構較低氧性肺血管收縮在肺動脈高壓發(fā)病機制中發(fā)揮著更為重要的作用,血管結構的重建主要表現為血管平滑肌細胞的肥大、增生和結締組織含量的變化[6]。生理情況下PASMCs存在有序的增殖與凋亡,二者保持平衡。低氧可刺激肺血管內皮細胞和平滑肌細胞等合成分泌多種細胞因子作用于PASMCs,使其增殖。PCNA是DNA多聚酶δ的輔酶,通常在細胞周期的G1末期DNA合成開始前才表達增加,在S期達高峰,是一個比較敏感的細胞增殖指標。有研究表明[7]在常氧條件下,PCNA在主動脈、肺動脈主干及肺內小動脈平滑肌細胞內均有微弱表達;但在缺氧時只有肺小動脈平滑肌細胞內表達增強,支持肺內小動脈平滑肌細胞對缺氧更為敏感的結論。另外有研究表明,Bcl-2家族的成員Bax促進細胞凋亡[8]。因此在實驗中我們分別檢測各組細胞PCNA及Bax蛋白的表達情況作為反映細胞增殖、凋亡情況的指標。
鉀離子通道在PASMCs分布廣泛且種類繁多復雜,因其平衡電位與靜息膜電位相似,而成為靜息膜電位的主要維持因素。一直以來,Kv被認為是與低氧性肺血管收縮發(fā)生機制關系最大的鉀離子通道:低氧抑制Kv,引起細胞膜的去極化增強,細胞外鈣離子內流增多,從而導致PASMCs收縮,進一步引起肺動脈的收縮,平滑肌細胞增殖,肺血管重構。研究表明,Kv主要通過2個方面來增加肺血管阻力[9-10]:(1)Kv通道表達密度改變導致細胞膜去極化,激活電壓依賴性鈣離子通道(voltage-dependent calcium channel,VDCC),Ca2+進入細胞后結合肌球蛋白輕鏈激酶使興奮收縮偶聯增強,從而引起PASMCs收縮和肺血管收縮,最終導致肺血管阻力增加,并且VDCC活性增加可抑制Kv通道并且使細胞膜處于持續(xù)去極化狀態(tài),從而引起PASMCs增殖速度的加快,進而導致肺血管肥厚;(2)Kv通道有關亞基表達的下調及全細胞鉀電流的抑制將抑制細胞凋亡,引起肺血管肥厚,從而最終引起肺動脈高壓的發(fā)生。其中Kv1.5被認為是重要的Kv通道亞型之一[11],其產生的電流是細胞膜電位去極化電流的主要成分之一,長期缺氧可導致包括Kv1.5在內的氧敏感鉀離子通道受損,肺血管長期收縮引起肺動脈高壓的發(fā)生。有報道稱,低氧明顯抑制Kv1.5通道活性和表達,隨低氧時間長短不一致,其下降程度不相同,并且常壓條件下,離體培養(yǎng)的大鼠肺動脈平滑肌細胞在低氧的環(huán)境中時間越長,Kv1.5 mRNA及蛋白表達下降越明顯[12-13],表明 Kv1.5在肺血管重構導致的肺動脈高壓中起重要作用。另外有研究表明Kv1.5的基因轉移能有效降低肺動脈高壓[2]。本研究結果也顯示,慢性低氧高二氧化碳可以明顯降低PASMCs上Kv1.5蛋白表達水平,這與已有的研究結果相一致。這進一步說明低氧高二氧化碳可能通過降低Kv1.5通道的表達引起肺動脈高壓的形成。
MAPK信號通路是一條細胞外信號引發(fā)細胞核內反應的重要途徑,在細胞形成、分化、生長、增殖及凋亡過程中發(fā)揮著重要作用[14]。目前發(fā)現MAPK家族有4條信號通路:即細胞外信號調節(jié)激酶(extracellular signal-regulated kinase,ERK)、c-Jun N 端激酶(c-Jun N-terminal kinase,JNK)、p38 絲裂原活化蛋白激 酶 (p38 mitogen-activated protein kinase,p38 MAPK)和 ERK5/BMKl通路。目前很多研究發(fā)現MAPK信號轉導通路與細胞增殖、血管重構之間的關系相當密切,并認為MAPK可能成為血管重構的治療靶點[4]。以往有研究表明[5]ERK1/2 及 p38 MAPK通路的激活參與了肺動脈高壓的形成。而目前尚未見p38 MAPK及ERK1/2對低氧高二氧化碳條件下大鼠PASMC上Kv1.5表達變化及對PASMCs增殖、凋亡影響的詳細報道。
實驗中我們在低氧高二氧化碳條件下分別用p38 MAPK抑制劑-SB203580、ERK1/2抑制劑-U0126及MAPK激動劑-茴香霉素處理 PASMCs,檢測 Kv 1.5、PCNA及Bax的表達情況,以此來探討MAPK對低氧高二氧化碳條件下大鼠PASMCs上Kv1.5表達變化的影響及與PASMCs增殖、凋亡的關系。本實驗發(fā)現,低氧高二氧化碳時PASMCs上 Kv1.5蛋白、Bax蛋白表達降低,細胞活性、PCNA蛋白表達上調,而p38 MAPK通路抑制劑-SB203580能夠明顯逆轉這種現象;ERK1/2通路特異阻斷劑-U0126與SB203580有相似的作用。這說明低氧高二氧化碳可能是通過激活p38 MAPK通路和ERK1/2通路引起PASMCs Kv1.5表達降低,從而導致鉀離子外流減少,細胞膜去極化,鈣離子內流增加,從而導致PASMCs收縮和增殖加強,凋亡延緩。另外,低氧高二氧化碳性PASMCs給予MAPK通路激活劑-茴香霉素可導致Kv1.5蛋白、Bax蛋白表達繼續(xù)升高,同時也明顯高于HU組或HS組;而細胞活性、蛋白PCNA表達繼續(xù)下降,低于HU組及HS組。推測可能是由于在此種低氧高二氧化碳性條件下茴香霉素主要激活了MAPK通路中負責促進細胞凋亡的亞族及亞基,如JNK通路或p38 MAPK中的某一亞基,具體機制還需要進一步的探索。
本研究的結果顯示SB203580和U0126對低氧高二氧化碳條件下各組細胞活性,Kv1.5、PCNA及Bax蛋白表達的影響是相似的,說明p38 MAPK通路和ERK1/2通路在低氧高二氧化碳性肺動脈高壓中的作用可能相似,這與已有的大部分研究結果是一致的。但有學者認為[15]ERKl/2通路和p38 MAPK通路之間可能存在相互制約關系,抑制p38 MAPK通路可加強ERK1/2的激活,導致凋亡延遲。也有人認為ERK1/2和p38 MAPK通路之間存在著平衡和精確的調節(jié),而這種相互作用是影響細胞增殖、分化、凋亡和死亡的重要調控機制。另有研究表明,ERK1/2通路還可拮抗JNK和p38 MAPK通路的促細胞凋亡活性,促進細胞的生存[16],并且ERK1/2通路激活可誘導MKP(mitogen-activated protein kinase phosphatase)表達增強,從而抑制p38 MAPK的過度激活[17]。另外MAPK與其它信號通路間也存在著復雜的相互調節(jié)關系,也許是通過PKC、PKA、Raf-MEK-ERK1/2等多種途徑共同影響低氧高二氧化碳性肺動脈高壓的發(fā)生發(fā)展。這是一個相當復雜精密的調控系統,其中的具體作用機制尚需進一步探究。
[1] Semenza GL.Oxygen sensing,homeostasis,and disease[J].N Engl J Med,2011,365(6):537-547.
[2] Pozeg ZI,Michelakis ED,Dyck JR,et al.In vivo gene transfer of the O2-sensitive potassium channel Kv1.5 reduces pulmonary hypertension and restores hypoxic pulmonary vasoconstriction in chronically hypoxic rats[J].Circulation,2003,107(15):2037-2044.
[3] Remillard CV,Tigno DD,Platoshyn O,et al.Function of Kv1.5 channels and genetic variations of KCNA5 in patients with idiopathic pulmonary arterial hypertension[J].Am J Physiol Cell Physiol,2007,292(5):C1837-C1853.
[4] 孔春初,戴愛國.慢性阻塞性肺疾病患者肺部絲裂原活化蛋白激酶、蛋白激酶B和缺氧誘導因子1α的表達[J].中華結核和呼吸雜志,2006,29(6):372-375.
[5] 黎關龍,黃林靜,王萬鐵,等.容量激活性氯離子通道在低氧高二氧化碳處理的大鼠PASMCs的表達及其與MAPK通路的關系[J].中國病理生理雜志,2014,30(1):25-29.
[6] Burke DL,Frid MG,Kunrath CL,et al.Sustained hypoxia promotes the development of a pulmonary artery-specific chronic inflammatory microenvironment[J].Am J Physiol Lung Cell Mol Physiol,2009,297(2):L238-L250.
[7] 羅 穎,李志超,宋祖軍,等.VEGF和PCNA在慢性低氧性肺動脈高壓大鼠主動脈、肺動脈平滑肌細胞中表達[J].細胞與分子免疫學雜志,2006,22(1):103-105.
[8] Oltvai ZN,Milliman CL,Korsmeyer SJ.Bcl-2 heterodimerizes in vivo with a conserved homolog,Bax,that accelerates programmed cell death[J].Cell,1993,74(4):609-619.
[9] Fantozzi I,Platoshyn O,Wong AH,et al.Bone morphogenetic protein-2 upregulates expression and function of voltage-gated K+channels in human pulmonary artery smooth muscle cells[J].Am J Physiol Lung Cell Mol Physiol,2006,291(5):L993-L1004.
[10] Firth AL,Gordienko DV,Yuill KH,et al.Cellular localization of mitochondria contributes to Kv channel-mediated regulation of cellular excitability in pulmonary but not mesenteric circulation[J].Am J Physiol Lung Cell Mol Physiol,2009,296(3):L347-L360.
[11] Archer SL,Souil E,Dinh-Xuan AT,et al.Molecular identification of the role of voltage-gated K+channels,Kv1.5 and Kv2.1,in hypoxic pulmonary vasoconstriction and control of resting membrane potential in rat pulmonary artery myocytes[J].J Clin Invest,1998,101(11):2319-2330.
[12]徐賢華,彭華生,肖欣榮,等.二氯醋酸鈉對模擬高原肺動脈高壓大鼠肺動脈平滑肌細胞Kv1.5亞型的作用研究[J].中國呼吸與危重監(jiān)護雜志,2010,9(1):53-56.
[13]Jian W,Letitia W,Wenqian W,et al.Chronic hypoxia inhibits Kv channel gene expression in rat distal pulmonary artery[J].Am J Physiol Lung Cell Mol Physiol,2005,288(6):L1049-L1058.
[14]Cuevas BD,Abell AN,Johnson GL.Role of mitogen-activated protein kinase kinase kinases in signal integration[J].Oncogene,2007,26(22):3159-3171.
[15] Sheth K,Friel J,Nolan B,et al.Inhibition of p38 mitogen activated protein kinase increases lipopolysaccharide induced inhibition of apoptosis in neutrophils by activating extracellular signal-regulated kinase[J].Surgery,2001,130(2):242-248.
[16]杜宏舉,湯 寧.絲裂原活化蛋白激酶信號通路及其生物學功能[J].國外醫(yī)學:衛(wèi)生學分冊,2005,32(4):197-201.
[17]黃軼峰,龔開政,張振剛.p38 MAPKα/β和ERK1/2在心肌缺氧預處理信號傳遞中的不同作用[J].生理學報,2003,55(4):454-458.
