真蛸谷胱甘肽過氧化物酶和過氧化氫酶基因克隆、序列特性分析
2014-12-01 08:16:54洪婧妮蘇永全孫田田
廈門大學學報(自然科學版) 2014年4期
關鍵詞:實驗
洪婧妮,蘇永全,毛 勇,孫田田,王 軍
(廈門大學海洋與地球學院,福建 廈門361102)
真 蛸 (Octopus vulgaris)隸 屬 于 軟 體 動 物 門(Mollusca)、頭足綱(Cephalopoda)、八腕目(Octopoda)、無須亞目(Incirrata)、蛸科(Octopodidae)、蛸屬(Octopus),具有生活史短、生長速度快、食物轉化率高、對飼養環境和人工運輸具有較強的耐受力和適應力等諸多優良的養殖生物學特性[1-2].真蛸是暖溫種,在26℃以上水溫難于生存,有關其適宜生長溫度已有所研究[3-4].
生物體在常態下保持體內活性氧的收支平衡,已有研究發現,高水溫會使變溫生物細胞吞噬活性增強,從而導致大量活性氧爆發[5-6],過量的自由基如未能及時清除,會對機體的脂質代謝、酶活性和DNA等造成嚴重損害[7].為了避免過剩的自由基對機體造成損傷,生物體內形成一套抗氧化系統,在清除過量自由基,保持機體活性氧動態平衡中起到了重要作用[8-10].
1957年Mills等在牛紅細胞中發現谷胱甘肽(GSH)過 氧 化 物 酶 (glutathione peroxidase,GPX)[11],之后發現該基因為硒蛋 白,具 有催化 基團——硒代半胱氨酸(Selenocysteine,Sec U)[12-13].至今已發現了7種GPX種類,其中細胞內GPX1(cGPX)是第一個被發現的哺乳動物硒蛋白,也是目前克隆得到較多的類型.1863年首次發現一類具有分解過氧化氫作用的酶,1901年該酶被正式命名為過氧化氫酶(catalase,CAT),之后通過實驗證實卟啉環是CAT的活性中心[14].這2個基因主要在抗氧化系統中起到了清除過氧化氫的作用,保護生物體免受過氧化氫過量積累所帶來的損傷[15].
本文克隆了真蛸GPX(OvGPX)和真蛸CAT(OvCAT)基因全長,研究了2個基因氨基酸結構特性,以及其在高溫條件下的表達特征,以期詮釋真蛸抗氧化酶蛋白的功能,揭示環境脅迫下OvGPX和OvCAT蛋白對于真蛸的保護作用,為真蛸的良種選育和資源保護等方面提供基礎資料和理論依據.
1 材料與方法
1.1 實驗動物的暫養和取樣
本文所用的真蛸來自福建省霞浦縣北壁海區,帶回實驗室以活蟹和貝類喂食、蓄養1周(水溫22~24℃,鹽度26~27)后:1)選活潑正常的真蛸,取其消化腺置于由百泰克公司購買的RNA fixer保存液中,4℃過夜后移入-20℃冰柜保存、備用;2)選28只健康活潑的個體,經相應水溫馴化恒定后,分別暫養在4個溫度組(24,26,28,30℃)中,每個溫度組7只個體,2h后各組隨機取3只個體,分別取其消化腺和櫛鰓,同上處理保存備用.
1.2 RNA提取、cDNA合成和基因全長的獲得
總RNA提取利用Trizol方法,Trizol試劑購自TaKaRa公司,具體實驗步驟按說明書進行.提取的RNA樣品存放于-80℃冰箱中保存,采用ND1000微量紫外-可見分光光度計檢測RNA的純度和濃度;RNA第1鏈反轉錄采用TaKaRa公司的Reverse Transcriptase M-MLV 試 劑 盒 進 行,利 用 Oligo dT-RA為反轉錄引物,反轉錄實驗根據說明書操作,所得cDNA用作3′RACE模板.
反轉錄后的第1鏈cDNA,用NEB公司的RNahase試劑在37℃降解1h,加入2倍體積乙醇于-20℃ 靜置30min,4℃離心沉淀30min,加入超輕水(DDW)溶解,作為5′加尾的模板.5′加尾反應體系為:19μL cDNA溶液,依次加入5×TdT緩沖液10μL,TdT 酶 (15U/μL)1μL,dATP(0.5mmol/L)2.5 μL,0.1%(質量分數)牛血清白蛋白(BSA)5μL,加DDW補齊至50μL,試劑均購自TaKaRa公司,37℃溫育30min,所得即為5′端模板.
利用cDNA 末端快速克隆(rapid amplification of cDNA ends,RACE)[16]的方法對 OvGPX 和 OvCAT基因全長進行克隆,基因片段序列源自本實驗室構建的真蛸神經系統轉錄組文庫[17],片段用于進一步擴增.設計參與3′RACE的特異性引物分別為GPX-F1、GPX-F2、CAT-F1、CAT-F2;5′RACE 的反應特異性引物為 GPX-R1、GPX-R2、CAT-R1、CAT-R2(引物序列詳見表1).
PCR產物經瓊脂糖凝膠電泳后,利用鷺隆公司的凝膠回收試劑盒進行膠回收;將純化后的PCR產物利用TaKaRa公司的pMD19-T載體進行克隆,用DH5α感受態細胞進行轉化,感受態細胞試劑盒購自鷺隆公司,具體操作步驟根據說明書進行.轉化后氨芐培養基培養16h,隨機挑選6個克隆,PCR反應和瓊脂糖凝膠檢測后送南京金斯瑞公司測序.
1.3 序列分析和多重序列比對
通過 Blast(http:∥blast.ncbi.nlm.nih.gov/Blast.cgi)查找同源序列,利用ClustalX和Dnaman軟件進行多序列同源比對;以ORFfinder(http:∥www.ncbi.nlm.nih.gov/gorf/orfig.cgi)進行開放閱讀框(ORF)查找;在線 Protparam (http:∥web.expasy.org/cgi-bin/protparam/protparam)軟件分析氨基酸序列組成、分子質量和等電點;Smart在線軟件、Interproscan在線軟件預測編碼蛋白的基因特征基序、功能域(http:∥www.ebi.ac.uk/Tools/pfa/iprscan/、http:∥smart.embl-heidelberg.de/);利用 SECISearch2.19在 線 軟 件 (http:∥genome.unl.edu/SECISearch.html)預測GPX是否存在硒半胱氨酸(Sec)插入序列(SECIS)結構;利用 NetNGlyc軟件(http:∥www.cbs.dtu.dk/services/NetNGlyc/)預測GPX和CAT糖基結合位點.
1.4 實時定量PCR(qPCR)檢測不同溫度下OvGPX和OvCAT的基因表達變化
為了解OvGPX和OvCAT基因在不同溫度應激下的表達變化,本文以不同溫度組的消化腺和櫛鰓的cDNA第1鏈為模板進行qPCR反應.RNA提取和反轉錄反應分別采用百泰克公司的RNA Plus試劑盒和TaKaRa公司的 PrimeScript?RT reagent Kit試劑盒,按說明書進行實驗操作.應用TaKaRa公司的SYBR Premix Ex TaqTM II試劑盒,在 Rotor-gene3000熒光實時定量熱循環儀上進行qPCR反應,實驗步驟參照說明書進行.合成基因特異性引物GPX-F、GPX-R、CAT-F和 CAT-R用于該實驗,以β-actin 基因作為qPCR的內參基因(序列見表1).根據qPCR給出的Ct值,取3個平行樣本的平均值,利用2-ΔΔCt方法計算2個基因mRNA的相對表達量.

表1 實驗用引物表Tab.1 Primers used in this study
1.5 統計分析
利用SPSS 13.0軟件對所得數據進行單因素方差分析,數據以平均值±標準差的形式,顯著性分析水平為p<0.05,并做柱狀圖.
2 結 果
2.1 OvGPX和OvCAT基因序列分析
以RACE方法,拼接得到OvGPX基因.得到的OvGPX cDNA全長為925bp,包含5′非編碼區(5′UTR)36bp,3′非編碼區(3′UTR)316bp,ORF 573 bp,共編碼190個氨基酸,推測分子質量21.5ku,等電點pI為7.81.OvGPX推導的氨基酸序列(圖1)含有GPX家族2個典型酶活性位點基序,分別是從28GKVILVENVASLUGTT43和64LGFPCNQF71;并含有特有的Sec,利用SECISearch2.19在線軟件在OvGPX基因的3′UTR處預測到1個SECIS,全長91bp(圖2),含1個頂環和1個AG-GA SECIS核,頂環處存在保守的AAA序列.
得到的 OvCAT 全長為2 109bp,包含5′UTR 135bp、3′UTR 435bp和ORF 1 539bp,共編碼512個氨基酸,推測分子質量58.3ku,等電點pI為8.76(圖3).通過功能域預測,推導的OvCAT氨基酸序列含有1個過CAT活性位點(61FNRERIPERVVHAKGAG77)、1個近端血紅素配體標簽序列(351RLYSYSDT358)和1個酶免疫響應區(位點428~494).并發現了3個參與過氧化氫酶催化位點(His-71,Asn-144和 Tyr-354),同時預測到了3個糖基結合位點NASL、NYSQ、NVSS.

圖1 OvGPX的核苷酸和推導氨基酸序列Fig.1 Nucleotide and deduced amino acid sequences of OvGPX cDNA

圖2 OvGPX基因的SECIS元件示意圖.2The SECIS functional element of OvGPX
2.2 OvGPX和OvCAT基因氨基酸序列同源性分析
通過Blast、Clustal X和DNAman等分析軟件,將OvGPX推導的氨基酸序列與8種脊椎動物和無脊椎動物的GPX氨基酸序列進行多序列分析(圖4).結果表明OvGPX氨基酸序列高度保守,所有比對序列基本都含有GPX的2個酶活性位點基序,且都有Sec.與曼氏無針烏賊(Sepiella maindroni)的 GPX氨基酸序列同源性最高(74%),與人類(Homo sapiens)的GPX氨基酸序列同源性也達60%.
將本實驗得到的OvCAT的氨基酸序列與8種已有動物的CAT氨基酸序列進行多重序列比對(圖5).結果顯示,所比對物種的CAT氨基酸序列高度保守,基本都含有CAT的特征基序和3個參與催化的殘基.比對結果顯示,不同物種相似性較高,OvCAT氨基酸序列與同是軟體動物門的香港巨牡蠣(Crassostrea hongkongensis)和 盤 鮑 (Haliotis discus)的CAT氨基酸序列相似性高達76%和75%,與人類的CAT氨基酸序列相似性也有68%.

圖3 OvCAT的核苷酸和推導氨基酸序列Fig.3 Nucleotide and deduced amino acid sequences of OvCAT cDNA

圖4 OvGPX氨基酸序列比對Fig.4 Alignment of OvGPX amino acid sequences
2.3 OvGPX和OvCAT基因mRNA在不同溫度下的表達分析
以qPCR方法分析了OvGPX基因在24,26,28和30℃不同溫度組處理2h后消化腺和櫛鰓的mRNA表達量差異(圖6).
在消化腺和櫛鰓中,OvGPX mRNA表達量均在28℃組最高,分別達到24℃組的7.57倍(p<0.01)和2.19倍.在30℃組,櫛鰓OvGPX mRNA的表達水平下降,只有24℃組的1/3(p<0.05);而消化腺中OvGPX mRNA的表達量與室溫組沒有顯著差異.
不同溫度組處理2h,真蛸消化腺和櫛鰓中OvCAT mRNA的表達量差異如圖7,在消化腺和櫛鰓中,28℃組OvCAT mRNA表達量明顯升高,分別達到24℃的2.36倍(p<0.01)、2.13倍(p<0.01);在消化腺中26和30℃組的OvCAT mRNA表達量與24℃組無顯著差異;櫛鰓的OvCAT mRNA表達水平在30℃組低于24℃組,為24℃組的0.57.
不同溫度組qPCR結果表明,OvGPX和OvCAT 2個基因的表達水平在不同溫度組的表達差異相似,但是OvGPX的mRNA表達量變化波動較OvCAT明顯.
3 討 論
本文研究得到的OvGPX cDNA全長,除了具有2個高度保守的特征基序,預測的氨基酸序列中還發現了由UGA編碼的Sec U,這是GPX的中心催化基團,能夠催化GSH分解體內的氫過氧化物,從而防止細胞膜和其他生物組織遭受過氧化損傷[12-13].UGA在一般情況下作為終止密碼子,但在OvGPX基因中,其主要編碼被稱為第21種氨基酸的Sec[18].該編碼依賴硒元素,當細胞生長缺乏硒時,硒蛋白的翻譯會在UGA密碼子處中止,成為不完整而沒有功能的蛋白;而破譯UGA使其編碼Sec需要SECIS元件[19]的作用,SECIS元件是一個位于GPX基因3′UTR部分的一個莖-環發卡結構,本文得到的OvGPX基因,在3′UTR區同樣發現了SECIS序列,這段序列由螺旋莖和頂環、內環組成,在莖的基部具有保守的AUGAC和UGAC形成的形成非 Watson-Crick雙鏈,結構完整,這個完整的SECIS元件保證了OvGPX基因的正確轉錄.SECIS元件的分類主要根據保守的AAA序列是否位于頂環進行判斷,本研究得到的OvGPX基因的SECIS元件,AAA序列位于頂環,與人類的OvGPX1一樣屬于一類[20-21].
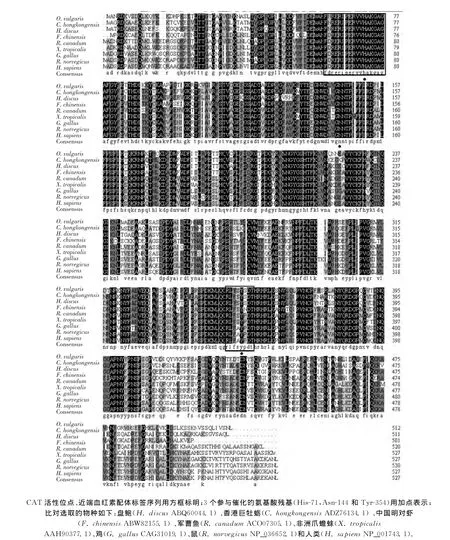
圖5 OvCAT氨基酸序列比對Fig.5 Alignment of OvCAT amino acid sequences

圖6 qPCR方法下的OvGPX在不同溫度的表達量Fig.6 The expression of OvGPX at different temperatures by qPCR
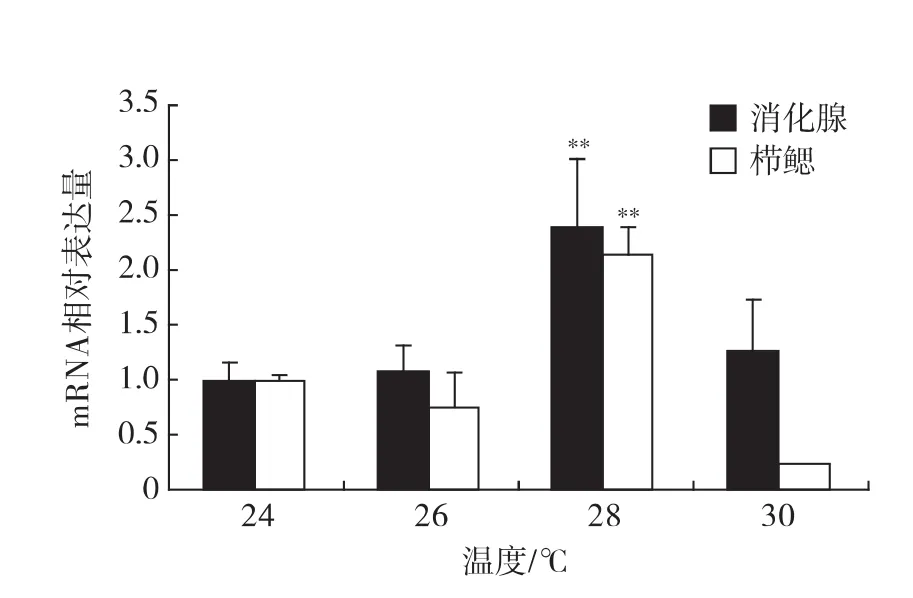
圖7 qPCR方法下的OvCAT在不同溫度的表達量Fig.7 The expression of OvCAT at different temperatures by qPCR
水生動物受到熱應激后的抗氧化酶的活性變化有所差異,翡翠貽貝(Perna viridis)熱激后,SOD、CAT、GST、GPX等酶活力都明顯的高于對照組[22];雙線血蛤((Hiatula diphos)和中國血蛤(Hiatula chinensis)在20~70℃不同的水溫刺激下,CAT酶活性變化趨勢表現為先上升,后下降,且雙線血蛤的變化幅度比中國文蛤明顯[23];在對盤鮑的28℃熱應激研究中發現,GPX、CAT和SOD 3個基因的mRNA表達量在熱應激后均上調[24].可見,熱激強度不同和熱激時間的長短,對抗氧化酶活性造成的變化并不相同,而同一實驗條件下,不同物種的GPX或CAT酶活性變化也有差異.這些結果說明GPX和CAT這2種抗氧化酶參與熱應激后的自由基清除活動,保證了機體免受過度自由基損傷.本實驗中,不同的溫度應激后,消化腺中的OvGPX和OvCAT基因的mRNA表達量在應激條件下均有上升,體現了2個抗氧化基因在熱應激壓力下清除過氧化氫,保護機體的積極作用;櫛鰓OvGPX和OvCAT的mRNA表達量在28℃組有所升高,26和30℃組表達量下降,說明消化腺和櫛鰓抗氧化能力可能有所不同.本文不同強度的熱應激實驗結果顯示,OvGPX mRNA表達量的變化幅度明顯大于OvCAT,顯示了OvGPX對氧化應激較為敏感.
本文克隆了OvGPX和OvCAT基因全長,并探索2個基因在不同熱激強度下的表達情況,初步確定2個基因參與熱應激脅迫后的抗氧化反應.這些數據為進一步研究OvGPX和OvCAT的生物學功能提供有效信息,并為深入探索頭足類動物的抗氧化體系奠定理論基礎.
[1]Navarro J C,Villanueva R.The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food:deviation from their natural fatty acid profile[J].Aquaculture,2003,219:613-631.
[2]Iglesias J,Otero J J,Moxica C,et al.The completed life cycle of the octopus (Octopus vulgaris,Cuvier)under culture conditions:paralarval rearing using Artemia and zoeae,and first data on juvenile growth up to 8months of age[J].Aquaculture International,2004,12 (4/5):481-487.
[3]徐實懷,馬之明,賈曉平,等.人工養殖條件下真蛸的生物學特性及胚胎發育[J].南方水產,2009,5(2):63-68.
[4]Delgado M,Gairin J I,Carbo R,et al.Growth of Octopus vulgaris (Cuvier,1797)in tanks in the Ebro Delta(NE Spain):effects of temperature,salinity and culture density[J].Scientia Marina,2011,75:53-59.
[5]Chilmonczyk S,Monge D.Rainbow trout gill pillar cells:demonstration of inert particle phagocytosis and involvement in viral infection[J].Journal of the Reticuloendothelial Society,1980,28(4):327-332.
[6]Halliwell B.Free radicals,antioxidants,and human disease:curiosity,cause or consequence [J].Lancet,1994,344:721-724.
[7]Wiseman H,Halliwell B.Damage to DNA by reactive oxygen and nitrogen species:role in inflammatory disease and progression to cancer[J].The Biochemical Journal,1996,313:17-29.
[8]Gotia S,Popovici I,Hermeziu B.Antioxidant enzymes levels in children with juvenile rheumatoid arthritis[J].Rev Med Chir Soc Med Nat Iasi,2001,105(3):499-503.
[9]Cetinkaya O,Silig Y,Cetinkaya S,et al.The effects of Rumex patientia extract on rat liver and erythrocyte antioxidant enzyme system[J].Die Pharmazie,2002,57(7):487-488.
[10]Muruganandan S,Gupta S,Kataria M,et al.Mangiferin protects the streptozotocin-induced oxidative damage to cardiac and renal tissues in rats[J].Toxicology,2002,176(3):165-173.
[11]Mills G C.Hemoglobin catabolism.I.Glutathione peroxidase,an erythrocyte enzyme which protects hemoglobin from oxidative breakdown[J].The Journal of Biological Chemistry,1957,229(1):189-197.
[12]Flohe L,Genzler W A,Schock H H.Glutathione peroxidase:a selenoenzyme[J].FEBS Letters,1973,32(1):132-134.
[13]Rotruck J T,Pope A L,Ganther H E,et al.Selenium:biochemical role as a component of glutathione peroxidase[J].Science,1973,179:588.
[14]劉冰,梁嬋娟.生物過氧化氫酶研究進展[J].中國農學通報,2005,21(5):223-224.
[15]Nordberg J,Arner E S J.Reactive oxygen species,antioxidants and the mammalian thioredoxin system [J].Free Radical Biology & Medicine,2001,31:1287-1312.
[16]薩姆布魯克J,拉塞爾 D W,著.分子克隆實驗指南[M].3版.黃培堂,等譯.北京:科學出版社,2002.
[17]Zhang X,Mao Y,Huang Z,et al.Transcriptome analysis of the Octopus vulgaris central nervous system[J].PLoS One,2012,7(6):e40320.
[18]Stadtman T C.Selenocysteine[J].Annual Review of Biochemistry,1996,65:83-100.
[19]Fagegaltier D,Lescure A,Walczak R,et al.Structural analysis of new local features in SECIS RNA hairpins[J].Nucleic Acids Research,2000,28(14):2679-2689.
[20]Grundener-Culemann E,Martin G W,Harney J W,et al.Two distinct SECIS structures capable of directing selenocysteine incorporation in eukaryotes[J].RNA,1999,5(5):625-635.
[21]Kryukov G V,Castellano S,Novoselov S V,et al.Characterization of mammalian selenoproteomes[J].Science,2003,300(5624):1439-1443.
[22]Verlecar X N,Jena K B,Chainy G B.Biochemical markers of oxidative stress in Perna viridis exposed to mercury and temperature[J].Chemico-biological Interactions,2007,167(3):219-226.
[23]孫業勇,高如承,溫揚敏.溫度對中國血蛤、雙線血蛤肝臟中水解酶和抗氧化酶活性的影響[J].水產科學,2008,10(27):543-544.
[24]De Zoysa M,Whang I,Lee Y,et al.Transcriptional analysis of antioxidant and immune defense genes in disk abalone(Haliotis discus discus)during thermal,low-salinity and hypoxic stress[J].Comparative Biochemistry and Physiology:Part B,2009,154(4):387-395.
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
