海州灣條斑紫菜養殖對浮游藻類群落結構及遺傳多樣性的影響
2014-12-15 03:35:04王洪斌李信書宋秀梅李士虎閻斌倫
海洋科學 2014年12期
王洪斌 , 李信書 宋秀梅 吳 言 李士虎 閻斌倫
(1.淮海工學院 海洋學院, 江蘇 連云港 222005; 2.江蘇省海洋生物技術重點實驗室, 江蘇連云港 222005)
在水生態系統中, 浮游藻類是海洋生態系統初級生產力代表, 也是海洋生態系統中食物鏈的重要環節,其種類組成、數量變化往往也是所處環境綜合作用的結果, 浮游藻類對外部環境因子的變化非常敏感[1]。因此, 浮游藻類群落組成的改變能在一定程度上反映水環境的狀態。目前, 越來越多的研究開始關注浮游藻類與環境因子之間的關系[2]。核酮糖1, 5-二磷酸羧化/氧化酶( RubisCO)是浮游藻類光合作用的關鍵酶,具有催化 CO2還原和 1, 5-二磷酸核酮糖氧化的雙重功能, 廣泛存在于藻類和高等植物中[3]。RubisCO 大亞基基因(rbcL) 是研究物種間系統學關系比較恰當的標志基因,rbcL基因沒有內含子結構, 可以直接PCR, 廣泛應用于浮游藻類系統學及多樣性研究[4]。
海州灣位于江蘇省北部連云港沿海, 近年來,由于城市化進程的快速發展, 大量含有高濃度氮、磷等營養物的污染物排入沿海, 使得近海的富營養化問題十分嚴重, 赤潮頻繁發生[5]。以往也有不少學者,對海州灣的理化因子與浮游植物的數量變化關系進行了研究[6], 并發現浮游植物的數量變化與環境因素的變化密切相關。而針對條斑紫菜養殖行為導致水質條件改變, 從而引起浮游藻類群落結構及組成的變化研究未見報道。
作者分別提取了江蘇海州灣條斑紫菜非養殖區及養殖區的浮游藻類總DNA, PCR方法擴增浮游藻類rbcL片段, 構建了條斑紫菜非養殖區及養殖區浮游藻類rbcL片段質粒文庫, 根據rbcL序列分析, 研究了海州灣條斑紫菜養殖對浮游藻類遺傳多樣性及群落結構的研究。
1 材料與方法
1.1 實驗樣品來源及試劑
2012年12 月, 于江蘇省海州灣近海條斑紫菜非養殖海區(1、3、5、7 )和養殖海區( 2、4、6、8 )采集表層海水, 共 8個樣點 (圖1)。于每個樣點(距水面0.2~0.3 m處)取200 mL表層水, 分裝在已滅菌的PVC瓶中, 所取水樣置于保溫箱中立刻運往實驗室,于4℃黑暗保存, 且所有樣品都在一周內進行分析。
PCR引物合成、測序由上海桑尼生物科技有限公司完成; pMD18-T載體購自寶生物(大連)工程有限公司;Escherichia coliJF1125由本實驗室保存; 其他分子生物學試劑均為分析純。
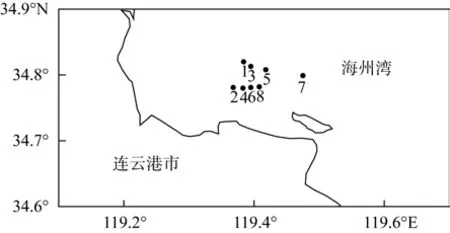
圖1 采樣點位置示意圖Fig.1 Map of sampling sites
1.2 海水中浮游藻類總DNA的提取
養殖海區(2、4、6、8)各采樣點分別取水樣100 mL混勻, 非養殖海區水樣處理同養殖區。400 mL冷藏水樣10 000 r/min、離心10 min, 棄上清, 沉淀中加入460 μL裂解緩沖液(250 mmoL Tris-HCl pH 8.0,60 mmoL EDTA), 340 μL 10%SDS, 蛋白酶 K(終質量濃度 0.2 mg/mL)15 μL, 37 ℃溫浴 3 h, 10 000 r/min、離心10 min, 上清轉至潔凈1.5 mL離心管, 等體積酚氯仿抽提, 12 000 r/min、離心1 min, 上清液用2倍體積的異丙醇4 ℃沉淀過夜, 12 000 r/min、離心10 min, 棄上清, 沉淀用50 μL TE緩沖液溶解, -20 ℃保存[7]。
1.3 PCR擴增rbcL基因及rbcL質粒文庫構建
以海水浮游藻類總DN A為模板, PCR法擴增獲得海洋浮游藻類rbcL片段。根據Ⅰ型 RubisCO 大亞基氨基酸保守區( KPKLGLS 和 VVGKLEG)[8]設計PCR 引物。引物序列分別為5′-AA(AG)CC(TA)AA(AG)(TC)TAGG(TG)(CT)T(AT)TC-3′和 5′-CC(TC)TC(TC)A(AG)(TC) TTACC(AT)AC(GAT)AC-3′。PCR 反應體系: 模板DNA為50 ng(1.5 μL), dNTP為0.2 mmol ,正反向引物各為 0.2 μmol, Taq 酶 2 unit, buffer 5 μL,50 μL反應體系[9]。 PCR反應條件: 95 ℃預變性5 min,94 ℃變性1 min, 48 ℃復性2 min, 72 ℃延伸1 min,72 ℃終止延伸10 min, 35循環。PCR 產物割膠回收,與pMD18-T載體( TaKaRa公司)連接, 轉化感受態細胞E.coliJF1125, 獲得rbcL片段質粒文庫。隨機挑選重組子, 抽提質粒, 由上海桑尼生物科技有限公司完成測序。經序列比對并用鄰接法構建系統進化樹[10]。
2 結果與分析
2.1 條斑紫菜養殖對浮游藻類群落結構的影響
PCR產物約300bp, 構建浮游藻類rbcL片段質粒文庫(養殖區 DrbcL, 非養殖區 CrbcL)。文庫中隨機選擇 50個陽性克隆, 測定序列, 并用鄰接法構建系統進化樹(圖2, 圖3), 根據序列比對及進化樹顯示的遺傳距離分析確定養殖及非養殖海區的浮游藻類的群落結構組成。本研究使用的PCR引物是Ⅰ型rbcL基因特異的, 而甲藻rbcL基因屬于Ⅱ型[9]。因此, 本研究使用的引物無法擴增甲藻rbcL基因。結果顯示, 非養殖區發現8種海洋微藻, 分別為隱鞭藻(Cryptomonas)、異彎藻(Heterosigma)、塔胞藻(Pyramimonas mantoriae chicroplast)、全溝藻(Teleaulax amphioxeia)、筒柱藻(Cylindrotheca)、小球藻(Chlorella)、海鏈藻(Thalassiosira)、骨條藻(Skeletonema)。養殖區發現 10種海洋微藻, 分別為異絲藻(Heterothrix debilis)、杜氏藻(Dunaliella)、 隱鞭藻(Cryptomonas)、骨條藻(Skeletonema)、小球藻(Chlorella)、菱形藻(Nitzschia longissima)、嚎殼藻(Ostreobium quekettii)、鏈帶藻(Desmodesmus pseudoserratus)、小環藻(Cyclotella)、擬伊藻(Ahnfeltiopsis leptophylla)。海洋中浮游藻類種類繁多, 影響其種類及豐度的主要因素為浮游藻類自身消長的特性、水體條件及浮游動物等的捕食。浮游藻類的豐度及組成受紫菜養殖的影響, 如骨條藻在養殖區為優勢種群, 但在非養殖區卻變為劣勢種群。養殖區和非養殖區雖然只隨機抽取了50個序列, 但是這些序列至少覆蓋了一個優勢種群, 如養殖區的異絲藻及非養殖區的隱鞭藻。隱鞭藻和小球藻為兩海區所共有的優勢類群且覆蓋克隆數比例均較高, 說明小球藻與隱鞭藻的適應性強。因紫菜養殖和非養殖區相距較近,海水隨潮汐和風浪而流動, 所以紫菜養殖和非養殖區的物化及環境生態因子既有差異也有相同之處。已有實驗證實, 紫菜養殖和非養殖區因紫菜光合作用導致營養鹽存在顯著性差異[11-12], 這種差異勢必對浮游藻類群落結構產生影響。
2.2 條斑紫菜養殖對浮游藻類遺傳多樣性的影響
條斑紫菜養殖及非養殖海區浮游藻類類群覆蓋克隆數見表1。依據覆蓋克隆數的百分比, 非養殖區8種海洋微藻為隱鞭藻(22%)、異彎藻(18%)、塔胞藻(14%)、全溝藻(13%)、筒柱藻(12%), 小球藻(12%),海鏈藻(6%), 骨條藻(2%), 見圖2。隱鞭藻、異彎藻在非養殖區所占比例較大, 含量豐富; 海鏈藻, 骨條藻多占比例較小。養殖區 10種海洋微藻為異絲藻(22%)、杜氏藻(12%)、 隱鞭藻(12%)、骨條藻(12%)、小球藻(10%)、菱形藻(8%)、嚎殼藻(6%)、鏈帶藻(6%)、小環藻(6%)、擬伊藻(2%), 見圖3。異絲藻在養殖區所占比例較大, 優勢度明顯; 鏈帶藻, 菱形藻等類群占比例差不多, 為6%~12%。如果將覆蓋克隆數大于5(10%) 的類群定義為優勢類型, 那么非養殖區的優勢類群有隱鞭藻(22%), 異彎藻(18%), 塔胞藻(14%)全溝藻(13%), 小球藻(12%), 筒柱藻(12%),養殖區的優勢類群有異絲藻(22%), 杜氏藻(12%),骨條藻(12%), 隱鞭藻(12%), 小球藻(10%)。優勢類群的形成不僅僅決定于營養、溫度等環境條件, 還與物種生理特性相關, 物種間競爭也會影響到優勢物種的形成[13]。15種浮游藻類類群中只有 3種(隱鞭藻, 骨條藻及小球藻)為兩海區共有, 其他浮游藻類類群均為不同區域所有, 證實條斑紫菜養殖區及非養殖區浮游藻類群落組成差異顯著。杜氏藻、異絲藻為養殖區所特有的, 異絲藻含量極高, 養殖區內的其他幾類浮游藻類類群的含量接近, 異彎藻,全溝藻, 塔胞藻, 屬于非養殖區特有優勢類群。兩海區共有的浮游藻類類群中骨條藻的豐度差異極大,在非養殖區所占比例極小; 小球藻與隱鞭藻在兩個區域中的豐富度都較高, 可能由于隱鞭藻和小球藻對溫度、營養條件及其他物化因子具有較強的適應能力, 廣泛存在于各個區域, 并且形成優勢類群。而非共有的 12個浮游藻類類群對環境因子需求較高,適應力較窄, 只有在特定水體條件下才能夠大量繁殖, 如異絲藻只在紫菜養殖區大量存在, 非養殖區沒有, 表明條斑紫菜養殖區的水體環境等因素適宜此類浮游藻類生長繁殖, 當生長條件適宜的時候, 具有競爭優勢的物種將快速繁殖, 大量的利用營養物質,抑制其他藻類的生長而迅速成為優勢種群。至于異絲藻與條斑紫菜養殖耦合的生態條件有待進一步研究。
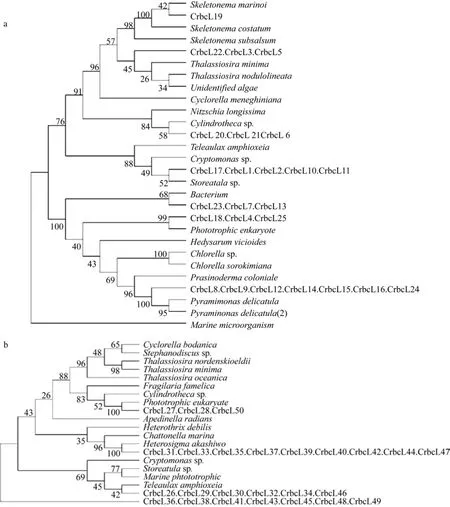
圖2 非養殖區浮游藻類進化樹Fig.2 System evolution tree of phytoplankton in natural region
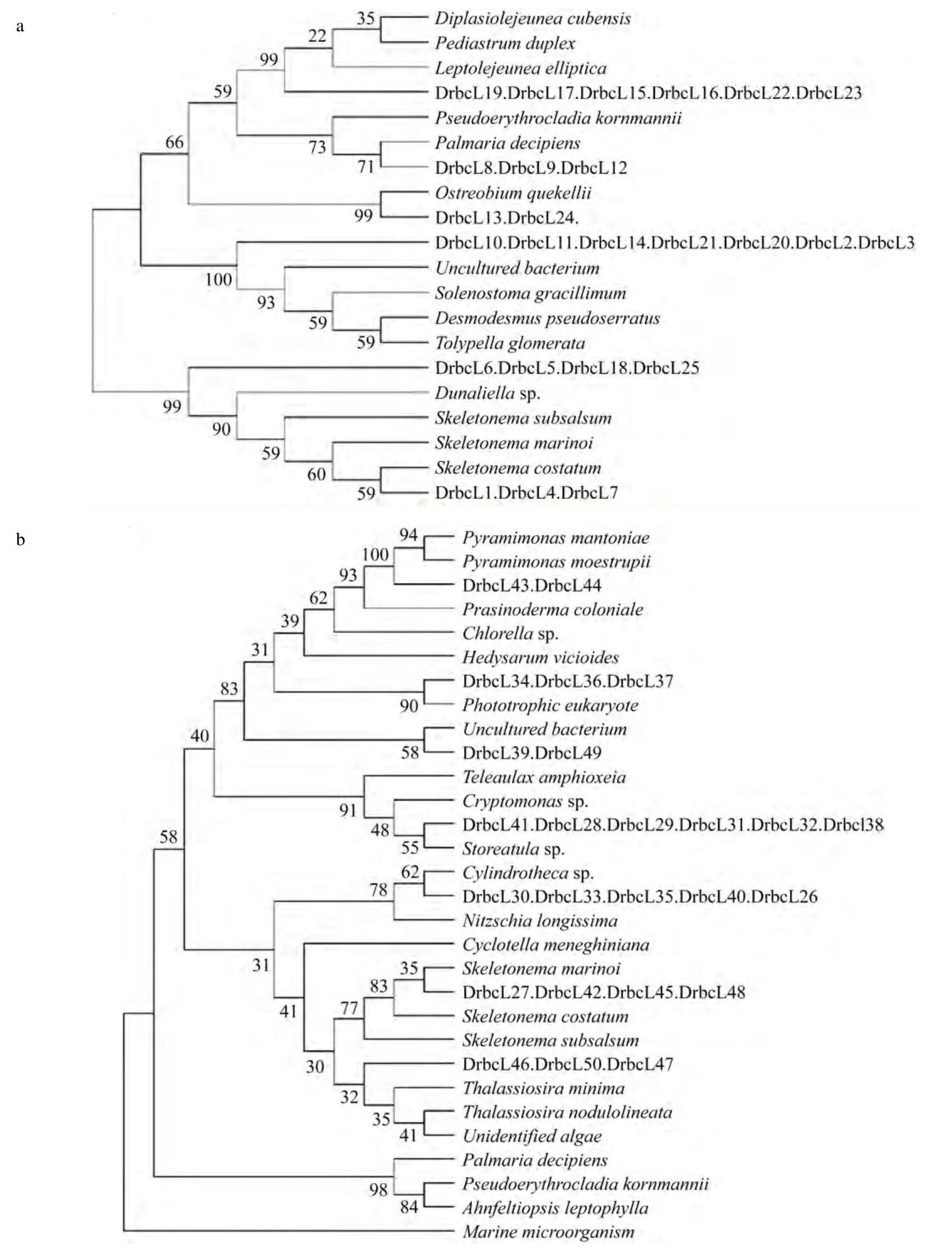
圖3 養殖區浮游藻類進化樹Fig.3 System evolution tree of phytoplankton in mariculture zone
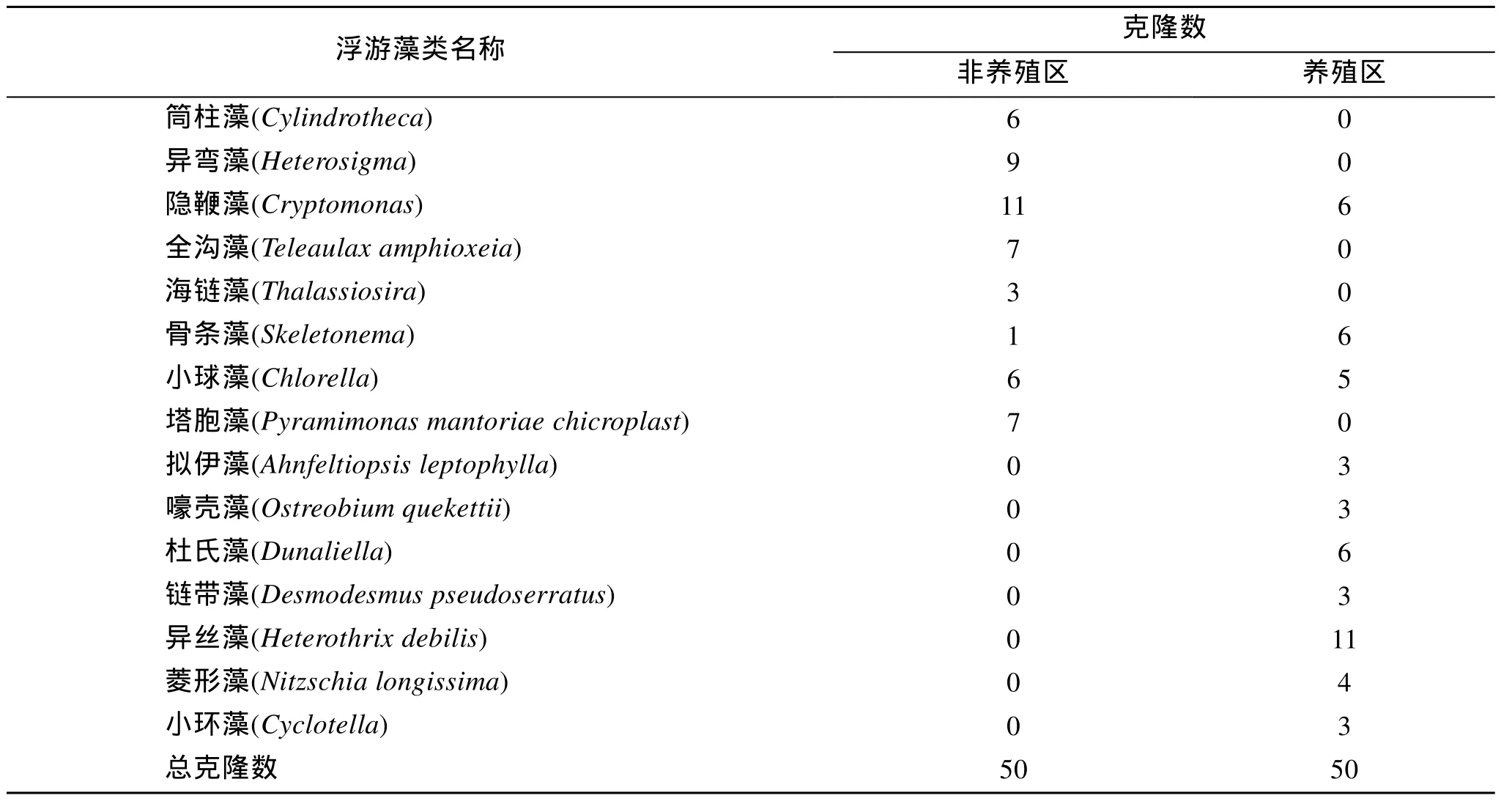
表1 海州灣養殖和非養殖海區浮游藻類分布Tab.1 Distribution of Phytoplankton in mariculture zone and natural region in Haizhou bay
2.3 條斑紫菜養殖對浮游藻類豐度的影響
物種多樣性指的是群落中物種數目的多少, 它是衡量群落的規模和重要性的基礎, 而以往研究多側重于營養鹽濃度對浮游植物的上行效應, 近年來,隨著研究深入, 浮游植物多樣性對營養鹽結構變化的響應也引起了國內外學者的廣泛關注[14-15]。表2顯示, 非養殖區的香農指數小于 3的浮游藻類類群,占了總類群的 50%, 而養殖區的香農指數小于 3的只占1個, 占了總類群的10%, 養殖及非養殖海區的香農指數均值為 3.654/3.273, 養殖區的浮游藻類香農指數普遍高于非養殖區, 養殖區生物多樣性豐富,與上述結論一致。其原因可能是條斑紫菜養殖行為導致本海區的物化及其他環境生態改變, 特別是N/P的變化, 營養鹽結構改變會導致浮游植物群落結構發生變化, 影響到海洋生態系統的多樣性和穩定性[16]。非養殖區的均勻度指數差異較大, 養殖區的均勻度指數差異較小, 養殖區平均均勻度為 1.040,非養殖區的平均均勻度為 1.090, 養殖區的平均均勻度指數小于非養殖區, 可見非養殖區的浮游藻類的物種分布較為均勻, 群落的成熟度與穩定性比養殖區高。說明不同種類浮游植物的營養鹽利用方式及生長勢的差異是影響其群落競爭格局的關鍵因素[17],非養殖區因無條斑紫菜存在, 海區物化及各種環境因子較穩定, 浮游藻類群落的穩定性較高。
3 結論
作者運用PCR法擴增浮游藻類核酮糖1, 5-二磷酸羧化/氧化酶大亞基(rbcL) 基因片段, 構建了條斑紫菜養殖及非養殖海區浮游藻類的rbcL片段質粒文庫。隨機選擇文庫50個克隆并測序, 經比對分析,條斑紫菜非養殖區發現 8種海洋微藻, 分別為隱鞭藻、異彎藻、塔胞藻、全溝藻、筒柱藻, 小球藻, 海鏈藻, 骨條藻。養殖區共發現 10種海洋微藻, 分別為: 異絲藻、杜氏藻、 隱鞭藻、骨條藻、小球藻、菱形藻、嚎殼藻、鏈帶藻、小環藻、擬伊藻。
浮游藻類類群中只有 3種(隱鞭藻, 骨條藻及小球藻)為兩個海區共有, 其他浮游藻類類群均為不同海區所有, 證實條斑紫菜養殖及非養殖海區浮游藻類群落組成差異顯著。杜氏藻、異絲藻為養殖區所特有的, 異絲藻優勢極高, 養殖區內的其他幾類浮游藻類類群的含量接近; 異彎藻, 全溝藻, 塔胞藻,屬于非養殖區特有優勢類群。
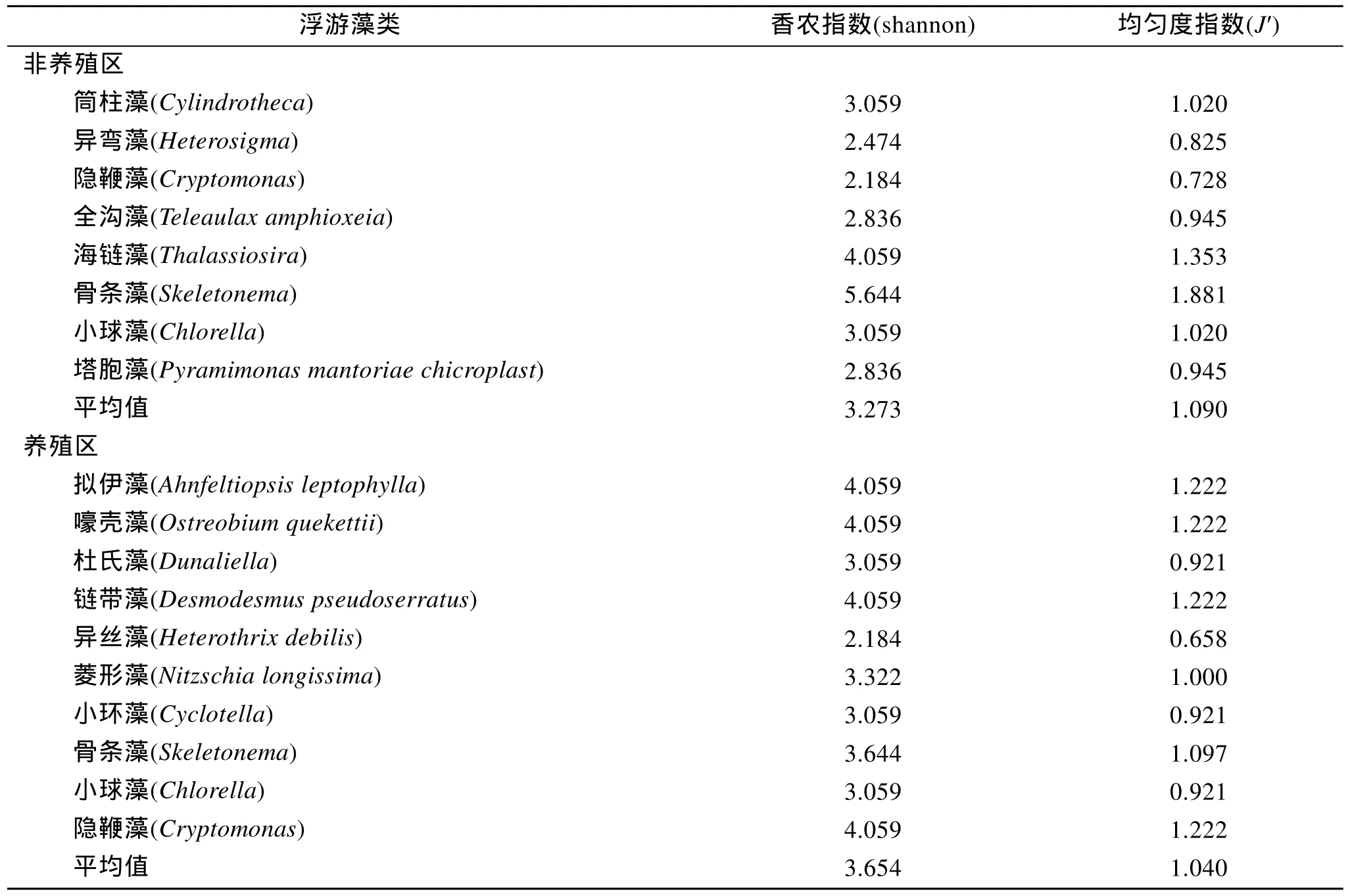
表2 海州灣養殖和非養殖區浮游藻類多樣性Tab.2 Genetic diversity of Phytoplankton in mariculture zone and natural region in Haizhou bay
實驗證實, 條斑紫菜養殖海區浮游藻類遺傳多樣性豐富, 而群落的成熟度與穩定性以非養殖區為高。說明表層海水浮游藻類群落結構及組成是動態的, 條斑紫菜的養殖改變了浮游藻類群落結構及遺傳多樣性。養殖海區浮游藻類的多樣性及豐富度高于非養殖海區, 而且養殖海區出現的優勢藻類均為海州灣非赤潮優勢藻[18], 多樣性豐富降低了單一海洋微藻因富營養化而導致的爆發性增長的可能, 減少赤潮發生的頻率。浮游藻類群落結構及遺傳多樣性改變對于條斑紫菜養殖行為使海域生態環境的修復具有重要意義。
[1]Paerl H W, Dyble J, Moisander P H, et al.Microbial indicators of aquatic ecosystem change: current applications to eutrophication studies [J].Microbiological Ecology, 2003, 46: 233-246.
[2]Liu Y Q, Yao T D, Zhu L P, et al.Bacterial diversity of freshwater alpine lake Puma Yumco on the Tibetan Plateau[J].Geomicrobiology Journa, 2009, 26: 131-145.
[3]Ellis R J.The most abundant protein in the word [J].Trends Biochem Sci, 1979, 4(1) : 241-244.
[4]Clegg M T.Chloroplast gene sequences and the study of plant evolution [J].Proc Natl Acad Sci USA, 1993,90(2): 363-367.
[5]黃偉, 朱旭宇, 曾江寧, 等.氮磷比對浙江近岸浮游植物群落結構影響的實驗研究[J].海洋學報, 2012,34(5): 128-138.
[6]徐虹, 黃祖英, 魏愛泓.海州灣浮游植物總量的多元分析[J].海洋環境科學, 2009(增刊), 28(1): 26-27.
[7]Polen-fuller M.A two-hour method for extraction of DNA from seaweeds[J].Phycol ogical Newsletter, 1991,23: 2.
[8]劉瑞玉.膠州灣生態學和生物資源[M].北京: 科學出版社, 1992.
[9]劉永健, 楊官品, 管曉菁, 等.膠州灣浮游植物遺傳多樣性及其季節變化研究[J].海洋學報, 2005, 27(5):103-111.
[10]孫力軍, 陸兆新, 別小妹, 等.1株抗菌植物內生菌EJH-2菌株的分離和鑒定[J].中國微生態學雜志,2006, 18(1): 23-26.
[11]陳聚法, 趙俊, 過鋒, 等.條斑紫菜對膠州灣濕地淺海富營養化狀況的生物修復效果[J].漁業科學進展,2012, 33(1): 93-101.
[12]張寒野, 何培民, 陳嬋飛 , 等.條斑紫菜養殖對海區中無機氮濃度影響[J].環境科學與技術, 2005,28(4): 44-45.
[13]王悠, 俞志明, 宋秀賢, 等.大型海藻與赤潮微藻以及赤潮微藻之間的相互作用研究[J].環境科學, 2006,27(2): 274-280.
[14]呂頌輝, 陳翰林, 何智強.氮磷等營養鹽對尖刺擬菱形藻生長的影響[J].生態環境, 2006, 15(4): 697-701.
[15]曲克明, 陳碧鵑, 袁有憲, 等.氮磷營養鹽影響海水浮游硅藻種群組成的初步研究[J].應用生態學報,2000, 11: 445-448.
[16]Piehler M F, Twomey J G, Hall N S, et al.Impacts of inorganic nutrient enrichment on phytoplankton community structure and function in Pamlico Sound,NC, USA[J].Estuarine, Coastal and Shelf Science,2004, 61: 197-209.
[17]黃偉, 朱旭宇, 曾江寧, 等.氮磷比對浙江近岸浮游植物群落結構影響的實驗研究[J].海洋學報, 2012,34(5): 128-136.
[18]錢燕, 張鷹, 劉吉堂.海州灣海域赤潮災害與成因探討[J].海洋湖沼通報, 2008, 3: 191-196.
