擬青霉FLH30胞內(nèi)β-葡萄糖苷酶特性研究
2014-12-16 08:09:10華承偉于江傲常景玲張志宏
食品工業(yè)科技 2014年13期
華承偉,于江傲,李 蘭,常景玲,張志宏
(河南科技學(xué)院生科院,河南新鄉(xiāng)453003)
β-葡萄糖苷酶能夠?qū)⒗w維二糖水解成葡萄糖,從而解除纖維素在酶解過程中產(chǎn)生的纖維二糖對纖維素酶的抑制作用[1],所以,在纖維素的水解中起著非常重要的作用。β-葡萄糖苷酶也是一類具有重要生理功能的水解酶,能催化氧親合糖基轉(zhuǎn)移的水解酶,在一些條件下可催化芳基、氨基、或烷基-β-D-糖苷以及生青糖苷、短鏈低聚糖和二糖中的糖苷鍵,此外,在特定的條件下會發(fā)生水解的逆反應(yīng),生成一些新的糖苷[2]。β-葡萄糖苷酶在食品中不僅可以用于改良果汁風(fēng)味、果酒增香、茶葉增香、生產(chǎn)大豆異黃酮活性苷元、去除苦澀物質(zhì)[3-4],而且,可用于酵母提取物的生產(chǎn)和海藻生物轉(zhuǎn)化生產(chǎn)可發(fā)酵葡萄糖,利用其轉(zhuǎn)移活性應(yīng)用于低聚糖和糖綴合物合成等。此外,在日用化工業(yè)、醫(yī)藥行業(yè)中也有廣泛的用途[5]。
β-葡萄糖苷酶按存在的部位,可分為胞內(nèi)和胞外2種,根據(jù)氨基酸序列相似性,β-葡萄糖苷酶分別隸屬于糖苷水解酶家族1和3[6-7]。家族1中的β-葡萄糖苷酶來自于細(xì)菌、植物和哺乳動物,家族3中的β-葡萄糖苷酶主要來自于真菌、細(xì)菌和植物。幾乎所有的β-葡萄糖苷酶對底物的糖基部分結(jié)構(gòu)的專一性較差,能裂解 C-O、C-S、C-N、C-F等鍵,有些對糖基部分的C4和C2構(gòu)形也不專一,能同時水解β-葡萄糖苷鍵和β-半乳糖苷鍵,甚至對C6位的專一性也不高,能夠水解木糖苷[8]。本實驗對分離得到的耐熱真菌擬青霉FLH30進(jìn)行細(xì)胞破碎,并經(jīng)凝膠層析和離子交換得到的電泳級β-葡萄糖苷酶純酶進(jìn)行性質(zhì)探索,為該酶的基因克隆及利用提供借鑒作用。
1 材料與方法
1.1 材料與儀器
菌株 Paecilomyces sp.FLH30,河南科技學(xué)院分離工程實驗室保存。
發(fā)酵培養(yǎng)基(g/L) 酵母提取物10,蛋白胨10,葡萄糖 20,KH2PO41.0,MgSO4·7H2O 0.3,CaCl20.4,F(xiàn)eSO4·7H2O 0.1,pH,自然;酵母提取物、蛋白胨Oxoid公司(英國);大麥葡聚糖、地衣多糖、昆布多糖、木聚糖、pNP-β-D-葡萄糖苷、pNP-α-D-葡萄糖苷、pNP-β-D-半乳糖苷、ONP-β-D-半乳糖苷、纖維二糖、龍膽二糖和乳糖 Sigma公司(美國);纖維寡糖(G3~G4) 自制;丙烯酰胺、甘氨酸、Tris Biomol公司(美國);Sephacryl-S-100HR Pharmacia公司(美國);DEAE52 Whatman公司(英國);低分子量蛋白質(zhì)標(biāo)準(zhǔn) TaKaRa公司(日本);葡萄糖氧化酶試劑盒 北京北化康泰化學(xué)試劑有限公司;其它試劑均為分析純。
超聲波超微細(xì)胞破碎儀 浙江寧波新芝生物科技股份有限公司;TU-1800PC紫外可見分光光度計北京普析通用儀器設(shè)備有限責(zé)任公司;Power Pac BasicTM型電泳儀 BIO-RAD公司(美國);GL-20B高速冷凍離心機(jī) 上海安亭科學(xué)儀器廠;Kieselgel 60層析板 Merck公司(德國);核酸蛋白層析系統(tǒng)上海滬西儀器廠。
1.2 實驗方法
1.2.1 粗酶液制備 取搖瓶發(fā)酵培養(yǎng)5d的培養(yǎng)物,3000r/min離心 10min,去除上清,菌絲體用適量50mmol/L pH6.0的磷酸緩沖液重懸,冰浴條件下超聲(300W),工作 3s,停 5s,循環(huán) 100 次,破碎液經(jīng)10000r/min 4℃冷凍離心 10min,取上清,即為粗酶液。
1.2.2 酶的純化及分子量檢測 粗酶液經(jīng)Sephacryl-S-100 HR凝膠層析(1×100cm,流速1mL/min),收集有酶活性的部分,經(jīng)超濾濃縮后,再用50mmol/L pH6.0的 NaH2PO4-Na2HPO4緩沖液透析過夜,經(jīng)DEAE52離子交換(1×10cm,上樣及平衡緩沖液同透析緩沖液,含有100mmol/L NaCl的緩沖液洗滌3~5個柱體積,100~500mmol/L NaCl的緩沖液梯度洗脫,流速 3mL/min),1mL/管,分步收集后,SDSPAGE電泳[9]分別檢測酶蛋白純度(分離膠和濃縮膠分別為5%和12.5%),收集有酶活且純度較高的部分,經(jīng)超濾濃縮和4℃冷凍離心后,取上清4℃保存,作為實驗用酶材料。
Sephacryl-S-100HR凝膠過濾法測定分子量以磷酸化酶B(97.4ku)、牛血清蛋白(66.2ku)、兔肌動蛋白(43.0ku)、牛乳酪蛋白(25.0ku)和細(xì)胞色素C(12.4ku)作為標(biāo)準(zhǔn)蛋白。按公式Ve=-b'lgMw+c',通過lgMw對Ve做圖,從曲線上求出分子量。
1.2.3 蛋白含量測定 參照Lowry等的方法[10],以牛血清白蛋白作為標(biāo)準(zhǔn)蛋白。
1.2.4 酶活測定 于試管中加入75μL用蒸餾水配制的 15mmol/LpNPG底物,然后加入 150μL 75mmol/L,pH6.0磷酸檸檬酸緩沖液,混勻,使底物終濃度為5mmol/L,緩沖液濃度為50mmol/L,50℃預(yù)熱3min,然后加入25μL適當(dāng)稀釋的酶液,反應(yīng)10min后,加入750μL飽和四硼酸鈉溶液終止反應(yīng),冷卻后測定A405,以pNP作標(biāo)準(zhǔn)計算酶活力。酶活力的單位定義為:在上述條件下,每分鐘生成1μmol pNP所需要的酶量。
1.2.5 最適反應(yīng)pH和pH穩(wěn)定性 45℃下,將緩沖液分別換成不同pH及不同體系(Na2HPO4-Citric,2.5 ~ 6.5;NaH2PO4-Na2HPO4,6.0 ~ 8.0;CHES,8.0~11.0;Na2HPO4~NaOH,11.0~12.0;KCl~NaOH,12.0~13.0)的緩沖液加入到底物中,然后按照標(biāo)準(zhǔn)方法測定酶活力,以酶活力最高點作為100%,確定最適pH;用上述不同的pH緩沖液分別稀釋純酶液,將稀釋好的酶液置于25℃水浴鍋中分別處理30min,迅速將樣品置于冰水中冷卻30min,然后測定殘余酶活力,以未經(jīng)處理的酶液作為對照,計算殘余酶活力的百分比,考查pH穩(wěn)定性。結(jié)果為三次實驗數(shù)據(jù)的平均值。
1.2.6 最適溫度和溫度穩(wěn)定性 將重組酶適當(dāng)稀釋于50mmol/L pH6.0的NaH2PO4-Na2HPO4緩沖液中,然后分別在30~70℃不同溫度下按照上述方法測定酶活力。溫度穩(wěn)定性測定:將酶液用50mmol/L pH6.0的NaH2PO4-Na2HPO4緩沖液稀釋后分別在不同的溫度下處理30min,然后置于冰水浴中冷卻30min,最后按標(biāo)準(zhǔn)的方法測定殘余酶活力,以未經(jīng)處理的酶液酶活力為100%對照。結(jié)果為三次實驗數(shù)據(jù)的平均值。
1.2.7 金屬離子及化合物對酶活力的影響 在緩沖液為50mmol/L,pH6.0的NaH2PO4-Na2HPO4緩沖液的酶溶液中分別加入不同的金屬離子,使其終濃度為4mmol/L,將酶液在25℃下處理 30min,然后在50℃條件下測定剩余酶活力,以沒有添加金屬離子的酶液作為對照。結(jié)果為三次實驗數(shù)據(jù)的平均值。
1.2.8 底物特異性 分別以pNP-糖苷、二糖、低聚糖和聚糖為反應(yīng)底物測定酶活力。pNP-糖苷類底物以水解釋放出 pNP的速率來計算酶活力,采用50mmol/L,pH6.0 NaH2PO4-Na2HPO4緩沖液配制5mmol/L濃度的底物,然后在50℃下反應(yīng)10min,最后測定405nm的吸光度值;低聚糖用50mmol/L,pH6.0 NaH2PO4-Na2HPO4緩沖液配制10mg/mL濃度的底物,然后在50℃下反應(yīng)10min,最后通過檢測水解過程中釋放出葡萄糖的速率來計算酶活力。葡萄糖的濃度采用葡萄糖氧化酶試劑盒測定;聚糖類底物用50mmol/L,pH6.0 NaH2PO4-Na2HPO4緩沖液配制成1%(w/v)濃度,然后在50℃下反應(yīng)10min,最后采用DNS法測定水解過程中釋放的還原糖量。酶活力單位定義為在上述條件下每分鐘反應(yīng)生成1μmol pNP、葡萄糖或還原糖所需要的酶量。然而,對于低聚糖中的纖維二糖和龍膽二糖,一個酶活力單位為生成2μmol的葡萄糖的量(水解一個糖苷鍵釋放出兩個葡萄糖)。所有的實驗均經(jīng)過三次重復(fù)。
1.2.9 聚糖和纖維多糖水解TLC分析 以50mmol/L,pH6.0的NaH2PO4-Na2HPO4緩沖液配制10%的纖維多糖和1%的大麥葡聚糖、地衣多糖和昆布多糖,添加10U/mL的重組酶,50℃保溫水解,定時取樣進(jìn)行薄層層析(TLC,Kieselgel60)分析,展層系統(tǒng)為正丁醇/乙酸/水系統(tǒng)(2∶1∶1,v/v/v),將水解的樣品點樣后展開兩次,吹干后用硫酸∶甲醇(5∶95,v/v)溶液浸濕,最后在130℃烘箱中烘烤顯色。
2 結(jié)果與討論
2.1 糖苷酶純化及分子量測定
經(jīng)Sephacryl-S-100HR凝膠層析和DEAE52離子交換,收集純度較高的部分再經(jīng)SDS-PAGE檢測(上樣10μg),達(dá)到電泳級純酶(圖1)。
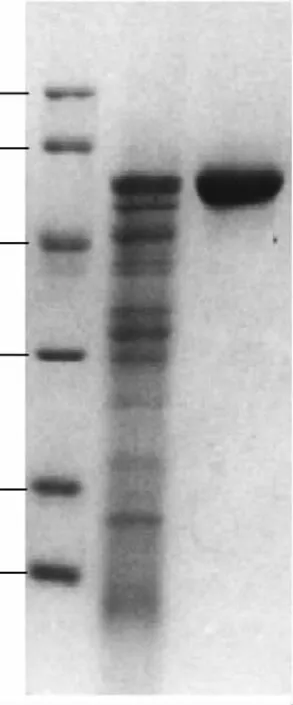
圖1 純化β-葡萄糖苷酶SDS-PAGE電泳檢測列Fig.1 Analysis of purified β-glucosidase by SDS-PAGE
微生物胞內(nèi)β-葡萄糖苷酶一般屬于1家族糖苷水解酶,分子量較小,而胞外糖苷酶大多屬于3家族糖苷水解酶,分子量較大,且由于β-葡萄糖苷酶的糖基化作用,分子量一般在100~250ku之間[11]。SDS-PAGE電泳結(jié)果顯示為單一條帶蛋白,分子量在44.3~66.4ku之間,為確定在活性狀態(tài)下該酶的分子量,凝膠過濾法測定了其分子量,根據(jù)凝膠過濾標(biāo)準(zhǔn)蛋白分子量曲線(圖2),測定活性狀態(tài)下的β-葡萄糖苷酶分子量為57.2ku,對比SDS-PAGE電泳結(jié)果,該酶應(yīng)為1家族糖苷水解酶且為單亞基蛋白。
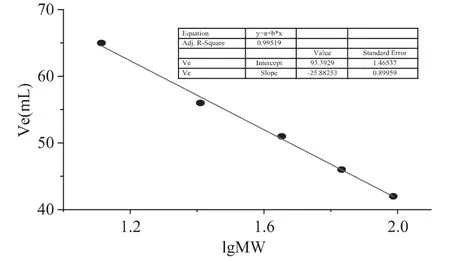
圖2 凝膠過濾測β-葡萄糖苷酶標(biāo)準(zhǔn)曲線Fig.2 The curve of gel filtration chromatography of standard protein
2.2 最適反應(yīng)pH和pH穩(wěn)定性
該葡萄糖苷酶的最適pH約為6.0,pH對其活性影響較大,pH低于5.0,嚴(yán)重抑制酶活性,在pH4.5時,只有最高酶活的12%,當(dāng)pH高于6.5時,酶活性也急劇下降(圖3a);該酶在pH5.0~12.0(圖3b)范圍內(nèi)比較穩(wěn)定,處理30min后,殘余酶活力仍保持在80%以上。
2.3 最適溫度和溫度穩(wěn)定性
在不同溫度下測得酶活性數(shù)據(jù)顯示該酶最適反應(yīng)溫度60℃左右,超過65℃,酶活性快速下降(圖4a);該酶在70℃以下處理30min非常穩(wěn)定,超過70℃后,酶活性快速喪失,85℃處理30min,只有原酶活性的12.5%。
2.4 金屬離子及化合物對酶活力的影響
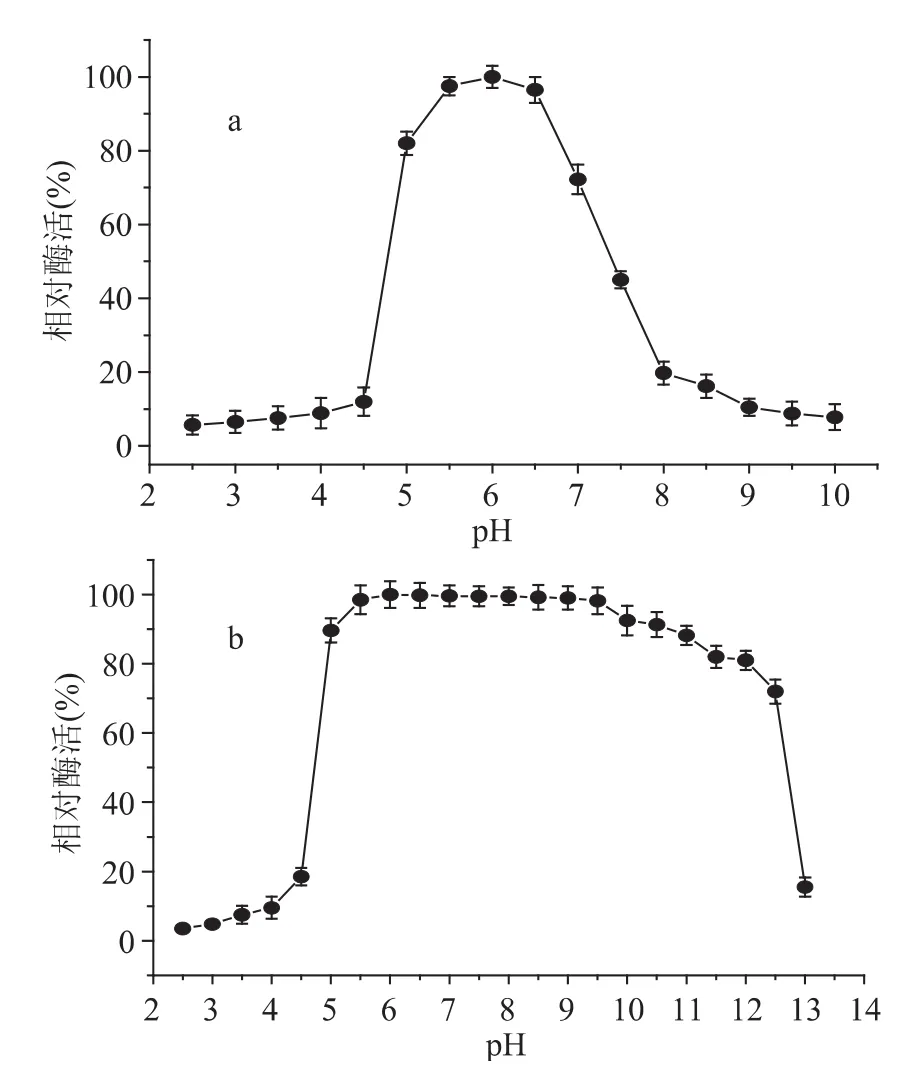
圖3 β-葡萄糖苷酶最適pH(a)和pH穩(wěn)定性(b)Fig.3 The optimal pH(a)and pH stability(b)of β-glucosidase
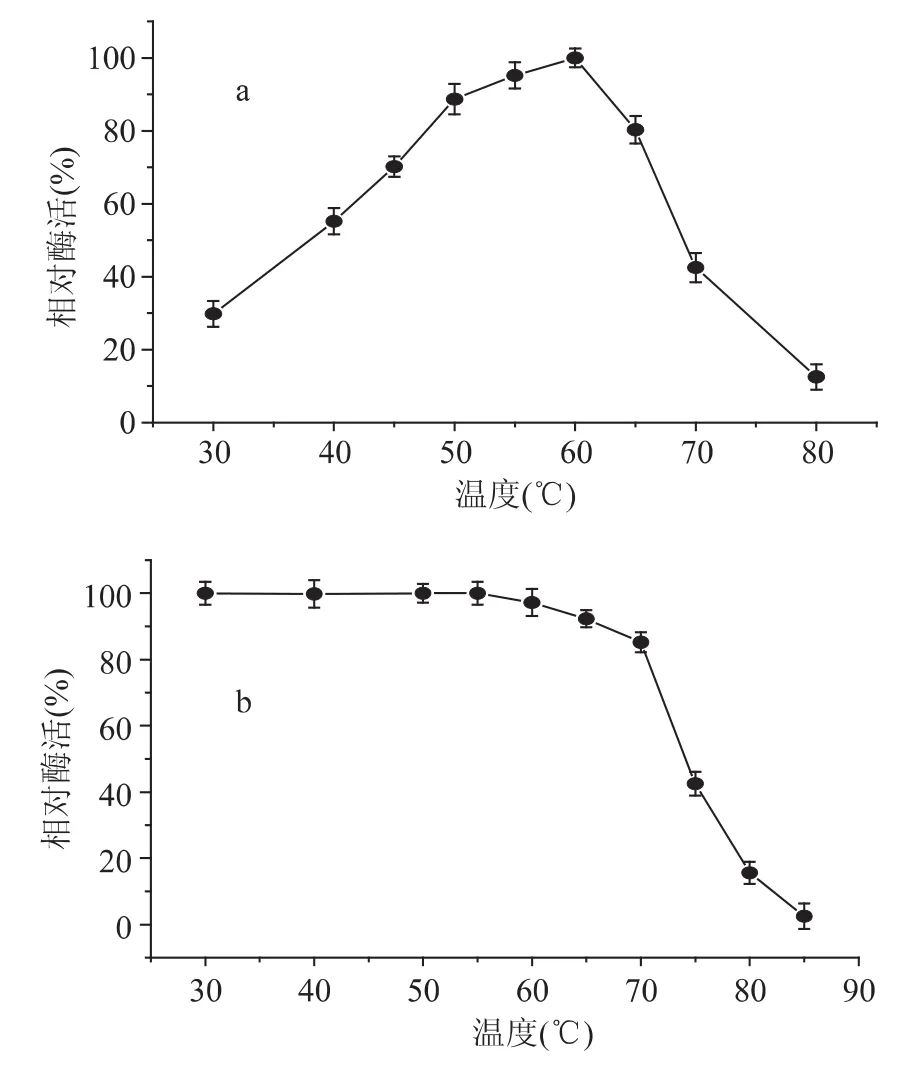
圖4 β-葡萄糖苷酶最適溫度(a)和溫度穩(wěn)定性(b)Fig.4 The optimal temperature(a)and temperature stability(b)of β-glucosidase
部分金屬離子及化合物處理數(shù)據(jù)顯示(表1),Ag+、Hg2+和Fe3+強(qiáng)烈抑制酶活性,Zn2+有較強(qiáng)的抑制作用。其它金屬離子對酶活沒有顯著的影響。其中,EDTA對酶活沒有顯著影響,表明金屬離子不是酶的必需輔助因子,SDS,DTT和β-巰基乙醇對酶活也沒有顯著影響,說明二硫鍵不是維持酶正常空間構(gòu)象的必需作用力。
2.5 底物特異性
該胞內(nèi)葡萄糖苷酶具有較寬的底物作用范圍(表2),對 pNP-(-D-半乳糖苷、ONP-β-D-半乳糖苷和纖維二糖有較高的比活力。此外,對部分聚糖也有水解能力,可以水解地衣多糖、昆布多糖和大麥葡聚糖,但不能水解木聚糖和羧甲基纖維素。該酶作用底物及糖苷鍵型比較廣泛,預(yù)示該酶具有較廣的用途。

表2 葡萄糖苷酶底物特異性Table 2 Analysis of substrate specificity of β-glucosidase
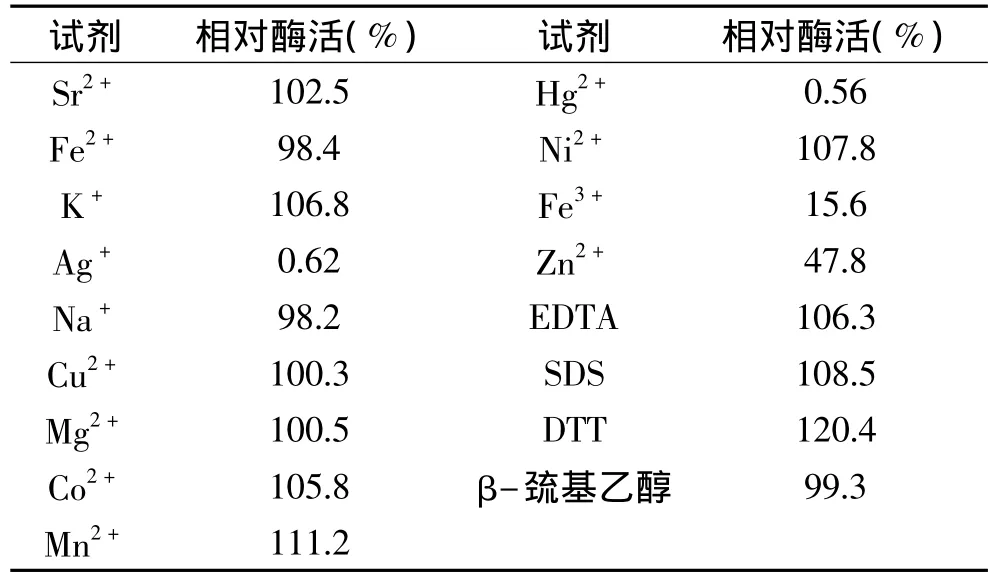
表1 金屬離子及其它化合物對β-葡萄糖苷酶的影響Table 1 The metal ions and other chemicals influence on β-glucosidase
2.6 聚糖和纖維多糖水解TLC分析
大麥葡聚糖、昆布多糖、纖維三糖和纖維四糖的水解實驗表明,水解大麥葡聚糖、昆布多糖沒有見到低聚寡糖的生成,產(chǎn)物主要以單糖為主(圖5),可能以外切的方式作用于這些聚糖;此酶可以完全水解纖維三糖和四糖,在4h后基本完全降解為單糖,同時,水解過程中伴隨著轉(zhuǎn)糖苷作用,這種轉(zhuǎn)糖苷作用在最初的2h內(nèi)較為明顯,1~2h之間在二糖和三糖之間有一新糖生成,可能為龍膽二糖,但隨水解時間增加,逐漸水解為單糖(圖6)。
3 結(jié)論
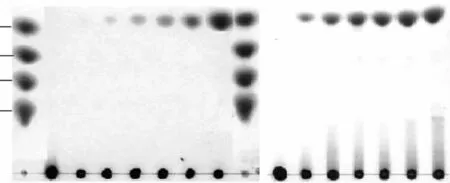
圖5 TLC分析β-葡萄糖苷酶水解大麥葡聚糖和昆布多糖Fig.5 Analysis of β-glucosidase hydrolysis of barley glucan and laminarin by TLC
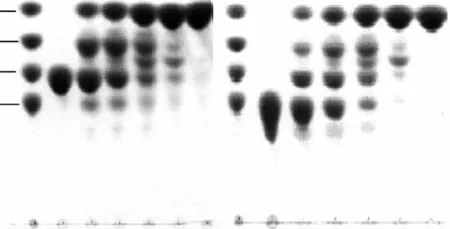
圖6 TLC分析β-葡萄糖苷酶水解纖維三糖和四糖Fig.6 Analysis of β-glucosidase hydrolysis of cellotriose and cellotraose by TLC
依據(jù)水解底物特異性的不同可將β-葡萄糖苷酶劃分成β-葡萄糖苷酶、纖維二糖酶和具有較寬水解特異性,能夠水解多種不同底物的β-葡萄糖苷酶三類。本擬青霉FLH30為耐熱真菌[12],胞內(nèi)β-葡萄糖苷酶作用底物廣泛,應(yīng)屬于第三類β-葡萄糖苷酶。1家族β-葡萄糖苷酶最適溫度一般在40~50℃之間,該β-葡萄糖苷酶最適溫度60℃,在此溫度下處理30min,酶活損失小于10%,70℃處理30min,酶活損失也小于20%。不同來源β-葡萄糖苷酶pI一般都在酸性范圍內(nèi)(3.5~5.5),并且差別不大,最適pH也多在酸性范圍內(nèi)且最適作用pH范圍較窄,本β-葡萄糖苷酶最適pH6.0,在pH4.5以下,酶活性急劇下降,但該酶非常耐堿,在pH12.0處理30min后,殘余酶活力仍保持在70%以上,預(yù)示該酶在食品、醫(yī)藥、紡織、能源等方面具有較大潛在應(yīng)用價值。
[1]Hong J,Tamaki H,Kumagai H.Cloning and functional expression of thermostable β-glucosidase gene from Thermoascus aurantiacus[J].Applied Microbiology and Biotechnology,2007,73:1331-1339.
[2]Mohamed G,Issam S.Fungus β-glycosidases:immobilization and use in alkyl β-glycoside synthesis[J].Journal of Molecular Catalysis B:Enzymatic,2004,29:89-94.
[3]JeyaM,JooAR,LeeKM,etal.Characterization of β-glucosidase from a strain of Penicillium purpurogenum KJS506[J].Applied Microbiology and Biotechnology,2010,86:1473-1484.
[4]Zhou C,Qian L,Ma H,et al.Enhancement of amygdalin activated with β-d-glucosidase on HepG2 cells proliferation and apoptosis[J].Carbohydrate Polymers,2012,90:516-523.
[5]潘利華,羅建平.β-葡萄糖苷酶的研究及應(yīng)用進(jìn)展[J].食品科學(xué),2006,27(12):803-806.
[6]Henrissat B,Callebaut I,F(xiàn)abrega S,et al.Conserved catalytic machinery and the prediction of a common fold for several families of glycosyl hydrolases[J].Proceedings of the National Academy of Sciences of the United States of America,1996,93(17):9302.
[7]Ketudat CJR,Esen A.β -Glucosidases[J].Cellular and Molecular Life Sciences,2010,67(20):3389-3405.
[8]Driskill LE,Bauer MW,Kelly RM.Synergistic interactions among beta-laminarinase,beta-1,4-glucanase,and betaglucosidase from the hyperthermophilic archaeon Pyrococcus furiosus during hydrolysis of beta-1,4-,beta-1,3-,and mixedlinked polysaccharides[J].Biotechnol Bioeng,1999,66(1):51-60.
[9]Laemmli UK.Cleavage of structural proteins during assembly of head of bacteriophage-T4[J].Nature,1970,227:680-685.
[10]Lowry O H,Rosebrough N J,F(xiàn)arr A L,et al.Protein measurement with the folin phenol reagent[J].Journal of Biological Chemistry,1951,193:265-275.
[11]Nakkharat P,Haltrich D.Purification and characterization of an intracellular enzyme with β -glucosidase and β -galactosidase ac tivity from the thermophilic fungus Talaromyces thermophilus CBS236.58[J].Journal of Biotechnology,2006,123(3):304-313.
[12]華承偉,于江傲,謝鳳珍,等.耐熱 β-1,3(4)-葡聚糖酶源真菌篩選、鑒定及產(chǎn)酶條件優(yōu)化[J].食品工業(yè)科技,2012,33(13):131-134.
