NaCl脅迫對結球甘藍幼苗生長及體內離子分布的影響
2015-01-01 02:14:48顧閩峰王乃頂王偉義費月躍彭亞明王神云李建斌
江蘇農業學報 2015年3期
顧閩峰, 王乃頂, 王偉義, 費月躍, 彭亞明, 王 軍, 王神云, 李建斌
(1.鹽城市新洋農業試驗站,江蘇 鹽城 224049;2.江蘇省農業科學院蔬菜研究所,江蘇 南京 210014)
中國的鹽堿地面積約9.91×107hm2,占世界鹽堿地面積的26.3%[1],其中江蘇省鹽堿地面積大,沿海灘涂資源豐富,是中國最大面積的沿海灘涂[2-3]。江蘇省沿海灘涂鹽堿地的開發利用是沿海開發戰略的重要內容,在鹽堿地上種植種類豐富的植物資源,生物治鹽是改良灘涂鹽堿地的主要途徑之一[4]。因此,研究植物的耐鹽特性,培育耐鹽植物,利用耐鹽植物開發灘涂鹽堿地資源,是發展現代鹽土農業的重要選擇,也是加快推進江蘇省沿海灘涂鹽堿地開發利用的關鍵。
結球甘藍(Brassica oleracea var.capitata)又稱洋白菜、卷心菜,是十字花科蕓薹屬植物,是中國重要的蔬菜作物之一[5],有較豐富的鉀元素、維生素C等營養物質,具有和胃、健脾、止痛、防衰老等功效[6-9]。目前國內外關于結球甘藍的研究主要集中在栽培技術、成分分析和抗蟲病害等方面,有關其在鹽脅迫下的幼苗生長、生理特性、離子分布運輸的報道較少[10-11]。本試驗以結球甘藍為材料,研究不同NaCl濃度處理對結球甘藍幼苗的生長和體內離子吸收分配的影響,以期為篩選耐鹽結球甘藍品種提供依據。
1 材料與方法
1.1 材料的培養和處理
供試結球甘藍種子采自南京農業大學江蘇大豐“863”試驗基地,試驗于2013年12月初將經過低溫沙藏并萌動露白的種子播種到裝有蛭石的營養缽中。每盆播種1粒,置于溫室中育苗。待幼苗生長至5片或6片葉齡,選取長勢一致的幼苗定植到塑料容器中進行水培。當長至第8片葉子時,施以1/2 Hoagland營養液,鹽脅迫前,對幼苗進行為期18 d的預培養,以使其適應水培條件,正常生長。試驗設7 個 NaCl濃度:0 mmol/L(對照)、17 mmol/L、34 mmol/L、51 mmol/L、68 mmol/L、85 mmol/L和 102 mmol/L。每個濃度水平重復3次,每個重復12株,營養液每2 d更換一次。為避免對幼苗造成滲透休克,采取每2 d增加NaCl的方式遞增NaCl濃度。試驗期間,溫室內晝夜溫度保持在22~28℃,相對濕度70% ~80%,經過21 d的NaCl脅迫處理,收集植物樣品用于分析。
1.2 鮮質量和干質量的測定
試驗結束時,將結球甘藍幼苗從塑料盆中取出,每個處理隨機選取甘藍幼苗12株,用去離子水沖去砂粒,然后迅速用吸水紙吸干,采集結球甘藍幼苗的根、莖、葉,分別稱其鮮質量,隨后放在105℃下殺青30 min,放入烘箱進行烘干。溫度為65~70℃,經檢查樣品烘干后,關掉烘箱,待溫度降到室溫后,分別測得各組分的生物量,并計算冠生物量(莖生物量+葉生物量)、全株總生物量(根生物量+冠生物量)、相對生物量(某處理的平均總生物量/對照的平均總生物量×100%)以及根冠比值(根生物量/冠生物量)。
1.3 無機離子含量的測定
在電子天平(精確至0.000 1 g)上用牛角勺稱取待測樣品0.1 g左右,將樣品小心轉移至干凈且干燥的消煮管內,放在管架上。向消煮管內加入2 ml優級純的濃HNO3,搖勻后,擰緊蓋子,進行消煮。消煮完畢后,取出消煮管并放在管架上,冷卻30 min。消煮液轉移至50 ml洗凈的容量瓶中,用高純水清洗消煮管2次(少量多次)并轉移入容量瓶中,再用高純水洗小漏斗后再定容后搖勻。靜置1 h以上后轉移上清液至15 ml離心管內保存(可以先用定容液潤洗離心管)。經微波消煮過的樣品直接用電感耦合等離子體發射光譜儀ICP-OES(Inductively coupled plasma optical emission spectrometer,Optima 2100DV,Pekin-Elmer,USA)測定甘藍幼苗根、莖、葉中 K+、Na+、Ca2+和 Mg2+含量,每次測定取3個重復,并計算各組中的 K+/Na+、Ca2+/Na+、和Mg2+/Na+值及無機離子累積量,無機離子累積量=離子濃度×質量。
1.4 數據處理
利用Microsoft Excel軟件、SPSS 19軟件進行試驗數據的統計,采用Duncan’s新復極差法進行顯著性分析。
2 結果與分析
2.1 NaCl脅迫對甘藍幼苗生物量累積與分配的影響
經不同濃度NaCl脅迫后,根部生物量在17 mmol/L和34 mmol/L處理下均略高于對照的生物量,51 mmol/L、68 mmol/L、85 mmol/L和 102 mmol/L處理下的生物量均低于對照。所有NaCl脅迫處理的結球甘藍幼苗的莖、葉以及全株總生物量均不同程度的低于對照(表1),且均隨著NaCl脅迫濃度的升高結球甘藍幼苗的莖、葉以及全株總生物量呈下降趨勢,說明NaCl脅迫抑制了植株生物量的積累。植株的相對生物量隨著NaCl濃度的升高而下降。7個NaCl脅迫處理間的根冠比無顯著差異。
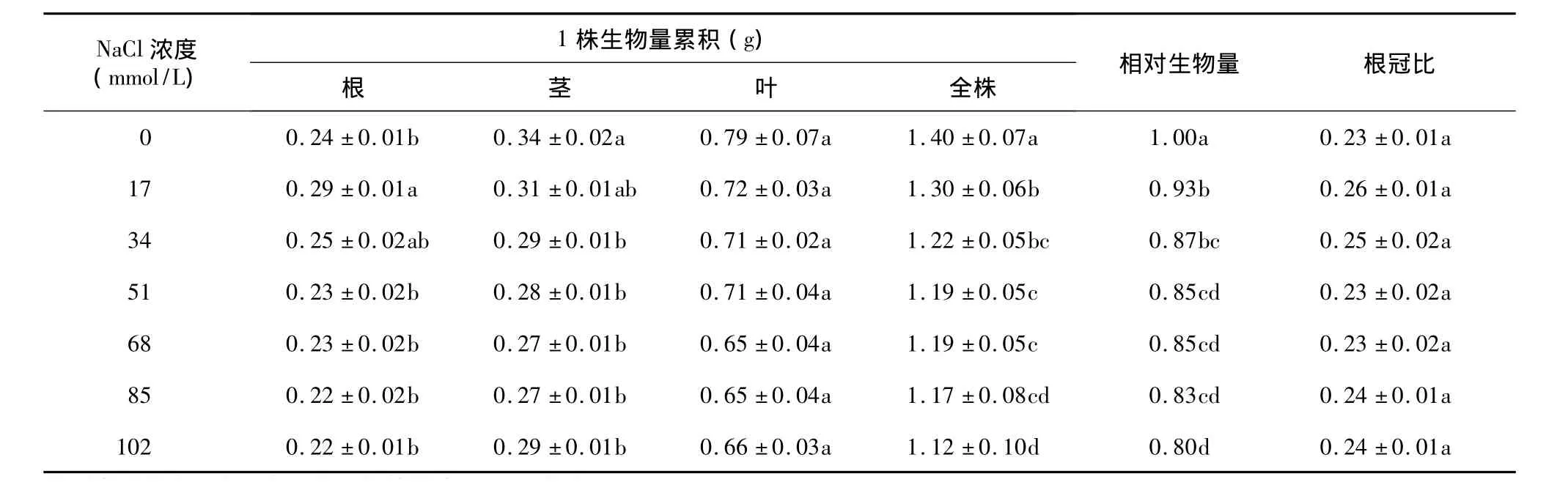
表1 不同濃度NaCl對結球甘藍幼苗生物量累積和根冠比的影響Table 1 Effects of NaCl concentrations on biomass accumulation and root and shoot ratio of Brassica oleracea var.capitata seedlings
2.2 NaCl脅迫對結球甘藍幼苗不同組織離子含量的影響
由圖1A可以看出,隨著NaCl濃度的增加,結球甘藍幼苗體內各器官中的Na+含量均呈逐漸升高的趨勢;不同NaCl濃度處理下,甘藍幼苗各器官中的Na+含量分布均為:莖>葉>根。與對照相比,在17 mmol/L處理下,根、莖、葉中的Na+含量均顯著增加,分別是對照植株的1.61、1.66和2.10倍。在102 mmol/L處理下,各器官中的Na+含量達到最大值,根、莖、葉中的Na+含量分別是對照植株的2.40、2.71和4.18倍。上述結果表明,當結球甘藍幼苗受到NaCl脅迫時,Na+大量進入植物體中,并將大部分的Na+滯留在莖部,根部Na+積累少,從而減輕鹽離子對根部的毒害。
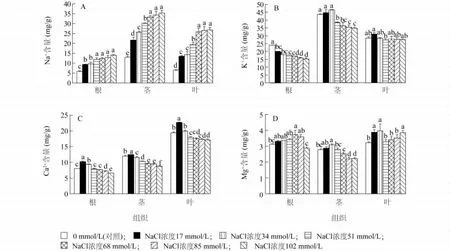
圖1 不同濃度NaCl對甘藍幼苗各組織K+、Na+、Ca2+和Mg2+含量的影響Fig.1 Effects of NaCl concentrations on K+,Na+,Ca2+and Mg2+contents in different tissues of B.oleracea var.capitata seedlings
由圖1B可以看出,隨著NaCl濃度的升高,結球甘藍幼苗根中的K+含量呈逐漸下降的趨勢,而K+含量在莖和葉中則呈先上升后下降的趨勢。在17 mmol/L和34 mmol/L NaCl處理下,幼苗莖中K+含量均高于對照,而隨著 NaCl濃度的增加,從51 mmol/L處理開始,莖中K+含量均顯著低于對照。葉中K+含量只有17 mmol/L處理下高于對照,其余各處理下均低于于對照。不同NaCl濃度處理下,結球甘藍幼苗各器官中的K+含量分布均為:莖>葉>根。
不同NaCl濃度處理下,結球甘藍各組織Ca2+含量由大到小依次為:葉、莖、根(圖1C)。隨著NaCl濃度的升高,Ca2+含量在各組織中都是呈先升高后下降的趨勢,在17 mmol/L處理下,根、莖、葉的Ca2+含量分別是對照的0.05、0.03和0.15倍;在34 mmol/L處理下,根和葉的Ca2+含量也高于對照;隨著鹽濃度的增加,從51 mmol/L處理開始,結球甘藍幼苗各器官中的Ca2+含量均低于對照。
由圖1D可以看出,在不同濃度NaCl脅迫下,結球甘藍幼苗各組織中Mg2+含量最高的是葉,其次是根和莖。隨著NaCl濃度的升高,根和莖中Mg2+含量均先升高然后下降,在17~85 mmol/L NaCl處理下,根部Mg2+含量均高于對照,在17~51 mmol/L NaCl處理下,結球甘藍莖部Mg2+含量均高于對照。在17~34 mmol/L和102 mmol/L NaCl處理下,葉部Mg2+含量均顯著高于對照,而51~85 mmol/L NaCl處理下,葉部Mg2+含量與對照差異不顯著。
2.3 NaCl脅迫對結球甘藍幼苗K+/Na+、Ca2+/Na+和Mg2+/Na+值的影響
由表2可以看出,不同濃度NaCl處理下,結球甘藍幼苗各器官中的K+/Na+、Ca2+/Na+和Mg2+/Na+值均低于對照,且隨著NaCl濃度的升高呈逐漸下降的趨勢。在17 mmol/L處理下,結球甘藍幼苗根、莖、葉的K+/Na+值,比對照分別下降了48%、38%和48%,Ca2+/Na+值分別下降了21%、37%和44%,Mg2+/Na+值分別下降了34%、38%和43%;在102 mmol/L處理下,K+/Na+、Ca2+/Na+和Mg2+/Na+值的降幅最大。在不同鹽濃度處理下,甘藍葉中Ca2+/Na+值高于根中相應比值,根中Ca2+/Na+值高于莖;Mg2+/Na+值在根中最高,其次是葉和莖。
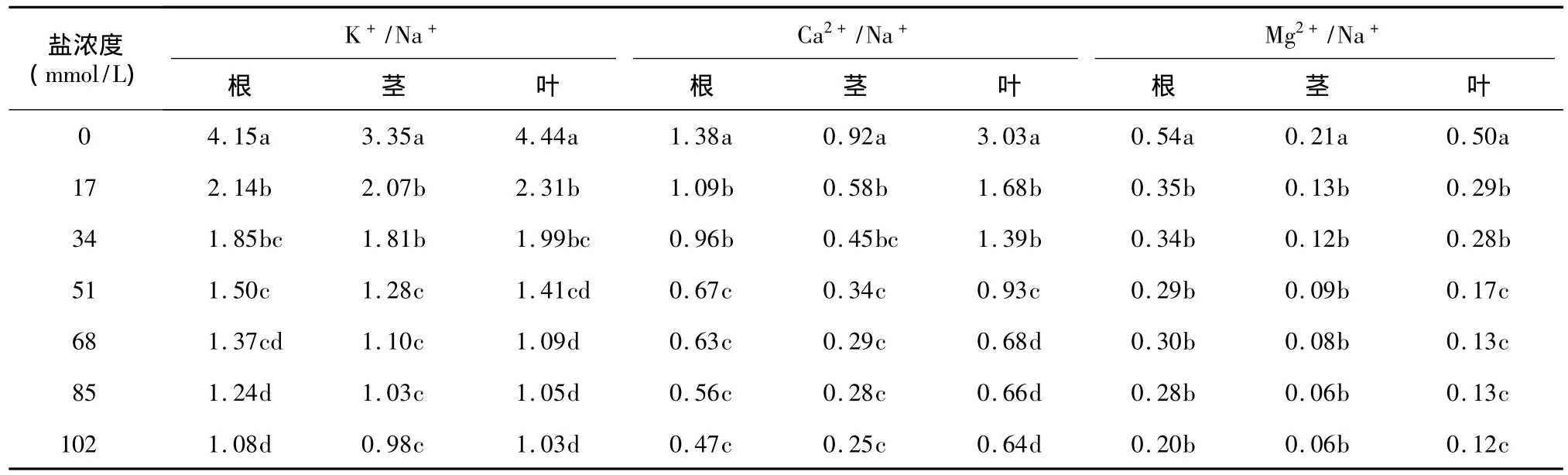
表2 NaCl脅迫對結球甘藍幼苗K+/Na+、Ca2+/Na+和Mg2+/Na+值的影響Table 2 Effects of NaCl concentrations on K+/Na+,Ca2+/Na+and Mg2+/Na+of B.oleracea var.capitata seedlings
2.4 NaCl脅迫對結球甘藍幼苗 K+、Na+、Ca2+和Mg2+離子積累的影響
由圖2A可以看出,隨著NaCl濃度的升高,結球甘藍幼苗Na+累積量呈現逐漸升高的趨勢。17 mmol/L、34 mmol/L、51 mmol/L和 68 mmol/L處理間,甘藍幼苗對Na+的吸收分配存在顯著性差異,4個處理下的整個植株的Na+累積量分別是對照的1.76、1.96、2.43 和 2.82 倍,而 68 mmol/L、85 mmol/L和102 mmol/L處理間差異性不顯著。各處理莖中Na+含量在整個植株中所占的比例均最大,分別占52%、49%、52%、49%、47%、47%和46%,而葉占25%、30%、29%、32%、36%、36%和35%,根占23%、21%、20%、19%、17%、17%和18%。
由圖2B可以看出,不同NaCl濃度處理下,結球甘藍K+累積量均顯著低于對照,分別是對照的74%、84%、74%、72%、68%和65%,且除17 mmol/L外,隨著NaCl濃度的升高,結球甘藍中的K+累積量逐漸下降。在不同NaCl濃度處理下,莖中的K+含量在整個植株中所占比例最大,其次是葉、根,其中34 mmol/L處理下,莖中K+含量所占比例最大。
由圖2C可以看出,在不同NaCl濃度處理下,結球甘藍幼苗Ca2+累積量均低于對照。且隨著NaCl濃度的升高,在34 mmol/L至102 mmol/L處理下,幼苗體中的Ca2+累積量呈下降的趨勢。17 mmol/L至102 mmol/L處理下,結球甘藍體中Ca2+累積量分別是對照的86%、91%、78%、76%、72%和66%。
由圖2D可以看出,對照Mg2+均高于NaCl處理,NaCl濃度最高時,Mg2+在植株中的累積量最低,是對照的79%。在51~102 mmol/L處理間,結球甘藍幼苗的Mg2+累積量無顯著差異。幼苗莖中Mg2+含量顯著低于根和葉。
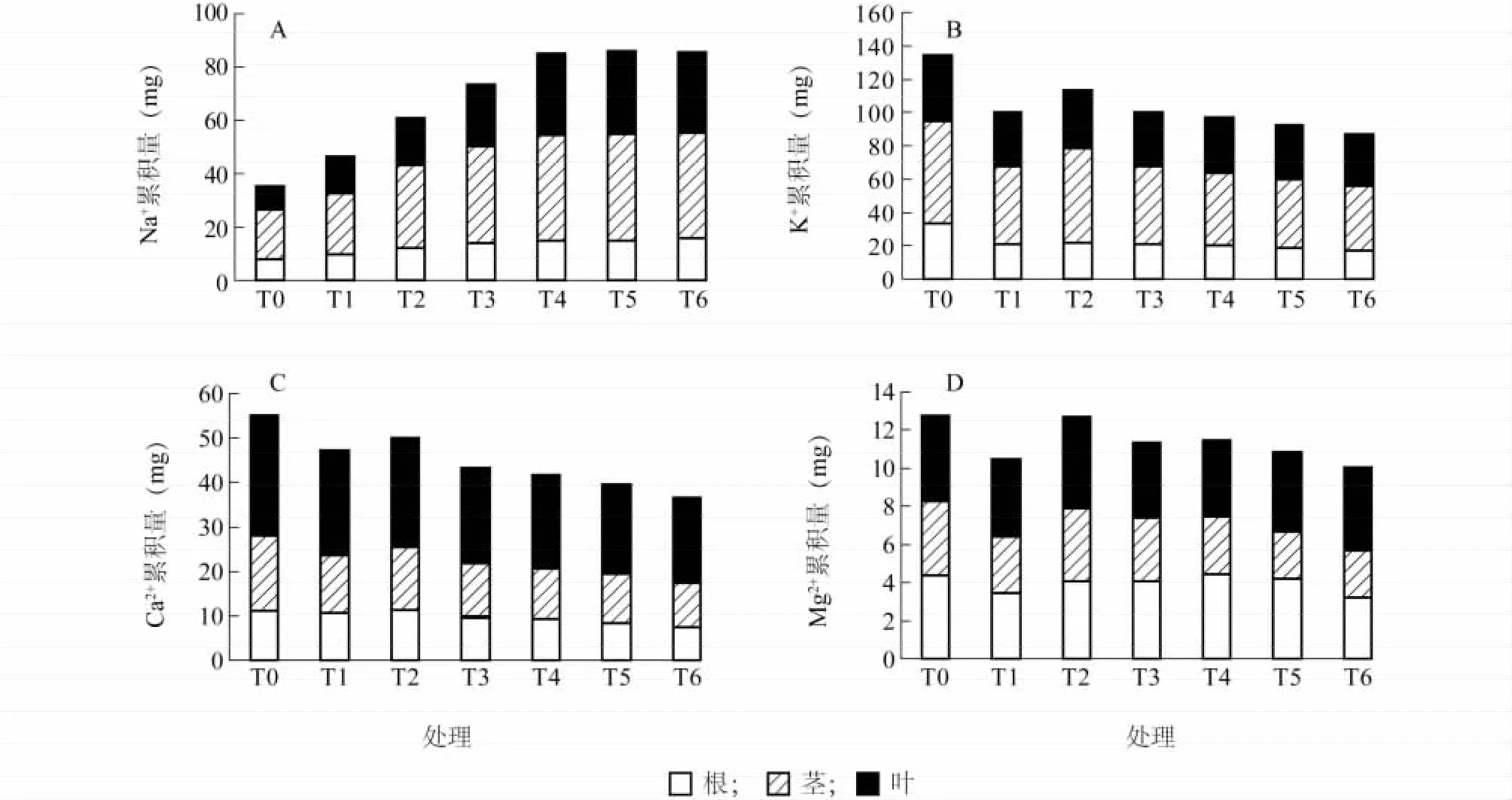
圖2 NaCl脅迫對單株結球甘藍幼苗K+、Na+、Ca2+和Mg2+積累的影響Fig.2 Effects of NaCl stress on K+,Na+,Ca2+and Mg2+accumulations in B.oleracea var.capitata seedlings
3 討論
鹽分通過多種脅迫方式對植物產生傷害,如滲透脅迫中土壤水勢的降低會導致細胞脫水,造成植物體的生理性缺水;離子脅迫會造成對植物細胞膜的損傷,喪失對離子選擇性吸收的功能,致使細胞內離子穩態失衡,影響植物生長發育[12-14],此外鹽脅迫還可能引發氧化脅迫致使植物體內積累過多的活性氧,這些活性氧自由基會啟動膜脂過氧化進程,從而產生氧化脅迫,改變細胞膜通透性,最終引起植物細胞死亡[15]。鹽脅迫對植物生長發育最普遍和最顯著的效應就是抑制植物的生長,降低植物的生物量[16-18]。本研究中隨著 NaCl濃度的升高,根、莖、葉和整株的生物量呈下降的趨勢。植物根冠的大小和功能在正常的條件下是保持平衡的,當植物受到某種資源限制時就會調節生物量分配來適應這種環境的變化,根冠比值是植物在逆境下生物量分配的一種策略,也是植物各器官對鹽脅迫敏感程度的一種表達方式[19-20]。本研究結果顯示,不同處理下甘藍幼苗根冠比間差異不顯著。說明在鹽脅迫下甘藍根系生長的敏感性要低于地上部分。
土壤中鹽分過多就會造成植物的滲透脅迫,并影響營養離子的平衡,導致植物的生理過程發生改變,進而影響到植物的新陳代謝,限制植物生長和發育[21-22]。本研究結果表明:在一定NaCl脅迫下,甘藍幼苗各器官中的Na+含量不斷增加,且不同組織中的增加幅度不一致,這與譚會娟等[23]研究結果一致。隨著NaCl脅迫濃度增加,結球甘藍幼苗各器官中的Na+含量分布均為:莖>葉>根,這說明莖具有較強的貯鹽能力,這可以減少Na+在葉片中的積累,對避免細胞受到離子脅迫和滲透脅迫,維持細胞正常的生理功能起到重要作用。K+是植物在生長發育中必須的大量元素和滲透調節物質,涉及許多生理過程,包括酶活性調節、蛋白質合成及滲透調節等,而且是唯一一種植物所必需的以相對高濃度存在的陽離子[24]。在本研究中,隨著NaCl脅迫濃度的增加,根部K+含量呈逐漸下降,莖和葉中K+含量則先上升后下降,且結球甘藍幼苗各器官中的K+含量分布均為:莖>葉>根,這可能是因為Na+和K+離子半徑和水合能相似,Na+對K+的吸收往往呈現出明顯的競爭性抑制作用[25],所以Na+往往利用K+的途徑進入植物體中,從而使Na+和K+之間相互競爭,造成結球甘藍根中K+含量下降,葉和莖中K+含量先升后降。Ca2+對于保持植物細胞膜結構完整性和功能穩定性具有重要的作用[26]。本研究發現,在低濃度NaCl處理下,各個器官中Ca2+含量弱微增加,而后隨著鹽濃度的升高開始下降,且各個器官中的下降趨勢不一致(圖1C)。Mg2+在作物耐鹽性中的報道較少,鎂也是植物生長必不可少的營養元素[27],本試驗結果表明,NaCl脅迫后,葉片中的Mg2+含量高于根和莖,從而有利于增強植株對鹽脅迫的抵御能力。
K+、Ca2+和Mg2+在植物的生長過程中具有重要的生理功能,Na+在植物氣孔調節、C4和CAM植物磷酸烯醇式丙酮酸的催化再生、鹽生植物的滲透調節等生理活動中發揮著重要的作用。這些離子只有在保持相對平衡的狀態下才能發揮正常的生理作用,當它們之間的平衡被打破,將對生理作用產生不良的影響[28]。本研究結果表明,結球甘藍幼苗各器官中的 K+/Na+、Ca2+/Na+和 Mg2+/Na+值均低于對照,且隨著NaCl濃度的升高呈逐漸下降的趨勢,主要是由于結球甘藍幼苗積累的Na+含量不斷增加,而其他離子含量有所減少造成的。
植物的不同器官具有積累不同離子的作用,在NaCl脅迫下,莖部對Na+的累積最高,其次是K+,而葉中則主要積累Ca2+和Mg2+,這與王冉等對黑籽南瓜幼苗受NaCl脅迫后的離子分布大體一致[29],說明結球甘藍幼苗主要將Na+積累在莖部,從而減輕Na+對根部和葉部的鹽害,而葉片中積累較多的Ca2+和Mg2+有利于甘藍幼苗的生長發育。
[1] 賈敬敦,張 富.依靠科技創新推進我國鹽堿地資源可持續利用[J].中國農業科技導報,2014,16(5):1-7.
[2] 陳建平.江蘇省沿海地區發展鹽堿地植棉戰略思考[J].中國棉花,2014,41(4):14-15.
[3] 張培通,郭文琦,李春宏.枸杞在江蘇沿海灘涂鹽堿地的應用前景及實用栽培技術[J].江蘇農業科學,2014,42(3):197-199.
[4] 劉正祥,張華新,楊 升,等.NaCl脅迫對沙棗幼苗生長和光合特性的影響[J].林業科學,2014,50(1):32-40.
[5] 黃小云,陶 鵬,王五宏,等.結球甘藍離體葉片不定芽的再生研究[J].浙江農業科學,2014(1):34-38.
[6] 董 輝,顧愛俠,軒淑欣,等.大白菜—結球甘藍單體異附加系AC3營養成分分析及評價[J].營養學報,2014,36(4):401-402.
[7] 王肖紅,曾愛松,高 兵,等.結球甘藍不定芽誘導及遺傳轉化的影響因子[J].江蘇農業學報,2013,29(4):918-920.
[8] 劉俊波,范鳳翠,李志宏,等.不同根土空間對結球甘藍生長貢獻率的影響[J].江蘇農業科學,2014,42(10):147-149.
[9] 吳國平,王建華,王麗娟,等.幼苗春化特性對結球甘藍開花及繁種的影響[J].江蘇農業科學,2013,41(6):121-122.
[10] VERMA R,MAURYA B R,MEENA V S.Integrated effect of bio-organics with chemical fertilizer on growth,yield and quality of cabbage(Brassica oleracea var.capitata)[J].Indian Journal of Agricultural Sciences,2014,84(8):914-919.
[11] CHOI S H,PARK S,LIM Y P,et al.Metabolite profiles of glucosinolates in cabbage varieties(Brassica oleracea var.capitata)by season,color,and tissue position[J].Horticulture,Environment,and Biotechnology,2014,55(3):237-247.
[12]夏尚光,張金池,梁淑英.NaCl脅迫對3種榆樹幼苗生理特性的影響[J].河北農業大學學報,2008,31(2):52-56.
[13]尤 佳.鹽脅迫對鹽生植物黃花補血草幼苗生理生化特性的影響[D].蘭州:西北師范大學,2012.
[14]王家源.苦楝種苗耐鹽脅迫的生理響應機制研究[D].南京:南京林業大學,2013.
[15]孟繁昊,王 聰,徐壽軍.鹽脅迫對植物的影響及植物耐鹽機理研究進展[J].內蒙古民族大學學報:自然科學版,2014,29(3):315-318.
[16]王寶山.逆境植物生物學[M].北京:高等教育出版社,2010:209-215.
[17]謝英贊,何 平,王朝英,等.外源 Ca2+、SA、NO對鹽脅迫下決明幼苗生理特性的影響[J].西南大學學報:自然科學版,2013,35(3):36-43.
[18] PARIDA A K,DAS A B,MITTRA B.Effects of salt on growth,ion accumulation,photosynthesis and leaf anatomy of the mangrove,Bruguiera parviflora [J].Trees,2004,18(2):167-174.
[19] BERNSTEIN N,MEIRI A,ZILBERSTAINE M.Root growth of avocado is more sensitive to salinity than shoot growth[J].Journal of the American Society for Horticultural Science,2004,129(2):188-192.
[20]王樹鳳,胡韻雪,孫海菁,等.鹽脅迫對2種櫟樹苗期生長和根系生長發育的影響[J].生態學報,2014,34(4):1021-1029.
[21]譚會娟,李新榮,趙 昕.紅砂愈傷組織適應鹽脅迫的滲透調節機制研究[J]. 中國沙漠,2011,31(5):1119-1123.
[22]張麗麗,張 戰,趙一洲,等.NaCl脅迫對水稻苗期生長及離子吸收和轉運的影響[J].北方水稻,2014,44(3):10-13.
[23]譚會娟,賈榮亮,劉玉冰,等.NaCl脅迫下紅砂愈傷組織中主要離子累積特征的研究[J].中國沙漠,2010,30(6):1305-1310.
[24]王樹鳳,胡韻雪,李志蘭,等.鹽脅迫對弗吉尼亞櫟生長及礦質離子吸收、運輸和分配的影響[J].生態學報,2010,30(17):4609-4616.
[25] WEI W,BILSBORROW P E,HOOLEY P,et al.Salinity induced differences in growth,ion distribution and partitioning in barley between the cultivar Maythorpe and its derived mutant golden promise[J].Plant and Soil,2003,250(2):183-191.
[26]楊立飛,朱月林,胡春梅,等.NaCl脅迫對營養液栽培嫁接黃瓜生物量及離子分布的影響[J].西北植物學報,2007,26(12):2500-2505.
[27]劉正祥,張華新,楊秀艷,等.NaCl脅迫下沙棗幼苗生長和陽離子吸收、運輸與分配特性[J].生態學報,2014,34(2):326-336.
[28]周俊國,朱月林,楊立飛,等.NaCl脅迫下中國南瓜雜交種和黑籽南瓜植株離子吸收與積累特性研究[J].植物營養與肥料學報,2008,14(3):546-551.
[29]王 冉,陳貴林,宋 煒,等.NaCl脅迫對兩種南瓜幼苗離子含量的影響[J].植物生理與分子生物學學報,2006,32(1):94-98.
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
媽媽寶寶(2017年3期)2017-02-21 01:22:28
爆笑show(2016年7期)2017-02-09 09:36:13
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
