不同生境小花山桃草自然種群表型變異與協(xié)變
2015-01-02 08:12:50劉龍昌杜改改司衛(wèi)杰王菲羅海軍周正軍
草業(yè)學(xué)報 2015年7期
關(guān)鍵詞:植物
劉龍昌,杜改改,司衛(wèi)杰,王菲,羅海軍,周正軍
(1.河南科技大學(xué)林學(xué)院,河南 洛陽471003;2.中國林業(yè)科學(xué)研究院林業(yè)研究所,北京100091)
外來種入侵不僅導(dǎo)致生物多樣性的減少或喪失,威脅區(qū)域生態(tài)安全,而且會對經(jīng)濟造成嚴(yán)重影響[1]。探明入侵種的入侵機制對有效控制其擴散和減少對入侵地造成的危害具有重要意義。
表型可塑性和遺傳分化是植物適應(yīng)異質(zhì)生境的兩種不同策略,但二者并不矛盾和排斥,均有利于外來植物適應(yīng)入侵地的新生境[2-3]。入侵物種往往比非入侵種具有更高的表型可塑性[4-5],表型可塑性通過改變植物的形態(tài)、生長、生物量分配和生理特性等來獲取資源,從而占據(jù)多樣的生境,增強其入侵能力[6-7]。對于遺傳多樣性低的外來植物(如自花授粉植物或克隆植物)來說,表型可塑性是其成功入侵和擴張過程的關(guān)鍵因素[2]。植物進入新生境后,在新環(huán)境壓力脅迫下遺傳物質(zhì)會發(fā)生快速適應(yīng)性變化,通過進化適應(yīng)產(chǎn)生表型變異以適應(yīng)新環(huán)境。由于表型特征是自然選擇的直接承受者,所以表型進化比基因進化要快得多。許多研究表明,入侵植物能通過遺傳分化以適應(yīng)不同的選擇壓力,即對不同環(huán)境做出局域適應(yīng)[8-9]。區(qū)域性梯度變化的溫度、降水和光周期等常引起植物的適應(yīng)性進化[10-11],生境因子如土壤水分、肥力等也是引起植物適應(yīng)性進化的重要原因[12-13]。
由于生理以及遺傳發(fā)育的相關(guān)性,植物在經(jīng)歷不同環(huán)境的自然選擇時,表型性狀間存在著表型協(xié)變(表型整合)和權(quán)衡關(guān)系。選擇作用可使不同的表型性狀具有不同式樣和不同程度的可塑性,一個性狀在不同環(huán)境下的穩(wěn)定性可能正是由于其他性狀具有可塑性的結(jié)果[14]。
小花山桃草(Gauraparviflora)是柳葉菜科山桃草屬一年生或越年生自花授粉植物,原產(chǎn)北美中東部。因其具有適應(yīng)性廣、繁殖能力強等特點,近幾年在我國呈快速擴散蔓延態(tài)勢,成為我國危害性較大的外來入侵植物之一[15-16],在一些地區(qū)已發(fā)展成惡性雜草[17]。小花山桃草在義馬市北露天礦不同排矸年限的矸石山均有分布。不同排矸年限矸石山在土壤水分、養(yǎng)分條件等方面差異很大[18],這為研究不同生境下小花山桃草種群表型性狀變異規(guī)律提供了良好條件。我們以分布于該矸石山的自然種群為對象,在小花山桃草果實成熟期,對16個表型性狀進行了觀測,分析不同種群植株表型性狀在種群內(nèi)、種群間的變化特點及其環(huán)境依賴性,以期為探明小花山桃草表型變異規(guī)律和機制提供線索。
1 材料與方法
1.1 研究區(qū)域概況
研究區(qū)位于河南省義馬市北露天礦煤矸石山(北緯34°41′-34°46′,東經(jīng)111°57′-111°59′),屬暖溫帶大陸性季風(fēng)氣候,年平均氣溫12.4℃,極端最高氣溫41℃,極端最低氣溫-18.5℃;無霜期平均215d;平均日照時數(shù)2257.4h;平均降雨量為666.9mm,最多為1013.6mm,最少為456.1mm;降水多集中于7-9月份,占全年降水量的60%;多年年平均蒸發(fā)量2052mm。
1.2 取樣方法
根據(jù)矸石山堆積形成的時間和立地條件,在全面調(diào)查的基礎(chǔ)上,選取相距1km以上且生境差異明顯的5個種群為研究對象[18],各種群的生境特征見表1。于2012年7月小花山桃草果實成熟期,在小花山桃草集中連片分布處設(shè)置20m×20m的樣地,再在每個樣地中沿對角線間隔5m布設(shè)5m×5m的樣方,每個樣地設(shè)3個樣方。在每樣方中隨機取10株生長正常的健康植株,用鐵锨將植株連根挖出,清理干凈后裝入紙袋帶回實驗室,在80℃的烘箱中干燥處理72h。
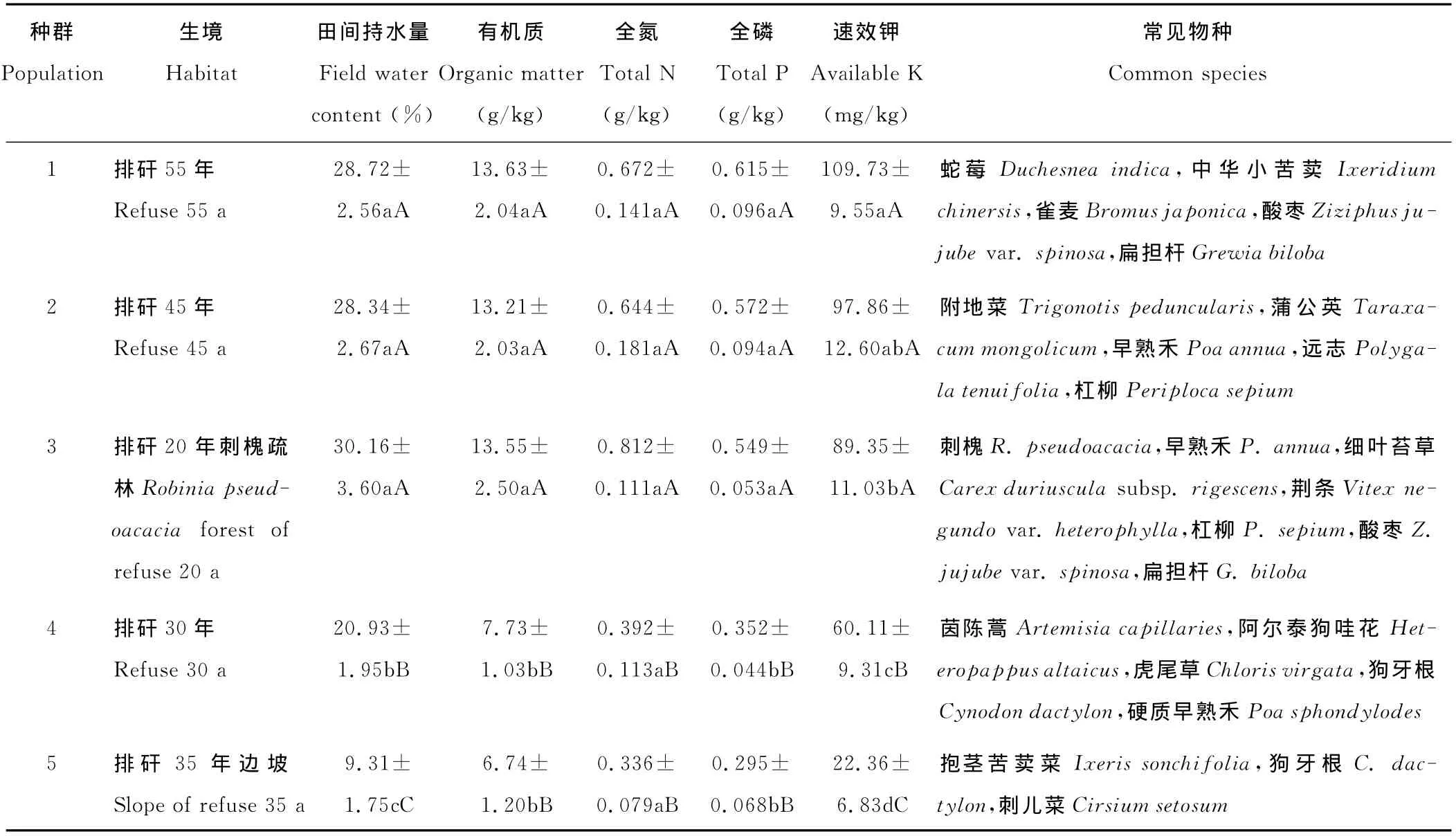
表1 小花山桃草5個種群的生境概況Table 1 Information of G.parviflorain five sample site
1.3 性狀選取和觀測方法
性狀選取時兼顧地上和地下部分、營養(yǎng)和繁殖性狀,共選取16個表型性狀:株高、節(jié)間距、莖粗、葉長、葉寬、主根長、分枝數(shù)(植株分枝數(shù))、主根重、莖重、葉重、果長、果寬、單果種子數(shù)、單株果數(shù)、單果重和比葉面積。葉面積用掃描儀和數(shù)字化方法獲得,以葉片面積除以葉片質(zhì)量得到比葉面積(SLA,cm2/g);株高和主根長度用直尺測量,其他指標(biāo)測量均用游標(biāo)卡尺測量;用萬分之一天平測量單果重、主根重、莖重和葉重,單果重采用單株果實總重與單株果實數(shù)目的比值來計算;每株隨機選取10個果實,剝?nèi)》N子,統(tǒng)計種子數(shù),其平均值為單果種子數(shù)。
1.4 土壤因子測定
選取對植物生長發(fā)育影響顯著、反映土壤水分和養(yǎng)分狀況的土壤因子,包括田間持水量,有機質(zhì)、全氮、全磷和速效鉀含量。在每個樣方中心取1處土壤樣點,挖取0~30cm土層的混合土壤樣品(具體做法為,用土鉆分別采集0~10cm,10~20cm和20~30cm土壤樣品,組成混合樣品),帶回實驗室進行分析。其中,田間持水量和有機質(zhì)含量參考劉龍昌等[18]的研究結(jié)果;全氮測定采用凱氏定氮法,全磷測定采用酸溶-鉬銻抗比色法,速效鉀測定采用醋酸銨溶-火焰光度法[19],測定結(jié)果見表1。
1.5 數(shù)據(jù)分析
計算各表型性狀種群內(nèi)和種群間的平均數(shù)、標(biāo)準(zhǔn)差和變異系數(shù),表型性狀的變異系數(shù)反映了種群內(nèi)表型性狀的變異水平[20]。
用巢氏方差分析法將表型性狀變異分解為種群間變異和種群內(nèi)變異,以探究種群間表型變異特征,各部分方差分量百分比說明了變異來源[21]。用表型分化系數(shù)(Vst)反映群體間表型分化情況,公式為,其中為群體間的方差分量,為群體內(nèi)的方差分量[22-23]。
用主成分分析法研究種群間不同性狀的協(xié)變情況。若多個性狀在第一主成分(或第二主成分)特征向量中的特征值大于0.5且符號相同,則說明這些性狀的協(xié)變性較大。種群間協(xié)變性狀相同且協(xié)變數(shù)目越多,表明性狀的分化程度越低,性狀具有相似的整合格局[24]。
采用分層聚類法探討5個種群間表型性狀的整體相似性[24],相似系數(shù)計算采用歐氏距離法,分析前數(shù)據(jù)進行標(biāo)準(zhǔn)化變換;對各表型性狀和生境土壤因子進行相關(guān)性分析,以探究生境因子對表型性狀的影響。所有統(tǒng)計和分析在SPSS 18.0、SAS 6.0和 Excel 2003統(tǒng)計軟件上進行。
2 結(jié)果與分析
2.1 種群內(nèi)和種群間表型變異特征
變異系數(shù)的大小可間接反映群體的表型多樣性豐富程度,變異系數(shù)大說明該群體的性狀變異幅度高,表型多樣性豐富;反之則說明該群體的性狀變異幅度低,表型多樣性低[25]。由表型性狀變異系數(shù)(表2)可知,不同種群各性狀平均變異系數(shù)由大到小依次為:種群2>種群5>種群1>種群3>種群4,平均變異系數(shù)最大的是拍矸時間45年的種群(種群2),為0.386,最小的是拍矸30年的種群(種群4),為0.338。平均變異系數(shù)<0.2的性狀有果長、果寬和比葉面積,說明這3個性狀最穩(wěn)定。平均變異系數(shù)>0.4的性狀有:葉寬、主根重、莖重、葉重和單株果數(shù),其中葉重和單株果數(shù)變異系數(shù)最大(>0.8),說明它們最不穩(wěn)定。
單果種子數(shù)和單果重在種群內(nèi)和種群間的差異均不顯著(P>0.05),其他性狀在種群間的差異均達(dá)極顯著水平(表3),說明各種群生長狀況差異較大。其中節(jié)間距、莖粗、葉長、葉寬、主根長、主根重、莖重、葉重、果寬、單株果數(shù)和比葉面積變化較大(F值大于10),葉寬和節(jié)間距在種群間的變化最大(F值大于50)。

表3 不同生境小花山桃草種群各性狀單因素方差分析(平均值±標(biāo)準(zhǔn)偏差)Table 3 One Way ANOVA of phenotypic traits variance between G.parviflora populations from different habitat(mean±SE)
在16個表型性狀中,株高、莖粗、單果種子數(shù)、單株果數(shù)、單果重和比葉面積6個性狀在種群內(nèi)的差異均不顯著;葉長、主根重、莖重和葉重4個性狀差異顯著(P<0.05);節(jié)間距、葉寬、主根長、分枝數(shù)、果長和果寬6個性狀差異極顯著(P<0.01),其中葉寬的變化最大。
2.2 種群內(nèi)和種群間表型分化
16個表型性狀的Vst變異范圍為26.85%~96.98%,平均為73.03%(表4),說明種群間變異是表型變異的主要來源,也表明不同排矸年限環(huán)境(即生境)對小花山桃草各種群表型變化有決定性影響。
果長、分枝數(shù)和葉寬的表型分化系數(shù)較低(<50%),尤其是果長僅為26.85%,說明果長、分枝數(shù)和葉寬3個性狀在種群內(nèi)有較大的變異,受種群內(nèi)微環(huán)境變化的影響強烈。株高、節(jié)間距、莖粗、主根長、葉重、單果種子數(shù)、單株果數(shù)和比葉面積8個性狀的表型分化系數(shù)達(dá)80%以上,種群間變異大于種群內(nèi)變異,表明這些性狀受種群內(nèi)微環(huán)境的影響較小,受不同生境水肥狀況影響較大。
2.3 不同生境種群表型整合格局
不同生境下5個種群的株高、莖粗、葉長、葉寬、主根重、莖重、葉重和單株果數(shù)8個性狀的第一主成分特征向量值均大于0.5,表明這些性狀的變化具有明顯的協(xié)同性(表5);種群2和種群3的節(jié)間距、種群1的主根長以及種群2和種群4的分枝數(shù)的第一主成分特征值大于0.5。在其他種群中,這3個性狀的第一主成分特征值雖然小于0.5,但仍可達(dá)到0.34~0.47,同樣表現(xiàn)出大體相同的協(xié)同變化趨勢。
單果重、果長、果寬3個果實性狀以及比葉面積與生長性狀(包括株高、節(jié)間距、莖粗、葉長、葉寬、主根長、主根重、分枝數(shù)、莖重和葉重)之間的協(xié)變性較弱。盡管這4個性狀在第二主成分中有較大的特征向量,但它們之間以及它們與生長性狀之間的協(xié)變性較低,協(xié)變方向也不一致。
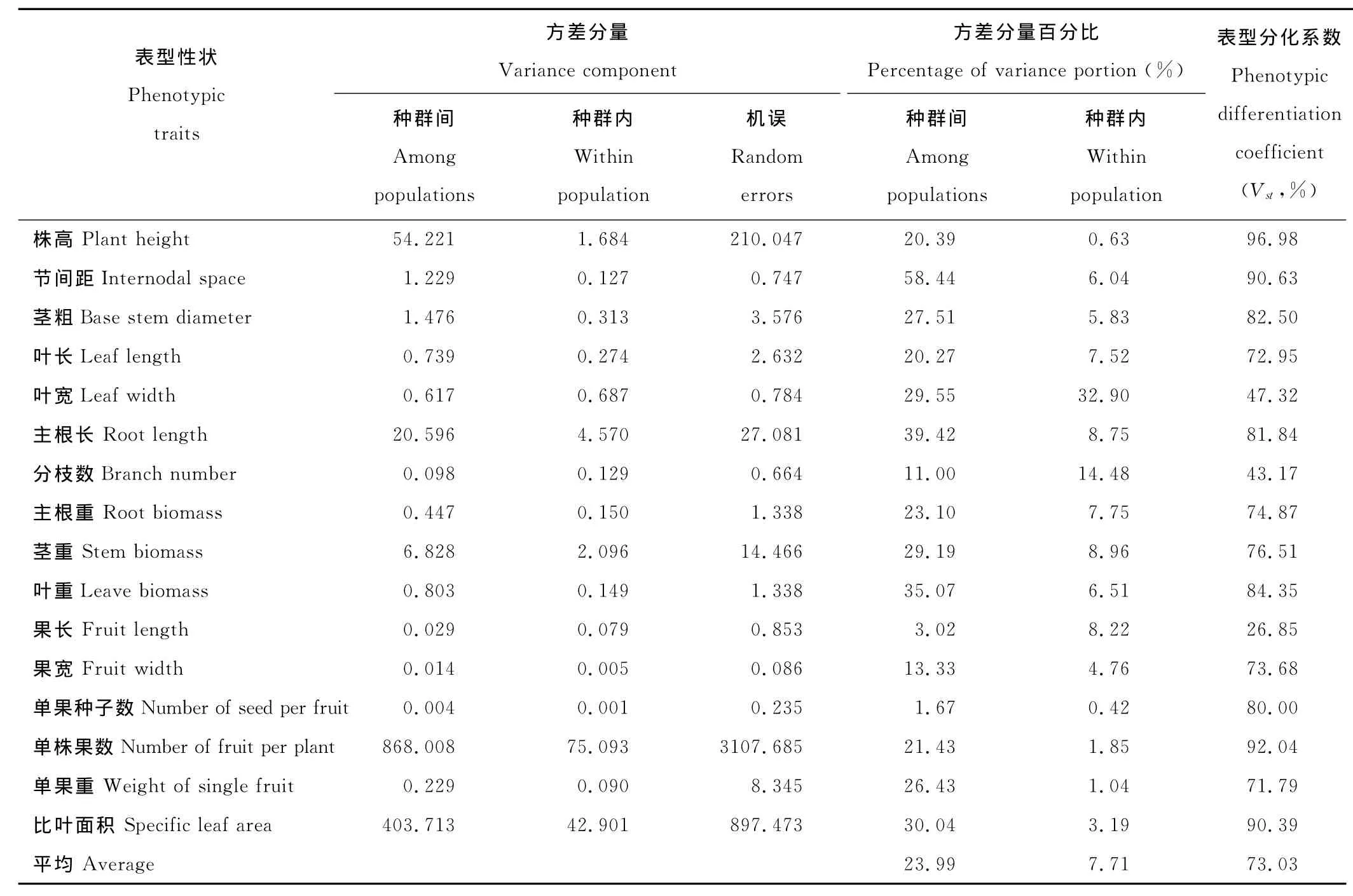
表4 不同生境下小花山桃草表型性狀的方差分量與居群間表型分化系數(shù)Table 4 The percentage of variance components and differentiation coefficients of phenotypic traits(Vst)within and among population of G.parviflorain different habitat

表5 不同生境下小花山桃草種群16個表型性狀主成分分析Table 5 Principal component analysis of sixteen phenotypic traits of G.parviflorain different habitat
2.4 小花山桃草表型性狀與生境因子的相關(guān)性
小花山桃草各性狀與生境因子的相關(guān)關(guān)系分析見表6,10個生長性狀中有9個(包括株高、節(jié)間距、莖粗、葉長、葉寬、主根長、主根重、莖重和葉重)與各生境因子存在顯著或極顯著的正相關(guān)關(guān)系;4個果實性狀中有3個(包括果寬、單果重和單果種子數(shù))與生境因子相關(guān)性低。這表明相對于生長性狀,果實性狀較為穩(wěn)定,不易受環(huán)境影響。果實長度與各生境因子存在顯著負(fù)相關(guān)關(guān)系,在貧瘠干旱的環(huán)境條件下,小花山桃草的果實更細(xì)長一些,果實中的種子數(shù)也少一些。分枝數(shù)與土壤含水量和全氮量呈顯著正相關(guān),與其他生境因子無顯著相關(guān)關(guān)系,說明分枝數(shù)主要受土壤含水量和全氮含量的制約。
2.5 小花山桃草種群間表型相似性的聚類分析
采用歐式距離法對不同生境的5個種群進行聚類分析(圖1)。結(jié)果表明,在閾值為10時可把5個種群分為2組。種群1、種群2和種群3被分為一組,結(jié)合表3可知,這3個種群在株高、節(jié)間距、莖粗、葉長、莖重、單果重6個表型性狀上比較相似。而種群4和種群5在節(jié)間距、莖粗、葉長、葉寬、主根重、莖重、葉重和單果重等表型性狀上較為相似,均明顯低于前3個種群,因此單獨聚為一類。
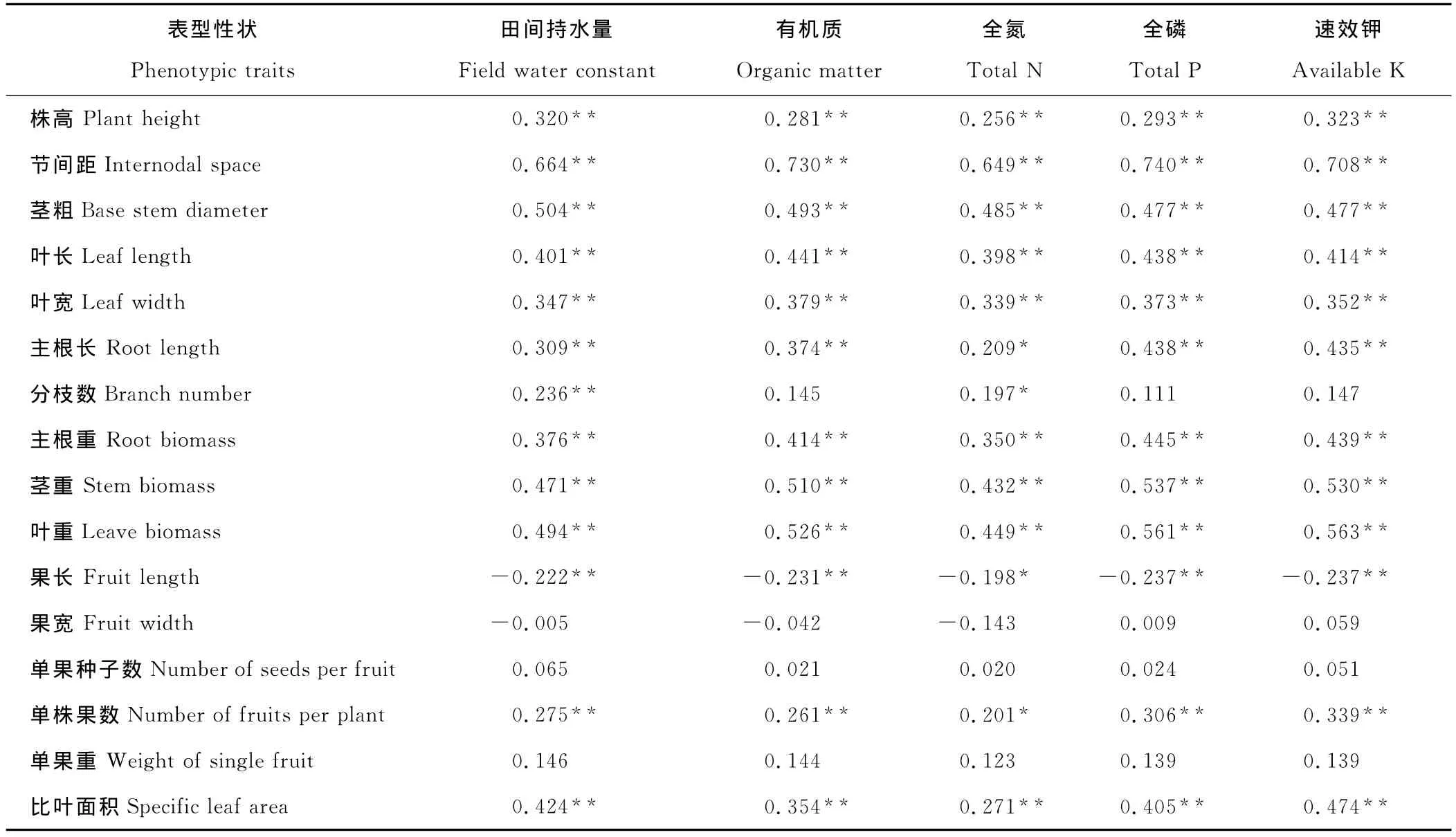
表6 小花山桃草表型性狀與生境因子之間的相關(guān)系數(shù)Table 6 Correlation coefficient between phenotypic traits and habitat factors of G.parviflora
結(jié)合各種群的生境條件(表1)可知,生境條件較好的3個種群聚在一個分支,生境條件較差的2個種群聚在另一分支。由此不難看出,小花山桃草表型特征受種群間環(huán)境條件(水、肥)的強烈影響。
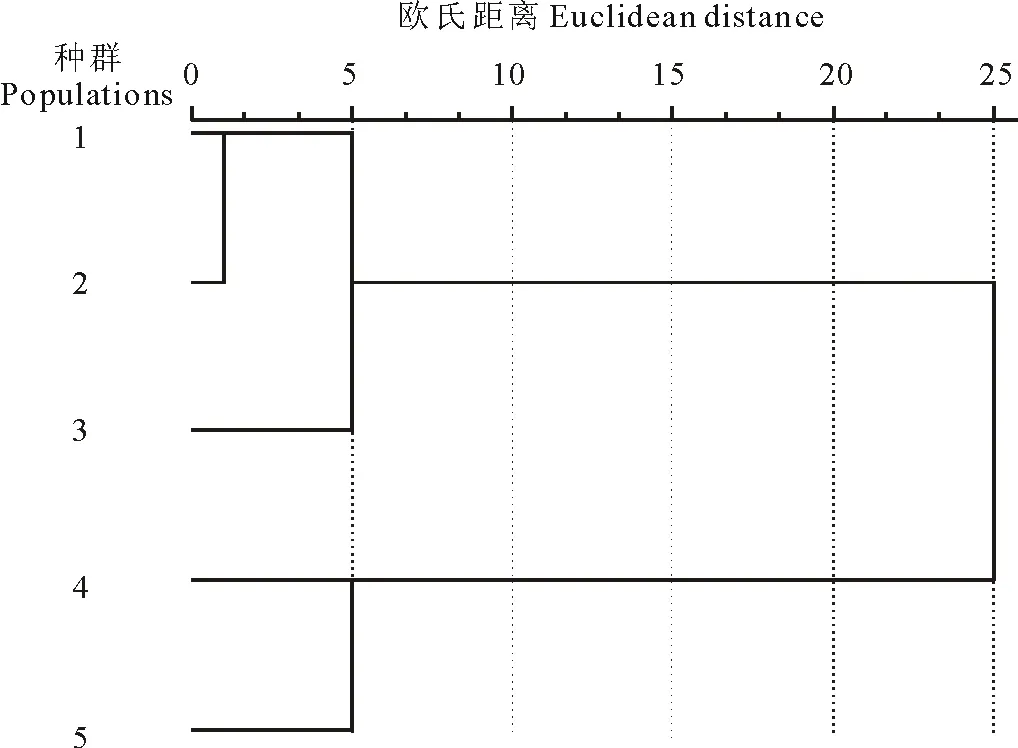
圖1 基于小尺度環(huán)境下小花山桃草5個種群16個表型性狀的聚類圖Fig.1 Hierarchical cluster based on sixteen phenotypic traits of five G.parviflora populations in different habitat
3 討論
表型多樣性作為遺傳多樣性與環(huán)境多樣性的綜合體現(xiàn),是了解生物遺傳變異的重要線索,也是生物多樣性的重要內(nèi)容[26]。植物表型性狀的變異幅度越大,表型多樣性越豐富[27]。本研究顯示,小花山桃草5個群體16個表型性狀的平均變異系數(shù)為36.30%(繁殖性狀的平均變異系數(shù)為31.42%)。明顯高于大多數(shù)非入侵植物的變異系數(shù),如滇北球花報春(Primuladenticulatassp.Sinodenticulata)的7.08%、野扁桃(Amygdalusledebouriana)的18.66%、野生芒(Miscanthussinensis)的 27.35%[28-30]。也顯著高于一些入侵植物的變異系數(shù),如刺萼龍葵(Solanumrostratum)的13.60%、劍葉金雞菊(Coreopsislanceolata)的8.24%(繁殖性狀)[13,31]。但低于有些入侵植物的變異系數(shù),如銀膠菊(Parthenium hysterophorus)的54.86%、喜旱蓮子草(Alternantheraphiloxeroides)的59.83%[12,32]。表明小花山桃草的表型多樣性較為豐富,暗示它對異質(zhì)生境有較為廣泛的適應(yīng)能力[33-34]。
一般來說,營養(yǎng)性狀對不同生境的反應(yīng)比繁殖性狀更為敏感,更容易發(fā)生變異。所以,營養(yǎng)性狀的變異系數(shù)常高于繁殖性狀的變異系數(shù)[35]。本研究結(jié)果也支持這一結(jié)論,小花山桃草種群營養(yǎng)性狀的變異系數(shù)(平均為40.52%)明顯高于繁殖性狀的變異系數(shù)(平均為31.42%)。繁殖性狀相對穩(wěn)定有助于保持子代與母代間遺傳物質(zhì)的穩(wěn)定,而較高的營養(yǎng)性狀變異有利于植物適應(yīng)多變的環(huán)境。
表型性狀的變化對環(huán)境具有明顯的依賴性,不同性狀對自然選擇的反應(yīng)不同[24]。本研究結(jié)果表明,在觀測的16個表型性狀中,有11個性狀在種群間表現(xiàn)出明顯的差異,包括株高、節(jié)間距、莖粗、葉長、主根長、主根重、莖重、葉重、果寬、單株果數(shù)和比葉面積,它們的表型分化系數(shù)均在71%以上。尤其是株高、莖粗、單株果數(shù)和比葉面積4個性狀在種群內(nèi)的差異不顯著,而在種群間的差異均達(dá)到極顯著水平,表型分化系數(shù)均在82%以上,表明種群間主要生境因子的差異是上述表型性狀變化的決定因素;節(jié)間距、葉長、葉寬、主根長、分枝數(shù)、主根重、莖重、葉重、果長和果寬在種群內(nèi)和種群間均表現(xiàn)出明顯的差異,尤其是節(jié)間距、葉寬、主根長、分枝數(shù)、果長和果寬6個性狀在種群內(nèi)的差異均達(dá)極顯著水平,表明這些性狀受種群內(nèi)微環(huán)境的影響較大。同時,研究還發(fā)現(xiàn),果長、分枝數(shù)和葉寬3個性狀對梯度生境因子和微環(huán)境條件均比較敏感,種群間和種群內(nèi)的方差分量比較接近,這應(yīng)該是其表型分化系數(shù)較低的主要原因。
功能性狀株高、比葉面積和單株果數(shù)在種群間表現(xiàn)出極高的變異性,表型分化系數(shù)均在90%以上,且與各生境因子呈顯著正相關(guān),這說明它們易受梯度生境因子影響。在充足的水分和良好的養(yǎng)分條件下,小花山桃草通過生長空間和葉面積的擴大捕獲更多的光能,積累更多的營養(yǎng)物質(zhì),生產(chǎn)更多的果實和種子,從而實現(xiàn) “表現(xiàn)最大化”[2]。
植株的分枝數(shù)常與土壤含水量呈正相關(guān)關(guān)系[36],本研究進一步證實了這一點。分枝數(shù)的這種可塑性變異具有生態(tài)適應(yīng)意義。在水分條件充足時,植物通過增加分枝數(shù)和冠幅,擴大生長空間,積累更多營養(yǎng),以實現(xiàn)生理功能的最大化。在水分不足時,通過減少分枝數(shù)和冠幅,減小枝葉構(gòu)件間的競爭作用和水分散失[37-38],以保證基本生理功能的維持。
從理論上說,同時具有豐富遺傳變異和高度表型可塑性的植物能夠更有效地適應(yīng)復(fù)雜多變的環(huán)境[2]。許多外來植物的成功入侵是其表型可塑性和局域適應(yīng)共同作用的結(jié)果[39]。在一個范圍很小的樣地內(nèi),植株間的遺傳變異往往很低(自花授粉和克隆植物尤為如此),所以種群內(nèi)植株間表型性狀的差異可能主要是表型可塑性的結(jié)果[23]。而生境的異質(zhì)性使得各種群承受了不同的選擇壓力,如果這種選擇壓力大于遺傳漂變和基因流的作用,則處于不同生境的種群可能在形態(tài)、生理和物候等表型性狀上產(chǎn)生分化,出現(xiàn)局部適應(yīng)現(xiàn)象[40]。本研究表明,小花山桃草種群間表型變異很高,種群間表型性狀分化遠(yuǎn)大于種群內(nèi)的變異(平均表型分化系數(shù)為73.03%)。同時,其多數(shù)表型性狀(16個表型性狀中有10個)在種群內(nèi)也有較高的變異。這暗示小花山桃草可能同時具有較高的表型可塑性和較強的適應(yīng)進化能力。為進一步明確這一問題,進一步開展不同尺度環(huán)境下表型變異規(guī)律和同質(zhì)生物園對比研究十分必要。
植物能夠通過生理整合調(diào)控其表型可塑性[41]。表型性狀間的協(xié)同性越高,植物體的整體功能就會越高效[42]。本研究表明,不同生境下小花山桃草種群的株高、莖粗、葉長、葉寬、莖重、葉重、主根重和單株果數(shù)具有一致的整合格局。說明小花山桃草地上部分(株高、莖重和葉重等)與地下部分(主根重)之間、植株生物量(莖重、葉重和主根重)與繁殖輸出(單株果數(shù))之間具有生理功能上的權(quán)衡性。由于單株果數(shù)和單果種子數(shù)是決定植物相對適合度高低的重要因素,所以莖重、葉重、主根重與單株果數(shù)的協(xié)同變化具有適應(yīng)意義[24]。
[1]Mack R N,Simberloff D,Lonsdale W M,etal.Biotic invasions:causes,epidemiology,global consequences,and control.Ecological Applications,2000,10(3):689-710.
[2]Geng Y P,Zhang W J,Li B,etal.Phenotypic plasticity and invasiveness of alien plants.Biodiversity Science,2004,12(4):447-455.
[3]Qin R M,Zheng Y L,Valiente-Banuet A,etal.The evolution of increased competitive ability,innate competitive advantages,and novel biochemical weapons act in concert for a tropical invader.New Phytologist,2013,197(3):979-988.
[4]Feng Y L,F(xiàn)u G L,Zheng Y L.Specific leaf area relates to the differences in leaf construction cost,photosynthesis,nitrogen allocation,and use efficiencies between invasive and noninvasive alien congeners.Planta,2008,228(3):383-390.
[5]Zhao X J,Liu W Y,Zhou M.Lack of local adaptation of invasive crofton weed(Ageratinaadenophora)in different climatic areas of Yunnan Province,China.Journal of Plant Ecology,2013,6(4):316-322.
[6]Liu J,Li J M,Yu H,etal.The relationship between functional traits and invasiveness of alien plants.Biodiversity Science,2010,18(6):569-576.
[7]Huang W,Carrillo J,Ding J Q,etal.Invader partitions ecological and evolutionary responses to above-and belowground herbivory.Ecology,2012,93(11):2343-2352.
[8]Dlugosch K M,Parker I M.Invading populations of an ornamental shrub show rapid life history evolution despite genetic bottlenecks.Ecology Letters,2008,11(7):701-709.
[9]Gao L X,Geng Y P,Li B,etal.Genome-wide DNA methylation alterations ofAlternantheraphiloxeroidesin natural and manipulated habitats:Implications for epigenetic regulation of rapid responses to environmental fluctuation and phenotypic variation.Plant Cell Environment,2010,33(11):1820-1827.
[10]Maron J L,Elmendorf S C,VilàM.Contrasting plant physiological adaptation to climate in the native and introduced range ofHypericumperforatum.Evolution,2007,61(8):1912-1924.
[11]Zhen Y,Ungerer M C.Clinal variation in freezing tolerance among natural accessions ofArabidopsisthaliana.New Phytologist,2008,177(2):419-427.
[12]Pan X Y,Geng Y P,Zhang W J,etal.Cover shift and morphological plasticity of invasiveAlternantheraphiloxeroidesalong a riparian zone in South China.Chinese Journal of Plant Ecology,2006,30(5):835-843.
[13]Chen T Y,Liu Z H,Lou A R.Phenotypic variation in populations ofSolanumrostratumin different distribution areas in China.Chinese Journal of Plant Ecology,2013,37(4):344-353.
[14]Klingenberg C P.Morphological integration and developmental modularity.Annual Review of Ecology,Evolution,and Systematics,2008,39:115-132.
[15]Du W B,Ye Y Z,Peng S L.Study on the seasonal growing dynamics and invasive characteristics ofGauraparviflora.Acta Ecologica Sinica,2003,23(8):1679-1684.
[16]Liu L C,Xu L,F(xiàn)eng P,etal.Seed dormancy and germination characteristics ofGauraparviflora,an exotic weed species in China.Acta Ecologica Sinica,2014,34(24):7338-7349.
[17]Wan F H,Liu Q R,Xie M,etal.Gauraparviflora.Biological Invasions:Color Illustrations of Invasive Alien Plants in China[M].Beijing:Science Press,2012:202-203.
[18]Liu L C,Song M Y,Xu G R,etal.Land suitability evaluation of vegetation restoration of coal-gangue pile in Yima Area.Journal of Fujian College of Forestry,2012,32(1):70-75.
[19]Bao S D.Soil and Agricultural Chemistry Analysis(3th edition)[M].Beijing:China Agriculture Press,2000:14-38.
[20]Wang D,Pang C H,Gao Y H,etal.Phenotypic diversity ofAcerginnala(Aceraceae)populations at different altitude.Acta Botanica Yunnanica,2010,32(2):117-125.
[21]Zhang H Q,An L J,Zhu Y G.Geographical variation of morphology characters for natural populations ofPinuskoraiensis.Acta Ecologica Sinica,1999,19(6):932-938.
[22]Ge S,Wang M X,Chen Y W.An analysis of population genetic structure of masson pine by isozyme technique.Scientia Silvae Sinice,1988,24(4):399-409.
[23]Hedrick P W.A standardized genetic differentiation measure.Evolution,2005,59(8):1633-1638.
[24]Li L,Liu T,Liu B,etal.Phenotypic variation and covariation among natural populations ofArabidopsisthalianain North Xinjiang.Biodiversity Science,2010,18(5):497-508.
[25]Dunn C P.Keeping taxonomy based in morphology.Trends in Ecology and Evolution,2003,18(6):270-271.
[26]Guo N,Yang S H,Ge W Y,etal.Phenotypic diversity of natural populations ofRosalaxaRetz.in Tianshan Mountains of Xinjiang.Acta Horticulturae Sinica,2011,38(3):495-502.
[27]Zhang C X,Ming J,Liu C,etal.Phenotypic variation of natural populations inLiliumregaleWilson.Acta Horticulturae Sinica,2008,35(8):1183-1188.
[28]Zhang R L,Jia Y,Zhang Q X.Phenotypic variation of natural populations ofPrimuladenticulatassp.sinodenticulata.Biodiversity Science,2008,16(4):362-368.
[29]Zeng B,Luo S P,Li J,etal.Morphological variations in natural populations ofAmygdalusledebouriana.Biodiversity Science,2008,16(5):484-491.
[30]Nie G,Zhang X Q,Huang L K,etal.Phenotypic variation of wildMiscanthussinensispopulations from southwestern China.Acta Prataculturae Sinica,2013,22(5):52-61.
[31]Zeng J J,Xiao Y A,Sun M.Reproductive traits associated with invasiveness inCoreopsislanceolata.Chinese Journal of Plant Ecology,2010,34(8):966-972.
[32]Pu G Z,Tang S C,Pan Y M,etal.Phenotypic plasticity and modular biomass of invasivePartheniumhysterophorusin dif-ferent habitats in south China.Guihaia,2010,30(5):641-646.
[33]Du N,Zhang X R,Wang W,etal.Foliar phenotypic plasticity of a warm-temperate shrub,Vitexnegundovar.heterophylla,to different light environments in the field.Acta Ecologica Sinica,2011,31(20):6049-6059.
[34]Richter S,Kipfer T,Wohlgemuth T,etal.Phenotypic plasticity facilitates resistance to climate change in a highly variable environment.Oecologia,2012,169(1):269-279.
[35]Xing Y,Zhao X,Dong K H,etal.A study on morphological variation of different populations ofLespedezadavurica.Acta Prataculturae Sinica,2008,17(4):26-31.
[36]Xu G F,Shen S C,Zhang F D,etal.Effect of soil-water conditions on survival rate and morphological plasticity of clonal plantMikaniamicranthaH.B.Kunth.Scientia Agricultura Sinica,2013,46(15):3134-3141.
[37]Luo Y Z,Li G.The effect of water stress on growth and biomass ofMedicagosativacv.Xinjiangdaye.Acta Prataculturae Sinica,2014,23(4):213-219.
[38]Zhou Z H,Li Z,Jiao J,etal.Analysis of clonal growth patterns and branching architecture ofNitrariatangutorumclonal ramet in the Southern margin area of the Tengger Desert.Acta Prataculturae Sinica,2014,23(1):12-21.
[39]Zhang C L,Li Y P,F(xiàn)eng Y L,etal.Roles of phenotypic plasticity and local adaptation inEupatoriumadenophoruminvasions in different altitude habitats.Acta Ecologica Sinica,2009,29(4):1940-1946.
[40]Kawecki T J,Ebert D.Conceptual issues in local adaptation.Ecology Letters,2004,7(12):1225-1241.
[41]Luo D,Qian Y Q,Liu J X,etal.Phenotypic responses of a clonal plant(Buchloedactyloides)to nutrient heterogeneity.Acta Prataculturae Sinica,2014,23(3):104-109.
[42]Marcus B T,Cynthia W.Plasticity and environment-specific covariances:an investigation of floral-vegetative and within flower correlations.Evolution,2007,61(12):2913-2924.
參考文獻:
[2]耿宇鵬,張文駒,李博,等.表型可塑性與外來植物的入侵能力.生物多樣性,2004,12(4):447-455.
[6]劉建,李鈞敏,余華,等.植物功能性狀與外來植物入侵.生物多樣性,2010,18(6):569-576.
[12]潘曉云,耿宇鵬,張文駒,等.喜旱蓮子草沿河岸帶不同生境的蓋度變化及形態(tài)可塑性.植物生態(tài)學(xué)報,2006,30(5):835-843.
[13]陳天翌,劉增輝,婁安如.刺萼龍葵種群在中國不同分布地區(qū)的表型變異.植物生態(tài)學(xué)報,2013,37(4):344-353.
[15]杜衛(wèi)兵,葉永忠,彭少麟.小花山桃草季節(jié)生長動態(tài)及入侵特性.生態(tài)學(xué)報,2003,23(8):1679-1684.
[16]劉龍昌,徐蕾,馮佩,等.外來雜草小花山桃草種子休眠萌發(fā)特性研究.生態(tài)學(xué)報,2014,34(24):7338-7349.
[17]萬方浩,劉全儒,謝明,等.小花山桃草.生物入侵:中國外來入侵植物圖鑒[M].北京:科學(xué)出版社,2012:202-203.
[18]劉龍昌,宋蒙亞,徐國瑞,等.義馬煤礦矸石山植被恢復(fù)土地適宜性評價.福建林學(xué)院學(xué)報,2012,32(1):70-75.
[19]鮑士旦.土壤農(nóng)化分析(第三版)[M].北京:中國農(nóng)業(yè)出版社,2000:14-38.
[20]王丹,龐春華,高亞卉,等.茶條槭不同海拔種群的表型多樣性.云南植物研究,2010,32(2):117-125.
[21]張恒慶,安利佳,祖元剛.天然紅松種群形態(tài)特征地理變異的研究.生態(tài)學(xué)報,1999,19(6):932-938.
[22]葛頌,王明庥,陳岳武.用同工酶研究馬尾松群體的遺傳結(jié)構(gòu).林業(yè)科學(xué),1988,24(4):399-409.
[24]李磊,劉彤,劉斌,等.新疆北部擬南芥自然居群表型變異與協(xié)變.生物多樣性,2010,18(5):497-508.
[26]郭寧,楊樹華,葛維亞,等.新疆天山山脈地區(qū)疏花薔薇天然居群表型多樣性分析.園藝學(xué)報,2011,38(3):495-502.
[27]張彩霞,明軍,劉春,等.岷江百合天然群體的表型多樣性.園藝學(xué)報,2008,35(8):1183-1188.
[28]張睿鸝,賈茵,張啟翔.滇北球花報春天然群體表型變異研究.生物多樣性,2008,16(4):362-368.
[29]曾斌,羅淑萍,李疆,等.新疆野扁桃天然居群形態(tài)變異的研究.生物多樣性,2008,16(5):484-491.
[30]聶剛,張新全,黃琳凱,等.中國西南區(qū)野生芒居群表型變異研究.草業(yè)學(xué)報,2013,22(5):52-61.
[31]曾建軍,肖宜安,孫敏.入侵植物劍葉金雞菊的繁殖特征及其與入侵性之間的關(guān)系.植物生態(tài)學(xué)報,2010,34(8):966-972.
[32]蒲高忠,唐賽春,潘玉梅,等.入侵植物銀膠菊在不同生境下表型可塑性和構(gòu)件生物量.廣西植物,2010,30(5):641-646.
[33]杜寧,張秀茹,王煒,等.荊條葉性狀對野外不同光環(huán)境的表型可塑性.生態(tài)學(xué)報,2011,31(20):6049-6059.
[35]邢毅,趙祥,董寬虎,等.不同居群達(dá)烏里胡枝子形態(tài)變異研究.草業(yè)學(xué)報,2008,17(4):26-31.
[36]徐高峰,申時才,張付斗,等.土壤水分對薇甘菊不同繁殖體單位存活能力和植株表型可塑性影響.中國農(nóng)業(yè)科學(xué),2013,46(15):3134-3141.
[37]羅永忠,李廣.土壤水分脅迫對新疆大葉苜蓿的生長及生物量的影響.草業(yè)學(xué)報,2014,23(4):213-219.
[38]周資行,李真,焦健,等.騰格里沙漠南緣唐古特白刺克隆分株生長格局及枝系構(gòu)型分析.草業(yè)學(xué)報,2014,23(1):12-21.
[39]張常隆,李揚蘋,馮玉龍,等.表型可塑性和局域適應(yīng)在紫莖澤蘭入侵不同海拔生境中的作用.生態(tài)學(xué)報,2009,29(4):1940-1946.
[41]羅棟,錢永強,劉俊祥,等.克隆植物野牛草對異質(zhì)營養(yǎng)的表型可塑性響應(yīng).草業(yè)學(xué)報,2014,23(3):104-109.
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
