慢病毒介導BTLA轉染未成熟樹突狀細胞的研究
2015-01-04 09:01:26王萱易寰劉佳李敏彭福華
新醫學 2015年12期
王萱 易寰 劉佳 李敏 彭福華
基礎研究論著
慢病毒介導BTLA轉染未成熟樹突狀細胞的研究
王萱 易寰 劉佳 李敏 彭福華
目的 探討構建攜帶B和T淋巴細胞衰減因子(BTLA)基因的慢病毒載體轉染未成熟樹突狀細胞(DC)的可行性。方法根據GeneBank公布的小鼠BTLA的mRNA序列構建ORF慢病毒表達質粒,合成BTLA基因的重組慢病毒載體。原代分離培養CL57/BL小鼠骨髓來源的未成熟DC,用包含BTLA基因的重組慢病毒載體以感染復數為15轉染至未成熟DC。采用免疫熒光、反轉錄PCR檢測BTLA基因的表達。結果成功構建有活性的慢病毒載體LPP-Mm26645-Lv203-400,慢病毒滴度為3.25×109TU/ml。以重組慢病毒LPP-Mm26645-Lv203-400轉染未成熟DC 96 h后在熒光倒置顯微鏡下可觀察到被感染的陽性細胞有綠色熒光蛋白表達,反轉錄PCR檢測到感染的DC中BTLA基因的表達量比未感染的DC的表達量高(P<0.05),提示轉染成功。結論構建攜帶BTLA基因的慢病毒載體可成功轉染BTLA至小鼠骨髓來源的未成熟DC。
B和T淋巴細胞衰減因子;慢病毒;樹突狀細胞;基因轉染
B和T淋巴細胞衰減因子(BTLA)是免疫球蛋白超家族的一員,是CD28家族被發現的第3個抑制性分子,與相應受體皰疹病毒侵入介質(HVEM)結合提供抗原特異性T淋巴細胞活化的第二信號,在免疫反應早期階段發揮重要的協同抑制作用[1]。樹突狀細胞(DC)是體內最強的專職抗原提呈細胞(APC),在免疫系統中起中心調節作用[2]。DC不同表型對T淋巴細胞接受抗原刺激的反應狀態有決定性作用,未成熟DC因缺乏共刺激分子表達,不能提供T淋巴細胞活化必需的第二信號,往往導致T淋巴細胞無能或凋亡,誘導免疫耐受,因此成為近年來研究的熱點[3]。未成熟DC誘導抗原特異性免疫耐受并維持其耐受狀態,對自身免疫性疾病和移植后排斥反應的預防和治療具有重要意義,但是單純的DC治療難以達到理想效果[4]。BTLA負性共刺激分子修飾未成熟DC,抑制T淋巴細胞的活化過程,可增強免疫耐受。在本研究中,我們構建了BTLA慢病毒表達載體轉染未成熟DC,以探討此法轉染BTLA至未成熟DC的可行性,從而為進一步研究BTLA修飾的未成熟DC的免疫耐受作用提供實驗基礎。
材料與方法
一、材料與動物
含葡萄糖、谷氨酰胺、丙酮酸鈉的DMEM培養基,含L-谷氨酰胺和25mM4-羥乙基哌嗪乙磺酸(HEPES)的RPMI1640培養基、NQBB胎牛血清、青霉素-鏈霉素雙抗、IL-4(IL-4Pepro Tech)、粒細胞-巨噬細胞集落刺激因子(GM-CSF Pepro Tech)、小鼠抗小鼠MHC-Ⅱ、CD80、CD86、CD11c單克隆抗體(eBioscience)。6周雌性CL57/BL小鼠。
二、方 法
采用實時PCR(qPCR)測定重組慢病毒載體滴度。采用免疫熒光、反轉錄PCR(RT-PCR)檢測未成熟DC上BTLA基因的表達情況。
1.慢病毒載體LPP-Mm26645-Lv203-400的構建與活性檢測
根據GeneBank公布的小鼠BTLA的mRNA (NM_001037719.2),委托廣州GeneCopoeia公司合成引物,構建ORF慢病毒表達質粒LV-BTLA并構建慢病毒LPP-Mm26645-Lv203-400,采用qPCR測定慢病毒LPP-Mm26645-Lv203-400滴度。用此慢病毒感染H1299細胞,培養48 h。使用熒光顯微鏡進行增強型綠色熒光蛋白觀察,有綠色熒光表示慢病毒有活性。將慢病毒液凍存于-80℃。
2.小鼠骨髓源性未成熟DC的分離培養與流式檢測
2.1 DC的分離培養
頸椎脫臼處死CL57/BL小鼠,用無菌手術取出股骨、脛骨,剪去骨兩端,吸取PRMI1640培養基(含20%胎牛血清、1%青霉素-鏈霉素雙抗、10 ng/μl的IL-4、10 ng/μl的GM-CSF)反復沖洗骨髓腔,將骨髓細胞沖至培養基中,反復吹打培養基,得到骨髓細胞懸液。收集骨髓細胞懸液至15 ml離心管后離心(1 500 rpm,5 min),棄上清液收集骨髓細胞沉淀。以適量RPMI1640培養基充分重懸細胞后以每孔1.5 ml將細胞懸液轉移至6孔細胞板中,水平輕搖后將細胞板放入37℃、5%二氧化碳培養箱中。培養第4日進行半量換液。
2.2 DC的收集與流式檢測
培養DC的第6日,于顯微鏡下觀察細胞生長情況并拍照。收集細胞懸液至15 ml離心管后離心(1 000 rpm,8 min),收集細胞沉淀后以適量RPMI1640培養基重懸細胞。取部分細胞與PE-Cy5.5標記的CD11c、FITC標記的CD80、PE標記的CD86、APC標記的MHC-Ⅱ分子在4℃反應30 min,經磷酸鹽緩沖液(PBS)沖洗2次后用流式細胞儀檢測。
3.重組慢病毒LPP-Mm26645-Lv203-400轉染DC
按上述方法培養的DC培養至第5日,收集DC并接種于12孔細胞板,每個孔內接種3×104個DC,鋪板時細胞的融合率為50%左右,每孔培養基體積為600 μl,水平輕搖后將細胞板放入37℃、5%二氧化碳培養箱中。24 h后取出構建的慢病毒LPP-Mm26645-Lv203-400液并稀釋至0.5× 108TU/mL,進行病毒感染時細胞的融合度約為70%,以感染復數為15分別加入重組慢病毒LPPMm26645-Lv203-400,補充完全RPMI1640培養基至終體積1 ml,加入Polybrene維持其終濃度于5 μg/ml,水平輕搖后將細胞板放入37℃、5%二氧化碳培養箱中。隔日補充新鮮RPMI1640培養基。96 h后使用熒光顯微鏡進行綠色熒光蛋白觀察,有綠色熒光者為BTLA基因表達陽性。未感染慢病毒的DC為對照組(DCN)。
4.RT-PCR檢測BTLA在DC中的表達
根據NCBI數據庫中基因BTLA-F、BTLA-R、GAPDH-F、GAPDH-R的序列分別設計合成的引物序列,收集重組慢病毒LPP-Mm26645-Lv203-400轉染的DC,提取RNA,以DNaseI消化,電泳檢測RNA完整性,反轉錄為互補DNA(cDNA),進行RT-PCR檢測。
5.重組慢病毒LPP-Mm26645-Lv203-400轉染的DC的表型檢測
取重組慢病毒LPP-Mm26645-Lv203-400轉染成功的DC與PE-Cy5.5標記的CD11c、FITC標記的CD80、PE標記的CD86、APC標記的MHC-Ⅱ分子在4℃反應30 min,經PBS沖洗2次后用流式細胞儀檢測。
三、統計學處理
采用SPSS 20.0處理數據,計量資料組間比較采用t檢驗,P<0.05為差異有統計學意義。
結 果
一、慢病毒載體LPP-Mm26645-Lv203-400的構建、滴度與活性檢測
重組慢病毒顆粒由高純度ORF表達質粒(基因:BTLA、啟動子:CMV、C-flag標簽)、Endo-Fectin-LentiTM和Titer BoostTMreagents生成,并在Lenti-PacTMHIV packaging mix(Cat#:HPK-LvTR-20)作用下被包裝于293Ta細胞(Cat#:CLv-PK-01)中,轉染后48 h收集細胞,qPCR檢測本次病毒制備的滴度為3.25×109TU/ml。1 μl的LPPMm26645-Lv203-400轉染H1299細胞,3 d后在熒光顯微鏡下觀察,結果見圖1,重組慢病毒LPPMm26645-Lv203-400活性正常。
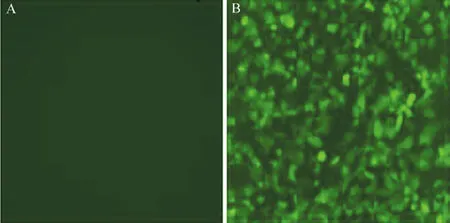
圖1 重組慢病毒LPP-Mm26645-Lv203-400轉染H1299細胞3 d后熒光顯微鏡觀察(×250)
二、體外培養DC的光鏡觀察與流式細胞儀檢測結果
CL57/BL小鼠的骨髓細胞在RPMI1640培養基體外培養24 h后,培養瓶底部可見半貼壁細胞,細胞呈圓形。培養第4日,細胞形態逐漸由圓形變為不規則形,個別細胞形成突起,可見細胞聚集現象,以粒細胞集落居多,集落由3~4個細胞組成,其后細胞直徑逐漸增大,集落中的細胞數可達8~9個。換液時傾去的培養基中的懸浮細胞主要為T淋巴細胞、B淋巴細胞、粒細胞、死細胞、細胞碎片及少量單核細胞和DC。培養第6日,細胞集落繼續增多,可見細胞逐漸呈懸浮狀態。收集培養5 d后(第6日)的DC進行流式細胞檢測,CD11c、CD86、CD80、MHC-Ⅱ是鑒別DC的主要標志物,其陽性表達率分別為73.59%、94.45%、47.84%、39.74%,為未成熟DC的表現。
三、重組慢病毒LPP-Mm26645-Lv203-400轉染DC結果
重組慢病毒LPP-Mm26645-Lv203-400以感染復數為15轉染培養5 d后的DC,96 h后在熒光倒置顯微鏡下可觀察到被感染的陽性細胞有綠色熒光蛋白表達(圖2)。在光鏡下觀察DC呈類圓形,部分細胞可見突起,細胞體積增加不明顯,大量DC呈懸浮狀態(圖3)。
四、RT-PCR檢測BTLA的表達
電泳檢測RNA完整,經過逆轉錄酶的作用得到cDNA,進行RT-PCR檢測,基因表達情況見圖4,與對照組相比,轉染BTLA基因的慢病毒載體的DC上BTLA基因表達量明顯提高。
五、轉染BTLA基因的慢病毒載體的DC的表型鑒定
收集重組慢病毒LPP-Mm26645-Lv203-400轉染96 h后的DC,流式細胞儀檢測CD11c--PE-Cy5.5、CD80--FITC、CD86--PE、MHC-Ⅱ-APC的陽性表達率分別為83.14%、89.06%、38.9%、41.28%,仍為未成熟DC的表現。
討 論
免疫抑制是當前治療自身免疫性疾病和移植后排斥反應的主要有效方法。特異性阻斷某些免疫反應而非全面地抑制免疫顯得尤為重要,近年來已有大量關于DC與T淋巴細胞作為配受體相互反應的研究[1]。抗原特異性T淋巴細胞的活化需要兩個信號,第一信號來自于APC主要組織相容性復合體(MHC)-抗原肽復合物與T淋巴細胞受體-CD3復合物,第二信號由APC表面的共刺激分子與T淋巴細胞上相應受體結合作用產生。BTLA是重要的負性抑制共刺激分子,包含免疫受體酪氨酸抑制基序(ITIM)。當BTLA與其配體HVEM結合,ITIM磷酸化,激活酪氨酸蛋白酶SHP-1和SHP-2,則會對T淋巴細胞增殖活化起抑制作用[5]。目前有研究表明,在一些自身免疫性疾病模型中,缺失BTLA-HVEM信號會引發嚴重的炎癥反應。BTLA基因敲除鼠誘導出的實驗性過敏性腦脊髓膜炎模型的發病率更高,病情更嚴重[6]。DC作為體內最強的APC,在免疫系統中起中心調節作用。Melton等[2]的研究表明DC對抗原的傳遞及其提呈的抗原肽的周轉均可影響實驗性自身免疫性疾病的結局。未成熟DC可以通過多種機制誘導對T淋巴細胞特異性抗原的耐受,但是單純的DC并不能完全抑制免疫反應。
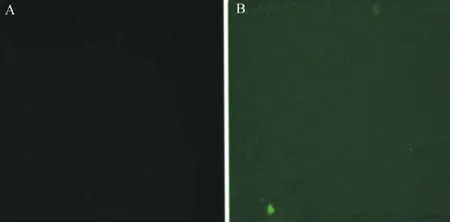
圖2 重組慢病毒LPP-Mm26645-Lv203-400轉染DC 96 h后熒光顯微鏡觀察(×250)
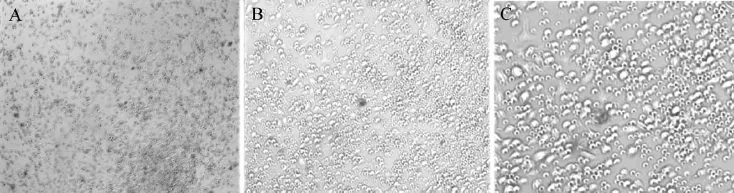
圖3 重組慢病毒LPP-Mm26645-Lv203-400轉染DC 96 h后倒置顯微鏡觀察
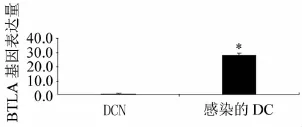
圖4 RT-PCR檢測感染的DC及DCN中BTLA基因表達與DCN組比較,*為P<0.05
慢病毒表達載體可以包裝、轉染、穩定整合所需要的遺傳信息,可以將外源基因有效整合到宿主染色體,從而達到持久性表達的目的,并且具有無插入致突變性、轉染率高、免疫反應小的特點。BTLA低表達于未成熟DC,故本研究根據Gene-Bank公布的小鼠BTLA的mRNA構建含有外源基因的重組載體,將測序正確、不含內毒素的重組質粒與復能基共轉染而得到高質量的病毒液LPPMm26645-Lv203-400液,并將其以感染復數為15轉染至原代培養至第6日的未成熟DC,經檢測證實未成熟DC有BTLA基因表達[7]。未成熟DC表面幾乎不表達BTLA,但本研究以慢病毒介導的BTLA成功轉染了DC,提高了未成熟DC上BTLA的表達量,而且感染的未成熟DC的表型并無變化。因此,構建攜帶BTLA基因的慢病毒載體可成功轉染BTLA至小鼠骨髓來源的未成熟DC,這為研究BTLA修飾的未成熟DC的免疫耐受作用甚至將其用于治療自身免疫性疾病或移植后排斥反應提供了實驗基礎。
致謝:感謝中山大學中山醫學院余新炳教授為本研究提供了技術性幫助。
[1]Del Rio ML,Lucas CL,Buhler L,Rayat G,Rodriguez-Barbosa JI.HVEM/LIGHT/BTLA/CD160cosignaling pathways as targets for immune regulation.J Leukoc Biol,2010,87(2):223-235.
[2]Melton AC,Bailey-Bucktrout SL,Travis MA,Fife BT,Bluestone JA,Sheppard D.Expression of αvβ8 integrin on dendritic cells regulates Th17 cell development and experimental autoimmuneencephalomyelitis in mice.J Clin Invest,2010,120(12):4436-4444.
[3]Said A,Weindl G.Regulation of dendritic cell function in inflammation.J Immunol Res,2015,2015:743169.
[4]Ilarregui JM,Rabinovich GA.Tolerogenic dendritic cells in the control of autoimmune neuroinflammation:an emerging role of protein-glycaninteractions.Neuroimmunomodulation,2010,17(3):157-160.
[5]Steinberg MW,Cheung TC,Ware CF.The signaling networks of the herpesvirus entry mediator(TNFRSF14)in immune regulation.Immunol Rev,2011,244(1):169-187.
[6]Shui JW,Steinberg MW,Kronenberg M.Regulation of inflammation,autoimmunity,and infection immunity by HVEM-BTLA signaling.J Leukoc Biol,2011,89(4):517-523.
[7]Murphy KM,Nelson CA,Sedy JR.Balancing co-stimulation and inhibition with BTLA and HVEM.Nat Rev Immunol,2006,6(9):671-681.
Study of immature dendritic cells transfected with lentiviral-mediated B and T lymphocyte attenuator
Wang Xuan,Yi Huan,Liu Jia,Li Min,Peng Fuhua.Department of Neurology,the Third Affiliated Hospital of Sun Yat-sen University,Guangzhou 510630,China
Objective To construct lentiviral vector expressing B and T lymphocyte attenuator(BTLA)gene and evaluate the feasibility of transfecting immature dendritic cells by this vector.Methods According to the mRNA sequence of mouse BTLA published by GeneBank,the ORF lentiviral plasmid was constructed and the recombinant lentiviral vector expressing BTLA was synthesized.Immature dendritic cells were subjected to primary isolation and culture from C57BL/6 mouse bone marrow.Immature dendritic cells were transfected with recombinant lentivial vector expressing BTLA at a multiplicity of infection of 15.The expression level of BTLA was assessed by immunofluorescence and RT-PCR.Results The lentiviral vector LPPMm26645-Lv203-400 was successfully constructed at a titer of 3.25×109TU/ml.At 96 h after immature dendritic cells transfected by the lentiviral vector LPP-Mm26645-Lv203-400,positive cells expressing green fluorescent protein were observed under inverted fluorescent microscope.The expression level of BTLA in the transfected immature dendritic cells was significantly higher compared with that in the non-transfected cells(P<0.05),indicating the successful transfection.Conclusion The lentiviral vector expressing BTLA gene can be successfully transfected into the mouse marrow-derived immature dendritic cells.
B and T lymphocyte attenuator;Lentivirus;Dendritic cell;Transfection
2015-07-15)
(本文編輯:洪悅民)

10.3969/j.issn.0253-9802.2015.12.004
國家自然科學基金(81271327);廣東省科技計劃項目(2011B03I8O0015)
510630廣州,中山大學附屬第三醫院神經內科
,彭福華,E-mail:pfh93@163.com
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
海峽科技與產業(2016年3期)2016-05-17 04:32:12
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
